
ABSTRACT
The diploid number of both Leiolepis belliana and L. boehmei was 2n = 2x = 36. In addition, the karyotype of the two species exhibited many conserved characteristics, including six pairs of macrochromosomes and 12 pairs of microchromosomes, the type and size of the macrochromosomes, and the position of the secondary constriction and nucleolar organizer region (NOR). However, while the position of the NOR region was identical in both species, in L. belliana it was found on both chromosomes of the first pair, and in L. boehmei it appeared on only one of the chromosome of the first pair, classified as 1a. No cytologically distinguishable sex chromosome was observed in either species. The karyotype formulae of L. belliana and L. boehmei were identical: 2n (36) = Lm4 + Mm6 + Sm2 + 24 microchromosomes.
Introduction
Leiolepis lizards, or butterfly lizards, are a member of the class Reptilia, order Squamata, suborder Lacertilia, family Agamidae, and subfamily Leiolepidinae. They are native to Thailand, Burma, Laos, Cambodia, Indonesia, and Vietnam. They are terrestrial lizards that prefer arid, open habitats. The genus Leiolepis Cuvier (1892), found only in southeast Asia, includes four bisexual diploids (2n) – L. belliana Hardwick & Gray (1827), L. guttata Cuuvier (1829), L. reevesii Gray (1831), and L. peguensis Peters (1971) – and two unisexual triploids (3n) – L. triploida Peters (1971) and L. guentherpetersi Darevsky and Kupriyanova (Citation1993). The genus also includes one putatively unisexual diploid, L. boehmei Darevsky and Kupriyanova (Citation1993) (Mertens Citation1961; Hall Citation1970; Peter Citation1971; Böhme Citation1982; Darevsky and Kupriyanova Citation1993; Schmitz et al. Citation2001; Das Citation2010; Grismer and Grismer Citation2010).
In Thailand, eight studies have reported on the chromosomal characteristics of Leiolepis species. L. reevesii rubritaeniata and L. b. belliana have diploid number of 36: 12 macro- (metacentric, m/submetacentric, sm) and 24 microchromosomes (Satrawaha and Ponkanid Citation1988; Puangwatana et al. Citation2002; Srikulnath et al. Citation2009, Citation2011; Phimphan et al. Citation2013). Satrawaha and Ponkanid (Citation1991) discovered parthenogenesis in a L. b. belliana population in Nakhon Si Thammarat and Songkhla Provinces by checking the chromosomes of embryos from unfertilized eggs: 2n = 36, 12 macro- (m/sm) and 24 microchromosomes.
Darevsky and Kupriyanova (Citation1993) were the first to describe L. boehmei, found in the southern Thai province of Songkhla, as a new Leiolepis species. The population they studied contained only females, leading to the parthenogenesis species hypothesis. In the same study, Darevsky and Kupriyanova (Citation1993) also reported female parthenogenesis in a L. guentherpetersi population in Vietnam. Its karyotype was a triploid number (3n) of 54 chromosomes (18 macrochromosomes and 36 microchromosomes), with secondary constriction or NOR on only one chromosome of the largest macrochromosome pair. Aranyavalai et al. (Citation2004) reported that L. boehmei from Nakhon Si Thammarat and Songkhla Provinces had a diploid number of 34, with 12 macrochromosomes and 22 microchromosomes. In addition, they found secondary constriction on the long arms of the largest metacentric chromosomes. Of 65 individuals examined, none were male. Srikulnath et al. (Citation2011) reported identical results in a chromosomal study of L. boehmei from Songkhla Province.
The molecular cytogenetics technique was used for chromosomal study in L. b. belliana and L. boehmei especially 18S and 28S rDNA genes. These two genes are located on the secondary constriction or nucleolar organizer region; they encode the pre rRNA and transform to 28S, 5.8S, and 18S rRNA. In L. b. belliana, the 18S-28S rDNA genes were located on the sub-telomeric region of the long arm of the largest macrochromosome in both chromosomes of the pair. In contrast, the 18S-28S rDNA genes of L. boehmei appeared in the same location of the largest macrochromosome, but the type was one normal chromosome and one changed chromosome of the pair (Srikulnath et al. Citation2011). In this study, we provide a karyomorphology by reviewing the current data and conducting additional chromosomal analysis using classical and Ag-NOR banding techniques.
Material and methods
Sample collection
Twenty Leiolepis belliana ()) specimens (10 male and 10 female) were collected at Ban Thiaraya (6°50ʹ55.9ʺN 101°20ʹ13.8ʺE), Takae Sub-district, Yaring District, Pattani Province, Thailand. Fifteen Leiolepis boehmei ()) specimens (all female) were collected from Sakom Beach (6°57'28.4"N 100°50'29.6"E), Sakom Sub-district, Thepha District, Songkhla Province, Thailand. They were transferred to the laboratory and kept under standard conditions for one day prior to the experiments.
Figure 1. General characteristics of Leiolepis belliana (A) and L. boehmei (B) from southern Thailand (scale bar indicates 5 cm).

Chromosome preparation, staining and analysis
Chromosomes were prepared by the direct method, from bone marrow and testis, followed by a colchicine, hypotonic, fixation, air-drying technique (Phimphan et al. Citation2013; Patawang et al. Citation2015a). The chromosomes were stained with 10% Giemsa for 30 min and NORs were identified through Ag-NOR staining (Howell and Black Citation1980; Rooney Citation2001). The length of short arm (Ls) and long arm (Ll) chromosomes were measured and calculated for the length of total arm chromosomes (LT, LT = Ls + Ll). Relative length (RL) and centromeric index (CI) were estimated. CI was also computed to classify the types of chromosomes (Turpin and Lejeune Citation1965; Chaiyasut Citation1989). All parameters were used in karyotyping and idiograming.
Results and discussion
Mitotic chromosome features from Giemsa staining
Both L. belliana and L. boehmei in this study had a diploid number of 36. The karyotype of these two species was comprised of four large metacentric, six medium metacentric, two small metacentric, and 24 microchromosomes. The fundamental number was 48, and no sex chromosome heteromorphisms were evident in either species (, ; and 3(A)). In L. belliana, the secondary constriction was found on the subtelomeric region of the long arm of the first macrochromosome pair; it appeared on both chromosomes of the first homologous pair (). In L. boehmei, the secondary constriction exhibited on the same region, but it appeared on only one chromosome of the first homologous pair, classifying as 1a ()). The diploid number and karyotypic features of L. belliana from this study accorded with Satrawaha and Ponkanid (Citation1988), Puangwatana et al. (Citation2002), Aranyavalai et al. (Citation2004), and Srikulnath et al. (Citation2011). In contrast, the diploid number (2n = 36) of L. boehmei from our study differed from Aranyavalai et al. (Citation2004) and Srikulnath et al. (Citation2011), who reported the same species with a diploid number of 34. However, we have carefully considered the diploid number of L. belliana and L. boehmei, as shown in . Each specimen of both L. belliana (20 specimens) and L. boehmei (15 specimens) shows the same chromosome number, 36 chromosomes per cell, in high percentages. Furthermore, only female L. boehmei were collected in this study; this agrees with the hypothesis of Darevsky and Kupriyanova (Citation1993) of parthenogenesis in L. boehmei. In addition, our presumption for the species of parthenogenetic L. b. belliana that was reported by Satrawaha and Ponkanid (Citation1991), it may be was the L. boehmei species.
Table 1. Mean length of the short arm chromosome (Ls), long arm chromosome (Ll), total arm chromosome (LT), centromeric index (CI), relative length (RL) and standard deviation (SD) of CI and RL from 20 metaphase cells of male and female Leiolepis belliana, 2n = 36.
Table 2. Mean length of the short arm chromosome (Ls), long arm chromosome (Ll), total arm chromosome (LT), centromeric index (CI), relative length (RL) and standard deviation (SD) of CI and RL from 20 metaphase cells of female Leiolepis boehmei, 2n = 36.
Table 3. A frequency of chromosome number per cell counts of Leiolepis belliana and L. boehmei.
Figure 2. Metaphase chromosome plates and karyotypes of male (A) and female (B) Leiolepis belliana, 2n = 36, by conventional staining technique. The secondary constrictions appear on the subtelomeric region of the long arm of the largest metacentric chromosome pair 1 (arrows). Scale bars indicate 5 µm.
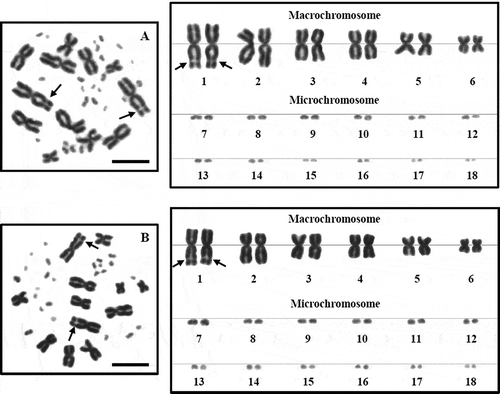
Figure 3. Metaphase chromosome plates and karyotypes of male (A) and female (B) Leiolepis belliana, 2n = 36, by Ag-NOR banding technique. Nucleolar organizer regions (NORs) appear on the subtelomeric region of the long arm of the largest metacentric chromosome pair 1 (arrows). Scale bars indicate 5 µm.
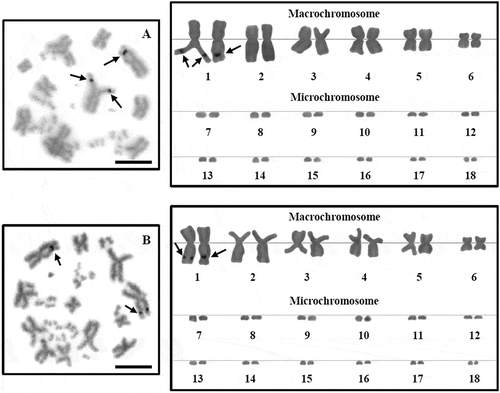
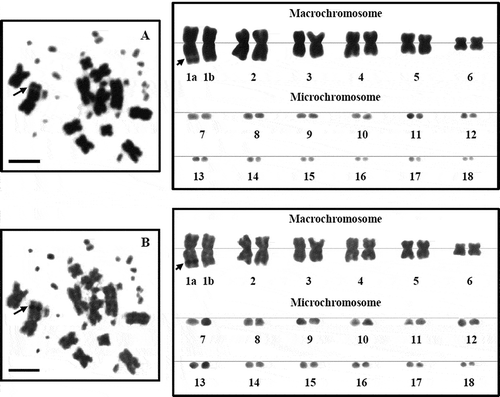
Figure 4. Metaphase chromosome plates and karyotypes of female L. boehmei, 2n = 36, by conventional straining (A) and Ag-NOR banding (B). The secondary constrictions or nucleolar organizer regions (NORs) appear on the subtelomeric region of the long arm of the largest metacentric chromosome pair 1a (arrow). Scale bars indicate 5 µm.
Nucleolar organizer region and meiotic cell characteristics
Our study, accomplished using the Ag-NOR staining technique, is the first cytogenetic report of L. belliana and L. boehmei. Nucleolar organizer regions (NORs) of these two species are clearly located in a subtelomeric position of the long arm of the 1st macrochromosome pair. However, the NOR of L. belliana appeared on both chromosomes of the first pair (()), while the NOR of L. boehmei appeared on only one chromosome of the first pair, classified as 1a (). Furthermore, the interphase and prophase cells of L. boehmei, which were stained by silver-NOR banding, showed one nucleolus complex per cell (). This feature supported the finding of the one signal that appeared on the metaphase chromosome of this species (). Our NOR findings from silver banding in both species were consistent with Srikulnath et al. (Citation2011) for both species; using a molecular cytogenetic method, they reported the appearance of 18S and 28S rDNA genes on both chromosomes of the 1st pair (1a) in L. belliana and on only one chromosome of the 1st pair in L. boehmei. However, we cannot detect the one changed chromosome by fusion of the 1st pair that was found by molecular technique following Srikulnath et al. (Citation2011). We forecast that the dissimilar of the signal resulting from responded different by the two techniques. The silver staining can positively detect argyrophilic associated protein, while the molecular technique can confidently detect DNA. We suggest the characteristics of NOR polymorphism by silver staining in L. boehmei; this feature is only one unique of parthenogenetic L. boehmei species.
Figure 5. Metaphase plate (A) and prophase (B) of female L. boehmei by silver-NOR banding, meiotic cell of male L. belliana during prophase by silver-NOR banding (C), and other meiotic cells of male L. belliana by conventional staining of mature sperm (D), leptotene (E), zygotene (F), pachytene (G), diakinesis (H), and metaphase II (I). Scale bars indicate 5 µm.
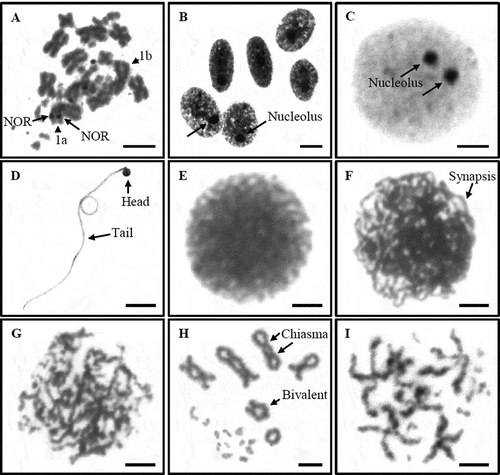
Our study is also the first to report on the meiosis features of male L. belliana; we found many chromosomal characterizations from testicular cells. Using silver-NOR banding, the prophase cells showed two nucleolus structures ()); this supported the finding that the NOR appeared on two metaphase chromosomes. Its sperm had a round head, about 1–1.5 µm in diameter, and a long tail, about 45–50 µm in length (). In prophase I, we found leptotene (), zygotene () and pachytene () by conventional staining technique. We also found 18 bivalent synapses between the homologous chromosome during diakinesis (). Moreover, we detected a haploid set (n) of 18 univalents of each chromosome pair in metaphase II (). These features, 18 bivalents in diakinesis and 18 univalents in metaphase II, confirmed the corrected diploid number, 2n = 36, of L. belliana.
Overview of chromosome study of Leiolepis and other agamid lizards
Our results agreed with much of the current knowledge of chromosomal and karyotypic features of Leiolepis, and we suggested a hypothesis for the cytotaxonomy and evolution in Leiolepis. All Leiolepis species have a highly conserved karyotype. The diploid and triploid Leiolepis species share many chromosomal features, including the number of homologous macrochromosomes (6) and homologous microchromosomes (12), the type and size of macrochromosomes, and the position of the secondary constriction or NOR. However, the secondary constriction or NOR differed little between the sexual and parthenogenetic species of Leiolepis. The secondary constriction or NOR appeared on both chromosomes of the largest pair in the species that reproduced sexually, while on only one chromosome of the largest pair in the diploid (from this study) and triploid (from Darevsky and Kupriyanova Citation1993) parthenogenetic species.
Leiolepis is one of many genera in the Agamidae family of lizards that have a conserved karyotype. For example, the species within the genus Calotes shared three classical unique features: a diploid number of 34, macro- and microchromosomes, and the position of the secondary constriction/NORs (Kritpetcharata et al. Citation1999; Patawang et al. Citation2015a). The genus Physignathus shares the chromosomal characteristics of Calotes, except it has a diploid number of 36 (Patawang et al. Citation2015b). From our result and other reports of the agamid lizard, these exhibited the high genetically conserved in chromosome level within each genus of this family. This supports the loss of few genetic materials in the evolutionary period of Agamidae.
Acknowledgments
This research was financially supported by the received scholarship grant from the Pattani Campus Research Fund, contract number: SAT600675S. We would like to thank the laboratory of Department of Science, Faculty of Science and Technology, Prince of Songkla University Pattani Campus for available help. The project was approved by the Institute of Animals for Scientific Purpose Development of National Research Council of Thailand (Resolution U1-02740-2559).
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Aranyavalai V, Thirakhupt K, Pariyanonth P, Chulalaksananukul W. 2004. Karyotype and unisexuality of Leiolepis boehmei Darevsky and Kupriyanova, 1993 (Sauria: Agamidae) from southern Thailand. Nat Hist J Chula Univ. 4:15–19.
- Böhme VW. 1982. Über Schmetterlingsagamen, Leiolepis b. belliana (Gray, 1827) der Malayischen Hallbinsel und ihre parthenogeneticschen Linien (Sauria: Uromastycidae). Zool Jahrb Syst. 109:157–169. [in Deutsch].
- Chaiyasut K. 1989. Cytogenetic and cytotaxonomy of genus Zephyranthes. Bangkok: Department of Botany, Faculty of Science, Chulalongkorn University. [in Thai].
- Darevsky IS, Kupriyanova LA. 1993. Two new all-female lizard species of the genus Leiolepis Cuvier, 1829 from Thailand and Vietnam (Squamata: Sauria: Uromastycinae). Herpetozoa. 6:3–20. [in Deutsch].
- Das I. 2010. A field guide to the reptiles of Thailand & south-east Asia. London: New Holland publishers Ltd.
- Grismer JL, Grismer LL. 2010. Who’s your mommy? Identifying maternal ancestors of asexual species of Leiolepis Cuvier, 1829 and the description of a new endemic species of asexual Leiolepis Cuvier, 1829 from Southern Vietnam. Zootaxa. 2433:47–61.
- Hall WP. 1970. Three probable cases of parthenogenesis in lizards (Agamidae, Chamaeleontidae, Gekkonidae). Experientia. 26:1271–1273.
- Howell WM, Black DA. 1980. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia. 36:1014–1015.
- Kritpetcharata O, Kritpetcharatb C, Luangpiromc A, Watcharanonc P. 1999. Karyotype of four Agamidae species from the Phu Phan National Park in Thailand. Sci Asia. 25:185–188.
- Mertens R. 1961. Die rassen der schmetterlingsagame, Leiolepis belliana. Senckenb Biol. 42:507–510. [in Deutsch].
- Patawang I, Tanomtong A, Chuaynkern Y, Chuaynkern C, Duengkae P. 2015a. Karyotype homology between Calotes versicolor and C. mystaceus (Squamata, Agamidae) from northeastern Thailand. Nucleus. 58(2):117–123.
- Patawang I, Tanomtong A, Jumrusthanasan S, Neeratanaphan L, Pinthong K, Jangsuwan N. 2015b. Karyological analysis of the Indo-Chinese Water Dragon, Physignathus cocincinus (Squamata, Agamidae) from Thailand. Cytologia. 80(1):15–23.
- Peter G. 1971. Die intragenerischen gruppen und die phylogenese der schmetterlingsagamen (Agamidae: Leiolepis). Zool Jahrb Syst. 98:11–130. [in Deutsch].
- Phimphan S, Tanomtong A, Patawang I, Kaewsri S, Jantarat S, Sanoamuang L. 2013. Cytogenetic study of Northeastern butterfly lizard, Leiolepis reevesii rubritaeniata (Squamata, Agamidae) in northeast Thailand. Cytologia. 78(2):133–140.
- Puangwatana V, Chulaluksananukul W, Pariyanonth P, Suwattana D. 2002. Karyological studied of the lizard Leiolepis belliana belliana at Samaesarn island and nearby islands in Chonburi Provinces. J Sci Res Chula Univ (Section T). 1(1):45–67.
- Rooney DE. 2001. Human cytogenetics constitution analysis. Oxford: Oxford University Press.
- Satrawaha R, Ponkanid S. 1988. Chromosome and karyotype of Leiolepis belliana rubritaeniata. KKU Sci J. 17:152–156. [in Thai].
- Satrawaha R, Ponkanid S. 1991. Parthenogenesis in Leiolepis belliana belliana (Gray) by chromosome number evaluation. J Wild Thailand. 1(1):53–59. [in Thai].
- Schmitz A, Vences M, Weitkus S, Ziegler T, Böhme VW. 2001. Recent maternal divergence of the parthenogenetic lizard Leiolepis guentherpetersi from L. guttata: molecular evidence (Reptilia: Squamata: Agamidae). Zool Abh. 51:355–360.
- Srikulnath K, Matsubara K, Uno Y, Thongpan A, Suputtitada S, Apisitwanich S, Matsuda Y, Nishida C. 2009. Karyological characterization of the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Squamata) by molecular cytogenetic approach. Cytogenet Genome Res. 125:213–223.
- Srikulnath K, Uno Y, Matsubara K, Thongpan A, Suputtitada S, Apisitwanich S, Nishida C, Matsuda Y. 2011. Chromosomal localization of the 18S-28S and 5S rRNA genes and (TTAGGG)n sequences of butterfly lizards (Leiolepis belliana belliana and Leiolepis boehmei, Agamidae, Squamata). Genet Mol Biol. 34(4):583–586.
- Turpin R, Lejeune J. 1965. Les chromosomes humains (caryotype normal et variations pathologiques). Paris: Gauthier-Villars. [in France].