Abstract
Palynologic assemblages from late Quaternary sediments from Disko Bugt, central West Greenland, contain a new acritarch species described herein as Palaeostomocystis subtilitheca sp. nov. The type of the genus Beringiella, Beringiella fritilla Bujak Citation1984, is transferred to the genus Palaeostomocystis Deflandre Citation1937, and the new combination Palaeostomocystis fritilla (Bujak Citation1984) is proposed. Palaeostomocystis subtilitheca sp. nov. and P. fritilla (Bujak Citation1984) comb. nov. are characteristic of polar to subpolar environments and may be potential environmental indicators.
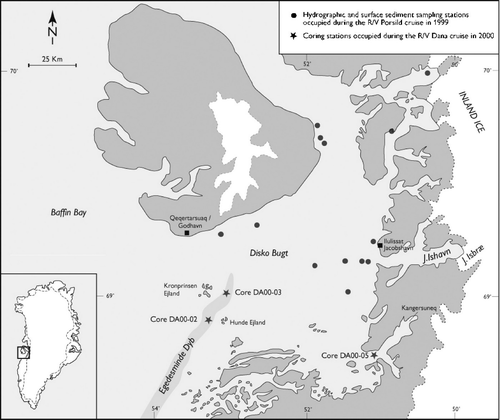
An extensive palynological study on Quaternary sediments from Disko Bugt, central West Greenland was carried out as part of an ongoing palaeoceanographic project at the Geological Survey of Denmark and Greenland (GEUS). Quaternary sediments are scattered in the central and eastern part of Disko Bugt, which is mainly dominated by outcrops of Palaeogene rocks and glacial deposits (CitationLloyd et al. Citation1999, Kuijpers et al. 2000). The water depth in the bay ranges between 200 and 400 m; however, a 990 m deep submarine valley (Egedesminde Dyb) occurs south of Disko in the area between Kronprinsen and Hunde Ejland and the shelf edge further to the southwest.
The Disko region has a polar maritime climate with high precipitation rates. In Qeqertarsuaq, the mean annual temperature is −4°C with 450 mm/yr precipitation (Gilbert et al. Citation1998). During the summer, the meltwater production from the Ilulissat glacier, Inland Ice margin, sea ice and iceberg discharges cause freshening of the sea surface establishing a water column stratification in the bay. High vertical and horizontal gradients in salinity and temperature characterise the bay during the summer, and a 20–30 m surface layer with temperatures between 3–7°C and salinity ≤33.25‰ occurs (E. Buch: pers. comm. 2002). In the autumn, lower gradients are observed in the horizontal and vertical distribution of salinity and temperature mainly due to a decreased meltwater production from the Ilulissat glacier. A 50–75 m surface layer with temperatures ≤1.5°C and salinity ranging 32.75–33.25‰ occurs. During the winter, seasonal sea ice covers the bay for 3–5 months per year (Nielsen et al. Citation2001).
The oceanographic conditions in Disko Bugt differs from what can be observed offshore southwestern Greenland because of the presence of the Ilulissat glacier as well as the lack of warm Atlantic water which does not enter the bay (E. Buch: pers. comm. 2002). The hydrography in the bay is characterised by the presence of the West Greenland Current and cold surface waters advected from the Baffin Bay (Buch Citation2000). The circulation is cyclonic: inflow occurs through the southern part of a line between Qeqertarsuaq and Egedesminde Dyb, whereas outflow occurs through the northern part (Gilbert et al. Citation1998). Archaeological and geomorphologic information combined in a discussion about relative sea level changes in the southern part of the bay during the Holocene are provided by Rash & Jensen (Citation1997).
In this paper, the new acritarch Palaeostomocystis subtilitheca is described and the new combination Palaeostomocystis fritilla (Bujak Citation1984) is proposed. These two taxa occur in sediments deposited during the last 2700 calendar years Before Present (cal. yr BP) (before present=1950), in a transect from fjord to shelf valley setting. The chronology of the material studied is based on accelerated mass spectrometry {\rm ^{14}} C dating on shells and marine plant remains. The ages are reported in cal. yr BP and a standard marine reservoir correction of 400 years is used in accordance with the international convention (Stuiver & Polach Citation1977).
MATERIAL AND METHODS
This paper was based on the study of palynofacies assemblages from 58 samples from three boreholes drilled during the 2000 cruise of R/V Dana in Disko Bugt, central West Greenland (). The geographic position of the borehole DA00-05 is in Kangersuneq Fjord, on the south east margin of the bay, at 68°42.92′N and 51°06.42′W, at a depth of 335 m and 1600 m from the shore. The boreholes DA00-02 and -03 from Egedesminde Dyb are located at 68°51.88′N, 53°19.72′W and 69°00.00′N, 53°08.00′W, and at a depth of 840 and 852 m respectively, both occur approximately 24 km from the shore.
Location of the coring stations in Disko Bugt, central West Greenland (modified from Kuijpers et al. Citation2000).
Palaeostomocystis subtilitheca sp. nov. and P. fritilla (Bujak Citation1984) comb. nov. were recorded in the upper 230 cm of core DA00-05 (c. 2700 cal. yr BP), 243 cm of core DA00-02 (c. 1300 cal. yr BP) and 267 cm of core DA00-03 (c. 1300 cal. yr BP). In these intervals, the sediments consist of olive grey to brown, muddy silt, locally mottled and laminated and characterised by specks, shell fragments and plant debris. The water content in the sediments from the upper part of the cores was high.
Standard palynological technique, involving the use of warm HCl and HF (70%), was used (Wood et al. Citation1996). The residue was sieved on 10 μm filter cloth and was not oxidised. The macerate was mounted in glycerine jelly on microscope slides. The type specimens of Palaeostomocystis subtilitheca sp. nov. and all specimens figured herein are from single mounts; they are identified with MGUH numbers and housed in the type collection of the Geological Museum of the University of Copenhagen, Copenhagen, Denmark. Core DA00-02, -03 and -05 are stored at GEUS, Copenhagen, Denmark.
SYSTEMATIC PALYNOLOGY
Acritarch genera and species mentioned below are referenced in Fensome et al. (Citation1990); informal taxa are referenced herein. The terminology follows Williams et al. (Citation2000). The format of the taxonomic descriptions follows Punt (Citation1994).
Group incertae sedis ACRITARCHA Evitt Citation1963
Genus Palaeostomocystis Deflandre Citation1937
Palaeostomocystis Deflandre Citation1937
Beringiella Bujak Citation1984
Type species. – Palaeostomocystis reticulata Deflandre Citation1937
Translation of the original diagnosis. – “The small vesicles of organic matter included in this genus are always characterised by the presence of a pore or more or less large opening. This opening can be with or without a variably developed collar. The general shape of the vesicle is variable in the species described until now: spheroidal, ellipsoidal and ovoidal; other shapes will certainly be found. The wall surface can be psilate or ornamented. The dimensions are always within the order of c. 10−40 μm” (Deflandre Citation1937: p. 52).
Remarks. – In 1937, Deflandre established the genus Palaeostomocystis to include small, single-layered, organic-walled vesicles characterised by the presence of a large pylome. This genus lacks morphological features of unequivocal dinoflagellate affinity (Fensome et al. Citation1990). Based on the original diagnosis, the shape of the vesicle is highly variable, the wall surface is psilate to ornamented and a collar may occur around the pylome (Deflandre Citation1937: p. 52). Although Stover & Evitt (Citation1978: p. 69) considered the description given by Deflandre (Citation1966: p. 6) as an emendation of Palaeostomocystis, this genus has never been formally emended in the literature.
Beringiella fritilla (Bujak Citation1984), the type species of the genus Beringiella Bujak (Citation1984) is herein transferred to Palaeostomocystis, making Beringiella a taxonomic junior synonym of Palaeostomocystis.
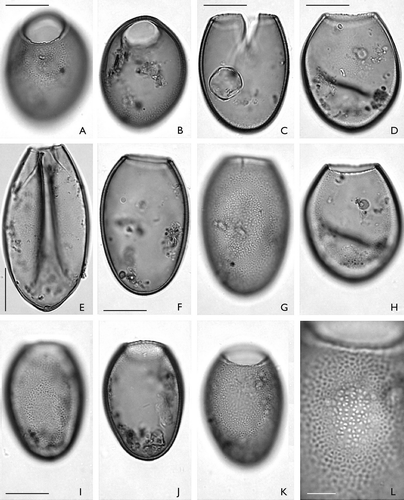
Palaeostomocystis fritilla (Bujak Citation1984) comb. nov. ( A−L)
1984 Beringiella fritilla n. sp. – Bujak: p. 195–196, plate 4, Figs 12–14.
A–L. Palaeostomocystis fritilla (Bujak Citation1984) comb. nov. from core DA00-03, Egedesminde Dyb, central West Greenland. The same scale is used for all specimens except (L). A. MGUH 26853. Semilateral view in high focus. Note the undulate pylome margin. B. Same specimen as (A). Semilateral view in sectional focus. C. MGUH 26855. Lateral view in sectional focus. Damaged specimen with operculum in the interior of the vesicle. D. MGUH 26856. Lateral view in high focus. E. MGUH 26858. Lateral view in sectional focus. F. MGUH 26854. Lateral view in low focus. Note the thick vesicle wall. G. Same specimen as (F). Lateral view in high focus. Note the faveolate wall structure. H. Same specimen as (D). Lateral view in low focus. I. MGUH 26857. Lateral view in low focus. J. Same specimen as (I). Lateral view in sectional focus. K. Same specimens as (I). Lateral view in high focus. L. Same specimen as (I). Faveolate wall structure in high magnification. The faveoli internal part is in focus in the centre of the picture; the reticulate muri network is shown on the periphery of the picture. Scale bars – 15 μm (A – K); 5 μm (L).
Basionym. – Beringiella fritilla Bujak Citation1984: Micropaleontology 30, pp. 195–196, plate 4, Figs 12–14.
Holotype. – Bujak Citation1984: plate 4, Figs 12,13; sample SGM 535, England Finder coordinates L39.
Repository. – Holotype deposited in the palynology type collection of the Geological Survey of Canada.
Type locality and type stratum. – DSDP Leg 19, Site 191B, core 1, section 5, interval 56–58 cm, from the Aleutian Basin, northern North Pacific (Bujak Citation1984: pp. 180 & 195).
Original diagnosis. – “Ovoidal to ellipsoidal organic-walled microfossils with thick faveolate wall. One of the poles is lost by the development of a large pylome” (Bujak Citation1984: p. 195).
Dimensions. – Range (N=10): overall vesicle length 55 (58.5) 66 μm, overall vesicle width 39 (43.4) 50 μm, pylome diameter 19 (24.2) 37 μm.
Stratigraphic range. – Late Pleistocene to late Quaternary.
Remarks. – According to Bujak (Citation1984: p. 195), B. fritilla differs from species of Palaeostomocystis by lacking “narrow, relatively high muri which form a reticulate network”. However, this feature is not mentioned in the original diagnosis of Palaeostomocystis which can have smooth or variably ornamented wall; thus, the diagnosis of B. fritilla is compatible with that of Palaeostomocystis.
Beringiella fritilla, the type species of the genus Beringiella, is transferred to Palaeostomocystis, making genus Beringiella a taxonomic junior synonym of Palaeostomocystis. Other species formerly assigned to Beringiella after 1998 or not mentioned by Williams et al. (Citation1998: Appendix A) must be reassigned.
Palaeostomocystis fritilla differs from the other species of this genus by having a faveolate wall ( L).
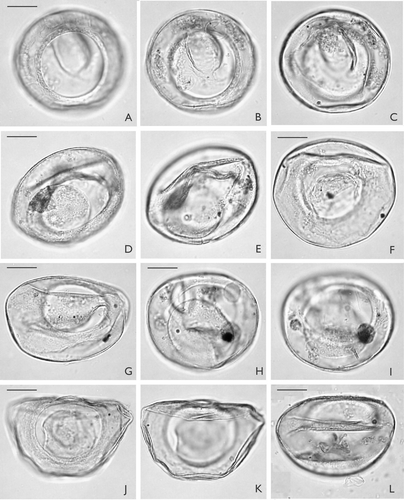
Palaeostomocystis subtilitheca sp. nov. ( A−L, 4 A−I)
Holotype. – MGUH 26841 ( A−C)
A–L. Palaeostomocystis subtilitheca sp. nov. from core DA00-03, Egedesminde Dyb, central West Greenland. Note that the operculum occurs in the interior of each specimen except G. The same scale is used for all specimens. A. Holotype. MGUH 26841. Apical view in high focus. Note the pylome rim. B. Same specimen as (A). Apical view in sectional focus. C. Same specimen as (A). Apical view in low focus. D. MGUH 26842. Semilateral view, sectional focus. E. Same specimen as (D). Semilateral view in low focus. F. MGUH 26843. Apical view in low focus. G. MGUH 26845. Antapical view in high focus. H. MGUH 26846. Antapical view in high focus. Note small pseudopylome at 2.00 o'clock. I. Same specimens as (H). Antapical view in low focus. J. MGUH 26847. Apical view in high focus. Specimen partly damaged by lateral compression. K. Same specimen as (J). Apical view in low focus. L. MGUH 26851. Apical view in low focus. Note dome-shaped operculum in the bottom of the vesicle. Scale bar – 15 μm.
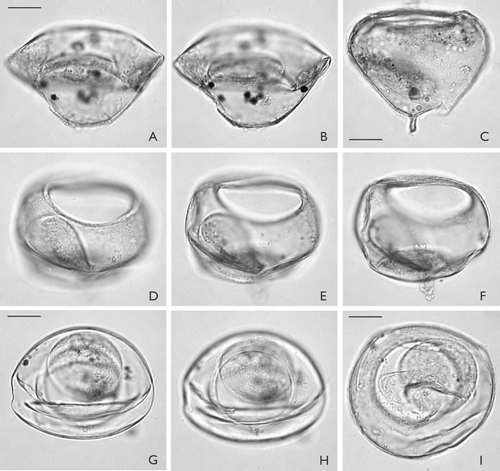
Paratypes. – MGUH 26848 ( D−F); MGUH 26850 ( I)
A–I. Palaeostomocystis subtilitheca sp. nov. from core DA00-03, Egedesminde Dyb, central West Greenland. The same scale is used for all specimens. A. MGUH 26844. Lateral view in high focus. Specimen partly damaged by vertical compression. B. Same specimen as (A). Lateral view in low focus. Note flat operculum in the interior of the vesicle. C. MGUH 26849. Lateral view in sectional focus. Note single, solid and blunt horn at the pole diametrically opposite the pylome. D. Paratype 1. MGUH 26848. Lateral view in high focus. Note the well-developed pylome rim and the dome-shaped operculum within the vesicle. E. Same specimen as (D). Lateral view in sectional focus. F. Same specimen as (D). Lateral view in low focus. G. MGUH 26852. Antapical view in sectional focus. H. Same specimen as (G). Antapical view in low focus. I. Paratype 2. MGUH 26850. Apical view in sectional focus. Scale bar – 15 μm.
Repository. – The type collection of the Geological Museum of the University of Copenhagen, Copenhagen, Denmark.
Type locality. – Egedesminde Dyb, Disko Bugt, central West Greenland.
Type stratum. – Holocene muddy silt from the upper 267 cm of core DA00-03.
Etymology. – Latin: subtilis, -e=thin; theca, -ae=wall. With reference to the thin wall that characterises this species.
Diagnosis. – Vesicula cum muro unico et subtili, corpore ovato vel obtusi-conico. Superficies muri levis vel perpolite granosa. Pyloma grandis, rotundus, circumsutus cum operculo grandi quod ex consuetudine liberum est intus vesicula. Nonnunquam visibilis parvus falsus pyloma, rotundus vel ovatus et locatus prope ab apice. Ovoidal to obtusi-coniform, unilayered, thin-walled vesicle with circular to oval equatorial outline. Wall surface psilate to microgranulate. Macropylome circular, rimmed and operculate with detached macro operculum usually in the interior of the vesicle. Small, circular to oval pseudopylome occasionally present at or near the antapical pole.
Description. – The vesicle consists of a single-layered, thin (≤0.5 μm) wall, which is smooth or finely granulate on the surface. Occasionally, a solid, blunt horn occurs in the antapical region ( C). The excystement structure is very large (>40% of vesicle diameter) ( A, 4 D) and generally, the area around the macropylome is characterised by few, concentric rings. A flat to dome-shaped, circular macro operculum detached from the macropylome rim, often occurs in the interior of the vesicle. The operculum surface is psilate to microgranulate. In some specimens, a small, antapical pseudopylome, circular to oval in shape occurs at the antapex, diametrically opposing the pylome ( H).
Dimensions. – Holotype: overall vesicle length 55 μm, overall vesicle width 57.5 μm, pylome diameter 35 μm. Range (N=16): overall vesicle length 45 (51.6) 55 μm, overall vesicle width 52.8 (63.3) 78 μm, pylome diameter 29.7 (35.5) 44 μm, pseudopylome length (N=7) 9 (12.5) 20 μm, pseudopylome width (N=7) 13 (12.5) 15 μm.
Occurrence. – The new taxon is found in Holocene sediments from Egedesminde Dyb and Kangersuneq Fjord, Disko Bugt, central West Greenland; and from Skálafjord, Eysturoy, Faroe Islands. The highest abundance is recorded in sediments deposited under conditions of stratified waters during the summer months of the year and characterised by 4°C mean summer sea surface temperature and salinity ≤33.25‰.
Discussion. – Palaeostomocystis subtilitheca differs from P.? pachytheca Cookson & Eisenack 1971, and P. ovata (Wilson 1967) Eisenack et al. 1973 by having a very thin, transparent wall. It differs further from P.? pachytheca by being larger and from P. ovata by having a psilate to microgranulate wall surface and detached operculum. The new species differs from Palaeostomocystis punctulosa Deflandre 1941, by being twice the size, thin-walled and by having a macropylome. Palaeostomocystis subtilitheca differs from P. fritilla (Bujak Citation1984) comb. nov. by lacking the readily discernible faveolate wall structure that characterises the latter taxon ( L). Palaeostomocystis subtilitheca differs from ?Palaeostomocystis sp. of Baltes (Citation1971) figured in Mudie (Citation1987: pl. 5, fig. 21) in its general outline and by having a circular pylome.
Palaeostomocystis subtilitheca resembles species of the genus Leiosphaeridia Eisenack (1958) Downie & Sarjeant 1963, in its equatorial outline and by being thin-walled. However, the new species differs from Leiosphaeridia by having ovoidal to obtusi-coniform outline in the lateral view and a macropylome.
Palaeostomocystis subtilitheca resembles also species of Fromea Cookson & Eisenack (1958) Yun Hyesu 1981, in overall size and shape but it is readily distinguishable by lacking indication of paracingulum and by having occasionally wall surface ornamentation.
The modern cyst of unknown biological affinity described as Fungella arctica by Cleve (Citation1899) and renamed Pacillina arctica by Cleve (Citation1903) closely resembles P. subtilitheca in overall shape and having a circular macropylome marked by concentric rings. However, the new species differs from Pacillina arctica by being thin-walled and lacking the antapically expanded outer layers that characterise the latter taxon (Reid & John Citation1981).
Palaeostomocystis subtilitheca resembles some of the morphotypes described by van Waveren (Citation1994) as tintinnid palynomorphs from recent marine sediments, particularly, the ARABNO-1 and -2 morphotypes (van Waveren Citation1994: Figs 11, 13 & 15). However, the size of the new taxon and the presence of an operculum in many of the study specimens exclude the possibility of describing it as tintinnid lorica. Palaeostomocystis subtilitheca may though represent a tintinnid palynomorph cyst and have biological affinities with heterotrophic cilates, which are abundant in the waters of Disko Bugt (Munk et al. Citation2003).
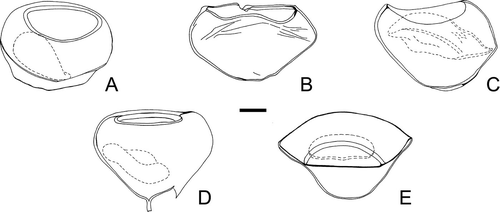
Despite the high morphologic variability observed in the lateral outline () and wall surface of P. subtilitheca, its circular to oval equatorial outline, its single-layered, thin wall and the presence of a circular macropylome are invariable features that facilitate its identification.
A–E. Variation in the overall shape of Palaeostomocystis subtilitheca sp. nov. seen in lateral view. The operculum is indicated by a dashed line. The specimens are in the same scale. A. Paratype 1, illustrated on D–F. D. Illustrated on C. E. Illustrated on A−B. Scale bar – 15 μm.
ACKNOWLEDGEMENTS
The author acknowledges funding from the Carlsberg Foundation, Copenhagen, Denmark and publishes with the permission of the Geological Survey of Denmark and Greenland. Thanks to Johnny E. Hansen (GEUS, Copenhagen, Denmark) for help in processing the samples. Antoon Kuijpers (GEUS) is thanked for making the core material available for this study as well as providing the AMS radiocarbon dates, which were performed under the supervision of Jan Heinemeier (University of Aarhus, Denmark). Clara Fini Sandri (Liceo San Carlo, Modena, Italy) kindly checked the Latin diagnosis. Barrie Dale (University of Oslo, Norway) and Jens Matthiessen (Alfred Wegener Institute, Bremerhaven, Germany) are thanked for their helpful comments on the initial manuscript.
Related Research Data
REFERENCES
- Baltes N. 1971 Pliocene Dinoflagellata and Acritarcha in Romania. – In: 2 {\rm ^{nd}} Planktonic Conf. Rome 1970. Proc. Vol. 1. (ed. A. Farinacci), pp. 1−11. – Org. Comm., Rome.
- Buch , E. 2000 . A monograph on the physical oceanography of Greenland waters. . – Dan. Meteorol. Inst. Sci. Rep. , 00–12 : 1 – 405 .
- Bujak , J. P. 1984 . Cenozoic dinoflagellate cysts and acritarchs from the Bering Sea and northern North Pacific, DSDP Leg 19. . – Micropaleontology , 30 : 180 – 212 .
- Cleve , P. T. 1899 . Plankton collected by the Swedish expedition to Spitzbergen in 1898. . – K. Sven. Vetenskapsakad. Handl. , 32 (3) : 1 – 48 .
- Cleve P. T. 1903 August 1902–May 1903: surface plankton collected in the North Sea. – In: Variations de la température de l'eau de surface dans certains carrés choisis de l'Atlantique pendant les années 1900–1913, Part D. 1902–1903 (ed. ICES/CIEM), pp. 296–297. – ICES/CIEM (Cons. Permanent Int. Explor. Mer) Bull. Hydrogr. 1903, Copenhagen.
- Deflandre , G. 1937 . Microfossiles des silex crétacés. Deuxième partie. Flagellés incertae sedis. Hystrichosphaeridés. Sarcodinés. Organismes divers. . – Ann. Paléontol. , 26 : 51 – 103 .
- Deflandre , G. 1966 . Addendum à mon mémoire: Microfossiles des silex crétacés. . – Centre Doc. Arch. Orig. No. 419, Cah. Micropaléontol. , 1 (2) : 1 – 9 .
- Evitt , W. R. 1963 . A discussion and proposals concerning fossil dinoflagellates, hystrichospheres and acritarchs. I. . – Proc. Natl Acad. Sci. USA , 49 : 158 – 164 .
- Fensome R. Williams G. L. Sedely Barss M. Freeman J. M. Hill J. M. 1990 Acritarchs and fossil prasinophytes: an index to genera, species and infraspecific taxa. – Contrib. Ser. AASP Found. No. 25. AASP, Houston TX.
- Gilbert , R. , Nielsen , N. , Desloges , J. and Rasch , M. 1998 . Contrasting glaciomarine environments of two Arctic fjords on Disko, West Greenland. . – Mar. Geol. , 147 : 63 – 83 .
- Kuijpers , A. , Lloyd , J. M. , Jensen , J. B. , Endler , R. , Moros , M. , Park , L. A. , Schulz , B. , Jensen , K. G. and Laier , T. 2000 . Late Quaternary circulation changes and sedimentation in Disko Bugt and adjacent fjords, central West Greenland. . – Geol. Surv. Greenland. Bull. , 189 : 41 – 47 .
- Lloyd J. M. Kuijpers A. Hansen E. 1999 Marine geological investigations in Disko Bugt as part of the UK NERC ARCICE project, Late Quaternary Ice Sheet Dynamics in West Greenland. – Cruise Report R/V Porsild 1999. Durham University, Durham UK.
- Mudie P. J. 1987 Palynology and dinoflagellate biostratigraphy of Deep Sea Drilling Project Leg 94, sites 607 and 611, North Atlantic Ocean. – In: Initial Reports of the Deep Sea Drilling Project. Vol. 94. (ed. W. F. Ruddiman, R. B. Kidd & E. Thomas et coll.), pp. 785−812. – U.S. Gov. Print. Office, Washington DC.
- Munk , P. , Hansen , B. W. , Nielsen , T. G. and Thomsen , H. A. 2003 . Changes in plankton and fish larvae communities across hydrographic fronts off West Greenland. . – J. Plankton Res. , 25 : 815 – 830 .
- Nielsen , N. , Humlum , O. and Hansen , B. U. 2001 . Meteorological observations in 2000 at the Arctic Station, Queqertarsuaq (69°15′N), Central West Greenland. . – Dan. J. Geogr. , 101 : 155 – 157 .
- Punt , W. 1994 . Format of descriptions of new taxa of fossil plants (genera, species). . – Rev. Palaeobot. Palynol. , 80 : 7 – 8 .
- Rash , M. and Jensen , J. F. 1997 . Ancient eskimo dwelling sites and Holocene relative sea-level changes in southern Disko Bugt, central West Greenland. . – Polar Res. , 16 : 101 – 115 .
- Reid , P. C. and John , A. W. G. 1981 . A possible relationship between Chitinozoa and Tintinnids. . – Rev. Palaeobot. Palynol. , 34 : 251 – 262 .
- Stover L. E. Evitt W. R. 1978 Analysis of pre-Pleistocene organic-walled dinoflagellates. – Stanford Univ. Publ., Geol. Sci. 15, UK.
- Stuiver , M. and Polach , H. A. 1977 . Discussion: Reporting of {\rm ^{14}} C Data. . – Radiocarbon , 19 : 355
- Van Waveren , I. M. 1994 . Tintinnomorphs from deep-sea sediments of the Banda Sea (Indonesia). . – Scr. Geol. , 105 : 27 – 51 .
- Williams G. L. Fensome R. A. Miller M. A. Sarjeant W. A. S. 2000 A glossary of the terminology applied to dinoflagellates, acritarchs and prasinophytes, with emphasis on fossils: third edition. – Contrib. Ser. AASP Found. No. 37. AASP, Houston TX.
- Williams G. L. Lentin J. K. Fensome R. A. 1998 The Lentin and Williams index of fossil dinoflagellates, 1998 edition. – Contrib. Ser. AASP Found. No. 34. AASP, Houston TX.
- Wood G. D. Gabriel A. M. Lawson J. C. 1996 Palynological technique – processing and microscopy. – In: Palynology: Principles and applications. Vol. 1. (ed. J. Jansonius & D. C. McGregor), pp. 29−50. – AASP Foundation, Dallas TX.