Abstract
Pollen morphology of ten Enkianthus species was examined using light (LM) and scanning electron microscopy (SEM) and previous infrageneric classification of Enkianthus was discussed in the light of the palynological characters obtained in this study. The pollen grains are monads, oblate spheroidal to prolate in shape and 3‐ to 5‐colpor(oid)ate. Exine sculpture varies from granulate to coarsely rugulate‐psilate with faint to distinct minute granules in SEM. Infraspecific geographical differences in palynological characters were found in E. deflexus. The species of Enkianthus are clearly differentiated into two pollen morphological groups that correspond to the sections Enkiantella + Meisteria and Andromedina + Enkianthus, respectively. The members of the sections Enkiantella and Meisteria have pollen grains typically with three apertures, prolate spheroidal to prolate, ratio of colpus length to polar axis (L/P) 0.69–0.84, relatively thinner exine and exine sculpturing tendency to granulate, character states that are probably plesiomorphic in this genus. The members of the sections Andromedina and Enkianthus have more derived pollen grains with four to five apertures, commonly oblate spheroidal, L/P 0.56–0.63, relatively thicker exine and exine sculpturing tendency to coarsely rugulate‐psilate. It seems to be difficult to differentiate between sections of the same group on the basis of palynological characters only. Adding the palynological characters to the infrageneric framework of Anderberg indicates an evolutionary trend in exine sculpture within Enkianthus from finely verrucate‐rugulate through coarsely rugulate to coarsely rugulate‐psilate. A dichotomous key for Enkianthus was prepared based on the palynological characters.
The genus Enkianthus Lour. belongs to the subfamily Enkianthoideae Kron, Judd and Anderb. (Ericaceae) and is composed of about 12 species occurring in E. Himalayas, China, Japan, Taiwan, and Indochina (Fang & Stevens, Citation2005). Monophyly for Enkianthus is strongly supported by morphological and molecular data (Ueno, Citation1950; Hsu, Citation1982; Anderberg, Citation1994; Kron et al., Citation2002). However, the number of species in the genus is still controversial. Palibin Citation(1899) recognized nine species in his revision of the genus Enkianthus. In recent studies, the number of species varies from 13 (Hsu, Citation1982) to 17 (Anderberg, Citation1994), 15 (Kron et al., Citation2002), or 12 (Fang & Stevens, Citation2005). Plants are shrubs or small trees and different species of this genus are used as ornamental plants in the garden or as a mixed border plant.
Enkianthus differs from other Ericaceae in having anthers with fibrous endothecium, pollen grains in monads without viscin threads, and seeds with vascular bundles in the raphe (Anderberg, Citation1994). Most recent morphological and molecular cladistic analyses indicate that Enkianthus is sister to the Ericaceae s.l. including Empetraceae and Epacridaceae (Anderberg, Citation1993, Citation1994; Judd & Kron, Citation1993; Kron & Chase, Citation1993; Kron, Citation1996, Citation1997; Kron et al., Citation2002) and is treated as a separate subfamily, Enkianthoideae, consisting of a single genus Enkianthus in the Ericaceae (Kron et al., Citation2002). Within the genus, species differ in leaf texture, inflorescence structure, corolla shape and seed morphology (Kron et al., Citation2002), and number of pollen apertures (Ueno, Citation1950; Ikuse, Citation1956, Citation2001; Anderberg, Citation1994). Several reports on infrageneric classification of Enkianthus are available (Palibin, Citation1899 cited from Anderberg, Citation1994; Hara, Citation1948; Ueno, Citation1950; Hsu, Citation1982; Anderberg, Citation1994). In the most recent classification, Anderberg Citation(1994) considered Enkianthus to be monophyletic and recognized four sections: i.e., Enkiantella, Meisteria, Andromedina and Enkianthus, based on cladistic analysis of morphological, anatomical, embryological and cytological data.
Mainly light microscope (LM) observations of pollen grains of Enkianthus have been carried out previously (Ueno, Citation1950; Ikuse, Citation1956, Citation2001; Nakamura, Citation1980; Wang et al., Citation1995). Ueno divided the genus into three subgenera, e.g. Euenkianthus, Andromedina and Tritomodon, based upon the characteristics of pollen grains by LM and crystals in the leaf. According to Ueno Citation(1950), pollen grains of Enkianthus are unicellular (monad) with three to four apertures, entirely different from other genera of the Ericaceae. Scanning electron microscopic (SEM) observations of pollen grains in two species, E. campanulatus and E. perulatus, were published previously (Kurosawa, Citation1991: Plate 83; Zhang & Anderberg, Citation2002: Fig. 8). In order to provide new information and to discuss the new pollen morphological data in light of recent infrageneric classification, the present study presents a detailed examination of pollen morphology of the genus using both LM and SEM.
Material and methods
Pollen of 13 taxa of ten species and 29 specimens of Enkianthus was examined by means of LM and SEM. Pollen used in this investigation was taken from dried specimens from the herbaria E, SAPS, SAPT and TI. Abbreviation of the herbarium names except for SAPT (the Botanic Garden, Hokkaido University, Sapporo) are according to the Index Herbariorum (Holmgren et al., Citation1990). Infrageneric classification follows Anderberg Citation(1994) and taxonomic treatment for Japanese species follows Yamazaki Citation(1993).
Pollen was acetolysed following the technique of Erdtman Citation(1960) modified by Takahashi Citation(1987). For LM, pollen was mounted in silicone oil (viscosity 3000 cs), and examined and measured with a Nikon Eclipse E200 microscope. The dimensions “P”, “E” and “L”, corresponding to the polar length, equatorial length and colpus length of pollen were measured, and the P/E and L/P ratio calculated. Measurements given in Table are based on at least ten grains from each specimen except in E. cernuus and one specimen of E. campanulatus (Sukawa s.n.). Pollen size and shape classes follow Erdtman Citation(1986). Pollen slides of the whole collection are deposited at the Hokkaido University Museum, Sapporo, Japan.
For SEM, acetolysed pollen samples were dehydrated in an ethanol series and mounted and air dried on aluminum stubs from 70% ethanol, and sputter coated with Platinum‐Palladium by a Hitachi E102 ion sputter. Subsequently these were examined and photographed with a Jeol JSM‐5310 LV scanning electron microscope operated at 15 KV. Descriptive terminology follows Punt et al. Citation(1994).
Palynological features (Table ) were plotted on the phylogenetic tree of Anderberg (Citation1994: Fig. 9). Palynological features of outgroup taxon, Clethra alnifolia were taken from Zhang and Anderberg Citation(2002).
Results
General pollen morphology
In LM, pollen grains are in monads, oblate spheroidal to prolate in shape (P/E 0.95–1.48), 17.1–29.9 X 15.1–30.6 µm, 3‐ to 5‐colpor(oid)ate; commonly 3‐colpor(oid)ate in members of the sect. Enkiantella and Meisteria, 4‐colporate in the sect. Andromedina, and 4‐5‐colpor(oid)ate in the sect. Enkianthus, colpi 13.1–24.3 µm long, 0.7–2.3 µm wide, L/P 0.56–0.84, significantly wider at the middle, ora commonly distinct, but indistinct in E. deflexus (Yamazaki 2537), E. campanulatus var. longilobus, E. cernuus f. rubens, E. perulatus and E. serotinus (Tables & ). Colpi are acute towards the end in members of the sections Enkiantella and Meisteria, and slightly tapering to obtuse in members of the sections Andromedina and Enkianthus. Sometimes colpi are constricted at the middle, colpus margin distinct. Costae are distinct except in E. cernuus where they are indistinct. Ora are commonly lalongate, but circular in E. deflexus (Yamazaki 2537), E. campanulatus (Tatewaki et al. s.n.), E. cernuus f. rubens and E. sikokianus or not clear in E. perulatus and E. serotinus. Exine is tectate, apocolpial exine 1.0–2.6 µm thick and mesocolpial exine 1.1–2.2 µm thick. Usually the apocolpial exine is thicker than the mesocolpial exine, but thinner apocolpial exine has been observed in E. chinensis, one specimen of E. deflexus (Yamazaki 2537), E. campanulatus var. palibinii, E. sikokianus, and E. subsessilis or equal in thickness in one specimen of E. deflexus (Hara et al. 21810), E. cernuus, one specimen of E. cernuus f. rubens (Matsuda s.n.), and E. nudipes (Table ). Exine sculpture is finely verrucate to finely rugulate or verrucate to rugulate in most of the species except in E. cernuus, and E. nudipes, psilate to fine ornamentation.
In SEM, mesocolpial exine sculpture varies from finely verrucate‐rugulate to coarsely rugulate‐psilate with faint to distinct and minute to very minute granules (Figures ). Exine sculpture along the colpi is similar to mesocolpial exine. Colpi membrane is granulate, but smooth colpus membrane is also found in E. deflexus and E. campanulatus (Table ).
Table I. Pollen morphological data based on light microscopic investigation.
Table II. Variation in pollen characters of Enkianthus; showing mean value and standard deviation (minimum – maximum values in parenthesis).
Table III. Matrix of palynological characters and taxa. Pollen observation of Clethra alnifolia (Zhang & Anderberg, Citation2002).
Specific pollen description
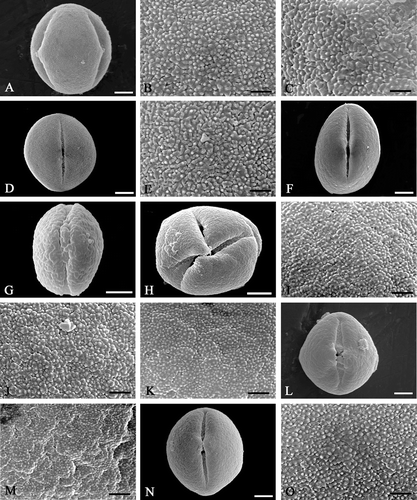
E. chinensis (Figure
Mesocolpial exine sculpture is finely verrucate‐rugulate (Figure ) with minute granules (d (diameter) <0.2 µm) or rarely moderately to coarsely rugulate with minute granules (d<0.2 µm). Apocolpial exine sculpture is finely verrucate‐rugulate (Figure ) with minute granules (d<0.2 µm).
E. deflexus (Figure
Pollen grains often shrink (Hara et al. 21810). Mesocolpial exine sculpture is finely rugulate (Figure ) with minute granules (d<0.15 µm) or rarely moderately to coarsely rugulate with minute granules (d<0.15 µm). Apocolpial exine sculpture is finely verrucate‐rugulate with minute granules (d<0.15 µm).
E. campanulatus (Figure
Only few pollen grains could be collected in one specimen (Sukawa s.n.). Pollen rarely syncolpate (see Figure ). Mesocolpial exine sculpture is finely verrucate‐rugulate (Figure ) with minute granules (d<0.2 µm) or finely to moderately verrucate‐rugulate (Figure ) with minute granules (d<0.15 µm). Few abnormal grains with coarsely rugulate exine sculpture, the rugulae transversely striate, are found in one specimen (Sukawa s.n.), (Figure ).
E. campanulatus var. longilobus (Figure
Pollen grains are often broken along colpus. Mesocolpial exine sculpture is moderately to coarsely rugulate (Figure ) with very minute granules (d<0.1 µm).
E. campanulatus var. palibinii (Figure
Mesocolpial exine sculpture is finely verrucate‐rugulate (Figure ) with minute granules (d<0.2 µm).
Figure 1 Scanning electron micrographs ofEnkianthus pollen. A–C. E. chinensis (Forrest 30465, TI). D–E. E. deflexus (Yamazaki 2537, TI). F–K. E. campanulatus: (F, J, & K) Takahashi 511, SAPS. (G & I) Sukawa s.n., SAPS. (H) Tatewaki et al. s.n., SAPS. L–M. E. campanulatus var. longilobus (Tashiro s.n.; TI). N, O. E. campanulatus var. palibinii (Tatewaki s.n.; SAPS). Pollen grains in equatorial view (A, D, F, G, L & N); syncolpate grain at polar view (H); micrographs with mesocolpial exine details (B, E, I, J, K, M & O); apocolpial exine (C). Scale bar – 5 µm (A, D, F–H, L, N); 1 µm (B, C, E, I–K, M, O).
E. cernuus (Figure
Pollen grains very often shrink. Both meso‐ (Figure ) and apocolpial (Figure ) exine sculptures are densely spaced granulate (granules d<0.25 µm).
E. cernuus f. rubens (Figure
Some pollen grains are circular at equatorial view. Mesocolpial exine sculpture is finely rugulate (Figure ) to scabrate (Figure ) with minute granules‐spinules (d<0.2 µm).
E. sikokianus (Figure
Some pollen grains are circular at equatorial view. Mesocolpial exine sculpture is moderately verrucate‐rugulate (Figure ) with minute granules (d<0.15 µm) or densely spaced granulate (Figure ) (granules d<0.25 µm). Apocolpial exine sculpture is sparsely spaced granulate (Figure ) with perforations (granules d<0.15 µm).
E. nudipes (Figures ,
Pollen grains amb is relatively angular, some grains circular at polar view. Mesocolpial exine sculpture is coarsely rugulate (Figure ) with dense minute granules (d<0.15 µm) to coarsely rugulate‐psilate without granules (Figure ). Apocolpial exine sculpture is coarsely rugulate (Figure ) with sparse and very minute granules (d<0.1 µm).
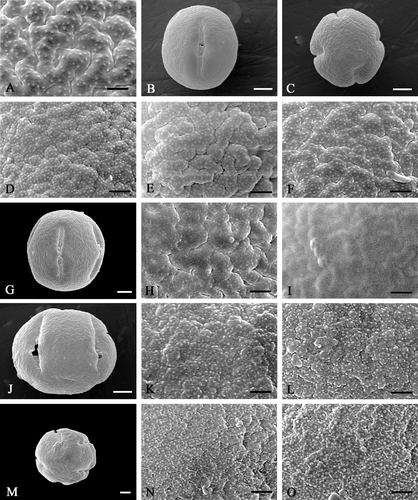
E. subsessilis (Figure
Pollen grains amb is relatively angular. Both meso‐ (Figure ) and apocolpial (Figure ) exine sculptures are coarsely rugulate with very minute granules (d<0.1 µm).
E. perulatus (Figure
Pollen grains are comparatively circular in polar view. Both meso‐ (Figure ) and apocolpial (Figure ) exine sculptures are coarsely rugulate‐psilate with faint very minute granules (d<0.1 µm) or mostly without granules. Size of the colpus granules is comparatively larger than other species (Figure ).
E. quinqueflorus (Figure
Pollen grains amb is relatively circular, most of the grains turning the polar face. Both meso‐ (Figure ) and apocolpial (Figure ) exine sculptures are moderately to coarsely rugulate with minute granules (d<0.15 µm).
E. serotinus (Figure
Both meso‐ (Figure ) and apocolpial (Figure ) exine sculptures are moderately to coarsely rugulate‐psilate with dense minute granules (d<0.15 µm).
Figure 2 Scanning electron micrographs ofEnkianthus pollen. A, B. E. cernuus (Takahashi 1827; SAPT). C–F. E. cernuus f. rubens: (C–E) M. Tohyama s.n., SAPS; (F) Matsuda s.n., SAPS. G–K. E. sikokianus: (G, H, J & K) Onooka 27245, SAPS; (I) Onooka 27206, SAPS. L–O. E. nudipes: (L, M & O) Onooka 27182, SAPS; (N) Nakajima s.n., SAPS. Pollen grains in equatorial view (C, G, L); pollen grain in polar view (H, M); details of mesocolpial exine (A, D‐F, I, J, N & O); apocolpial exine (B & K). Scale bar – 1 µm (A, B, D–F, I–K, N, O); 5 µm (C, G, H, L, M).
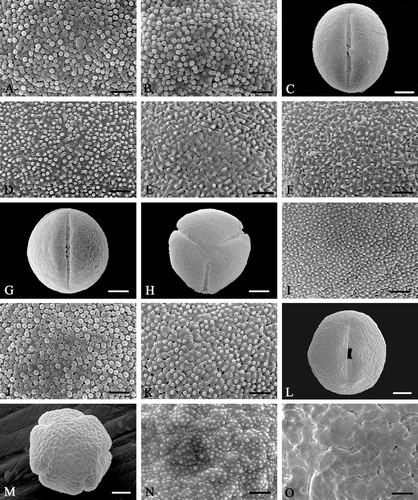
Figure 3 Scanning electron micrographs ofEnkianthus pollen. A. E. nudipes: Onooka 27182, SAPS; B–F. E. subsessilis: (B & D) Tohyama s.n., SAPS; (C, E & F) Ohashi et al. 11824, SAPT. G–I. E. perulatus: (G, H) Sarwar & Takahashi s.n., SAPS; (I) Takahashi 509, SAPS. J–L. E. quinqueflorus (McClure s.n.; TI). M–O. E. serotinus (Togashi & Murata 8032; TI). Pollen grains in equatorial view (B, G & J); pollen grain in polar view (C & M); details of mesocolpial exine (D, E, H, K & N); apocolpial exine (A, F, I, L & O). Scale bar – 1 µm (A, D–F, H, I, K, L, N); 5 µm (B, C, G, J, M).
Discussion
Distribution of palynological characters in Enkianthus
Apertures
Pollen aperture in Enkianthus vary from 3‐colpor(oid)ate to 5‐colpor(oid)ate. However, 3‐aperturate pollen is most common. Ueno Citation(1950) observed that aperture number in Enkianthus pollen varied from three to four, while 3‐ to 5‐aperturate pollen was observed by Ikuse (Citation1956, Citation2001), Kurosawa Citation(1991), Anderberg Citation(1994), and Zhang and Anderberg Citation(2002). Tricolpate pollen is the main and basic type found in most eudicots while other aperture types such as 5‐colpate, 6‐colpate, porate, colporate and pororate are regarded as derived among the eudicots (Walker & Doyle, Citation1975). Three‐aperturate pollen in the sections Enkiantella and Meisteria was shown by Anderberg Citation(1994) to be a plesiomorphic character and four to five apertures in the sections Andromedina and Enkianthus are more derived.
The aperture of the Enkianthus pollen is mainly colporate or colporoidate, but both types can be found in pollen of the same species; e.g., E. deflexus (Table ). Sometimes colpi are constricted at the middle and/or rarely pollen is syncolpate as in E. campanulatus, where all three colpi are fused at one pole (Fig. ). Granular colpus membrane in the outgroup and in most species of Enkianthus indicates that this character is symplesiomorphic while the smooth colpus membrane of E. campanulatus and E. deflexus is thought to have evolved in parallel in these two species.
The ratio of colpus length to polar axis (L/P) shows a distinct difference among the species (Table ). The L/P value is relatively higher, ranging from 0.69 to 0.84 in the sections Enkiantella and Meisteria, and relatively lower, ranging from 0.56 to 0.63 in the sections Andromedina and Enkianthus. The higher L/P value of the Enkiantella + Meisteria might be symplesiomorphic for the genus prevailing also in Clethraceae, and the lower L/P value of Andromedina + Enkianthus might be a synapomorphic character state like other characters of these sections (Anderberg, Citation1994).
Shape
Two distinct groups of pollen shape were found in Enkianthus (Table ). Species of sections Enkiantella and Meisteria have prolate spheroidal to prolate pollen (P/E 1.06–1.48) generally with relatively thinner exine. Species of sections Andromedina and Enkianthus typically have oblate spheroidal (P/E 0.95–0.97) pollen with relatively thicker exine, with the exception of E. nudipes (Table ), which has prolate spheroidal pollen (P/E 1.07). Prolate spheroidal pollen shape might be the plesiomorphic state while more derived states might have evolved independently; subprolate in E. campanulatus of sect. Meisteria; and oblate spheroidal in E. subsessilis of sect. Andromedina, and in sect. Enkianthus (Figure ). Another possibility is that the oblate spheroidal shape is synapomorphic for sects. Andromedina and Enkianthus, but revert to a plesiomorphic shape in E. nudipes of sect. Andromedina. It is possible that the oblate spheroidal pollen of the outgroup taxon Clethra alnifolia may have changed to the prolate spheroidal state in the ancestor of genus Enkianthus and evolved again to oblate spheroidal state. This supposition is supported by evolutionary trend of other characters viz., nos. 11, 14, 19 and 25, of Anderberg Citation(1994).
Size
All taxa examined in this study have pollen of almost similar size (24–30 µm) except E. cernuus, which has the smallest grains (17 µm) with thinner exine (Table ). Minute pollen might possibly represent a symplesiomorphic state and medium grains a synapomorphic state for section Enkianthus. However, like pollen shape, the medium pollen size could have evolved independently in E. campanulatus of sect. Meisteria and in E. nudipes of sect. Andromedina (Figure ) or be a synapomorphic state for in sects. Meisteria, Andromedina and Enkianthus, but reverts to a plesiomorphic state in E. cernuus of sect. Meisteria and E. subsessilis of sect. Andromedina.
Exine sculpture
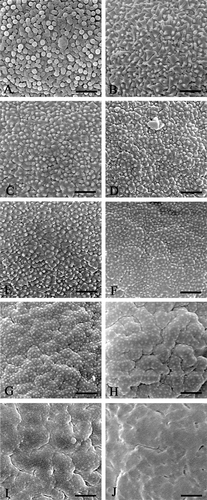
Exine sculptures within the genus show a more or less continuous and serial variation from finely verrucate‐rugulate to coarsely rugulate and coarsely rugulate‐psilate sculpture (Figures & ). Another trend in sculpture might be from finely verrucate‐rugulate (Figure ) to granulate and/or scabrate sculpture with granules‐spinules (Figure ). Since the variation in exine sculpture from psilate to finely verrucose has been observed in the genus Clethra, the synapomorphic state for the genus Enkianthus is not clear. Considering however, the evolutionary trend of other characters (Anderberg, Citation1994), the coarsely rugulate‐psilate sculpture might be the most specialized character state in the genus. Similarities and relationships between the different types of exine sculpturing are summarized in Figures and , and discussed in the latter part based on the infraspecific variation.
Figure 4 Morphological variation of mesocolpial exine sculpture inEnkianthus: (A) Granulate; (B) Scabrate with minute granules‐spinules; (C–E) Finely verrucate‐rugulate with minute granules; (F) Finely verrucate‐rugulate with minute granules; (G) Finely verrucate‐rugulate to moderately verrucate‐rugulate with minute granules; (H) Coarsely rugulate with minute granules; (I) Coarsely rugulate with faintly minute granules; (J) Coarsely rugulate‐psilate without granules. Scale bar – 1 µm (A–J).
Shrinkage
In E. cernuus and a Bhutanese specimen of E. deflexus (Hara et al. 21810), the pollen grains were very often shrunken and probably susceptible to acetolysis. Therefore, it was difficult to study these pollen grains with LM and SEM. Shrinkage might be due to poorly developed exine caused by genetic abnormalities and/or obstruction during pollen development process.
Taxonomic significance of pollen characters
Section Enkiantella
The sister‐group to all other species of Enkianthus (Anderberg, Citation1994). Pollen of Enkianthus chinensis and E. deflexus of this section are commonly 3‐aperturate, minute, prolate spheroidal in shape and exine sculpture finely verrucate‐rugulate to finely rugulate with minute granules. This result supports the earlier findings of 3‐colpor(oid)ate pollen for the two Enkianthus species in the LM study by Wang et al. Citation(1995). Both E. chinensis and E. deflexus are very similar to each other in external morphology and sometimes E. chinensis has been treated as a variety of E. deflexus (E. deflexus var. chinensis, Hara, Citation1966) and some plants with intermediate characters (e.g. leaf size, shape, indumentum etc.) have been observed (see Specimens Examined). Close similarity between the two species is also strongly supported by other pollen features (Figure ), but the investigation of further specimens is needed to clarify the status of these two taxa. The two taxa share several symplesiomorphic characters with E. campanulatus, which differs by having villous anther filaments and by lacking the densely lamellate seed surface (Anderberg, Citation1994). However, the two taxa share more ancestral palynological characters with E. cernuus than with E. campanulatus (Figure ). Infraspecific palynological variations in E. deflexus; i.e., 3‐colporoidate, minute, prolate spheroidal grains in cultivated Japanese specimen, 3‐(4)‐colporate, minute, subprolate grains in the Bhutanese specimen and 3‐(4)‐colporate, mediae, prolate spheroidal grains in the Chinese specimen, were found to correspond to geographical distribution (Table ). Geographical variation in the aperture number of Monotropa hypopitys was reported by Takahashi Citation(1987). Infraspecific difference of the pollen aperture number might be caused by the following reasons: i) ploidy level and/or pollen size (Lewis, Citation1964), ii) dimorphic flower (Kaplan & Mulcahy, Citation1971). Further study is necessary to clarify the infraspecific palynological variation in E. deflexus.
Section Meisteria
The sister‐group of the sections Andromedina and Enkianthus (Anderberg, Citation1994) includes the three Japanese species, E. campanulatus, E. cernuus and E. sikokianus. All three species have campanulate flowers arranged in racemose type of inflorescence, hairy pedicel, erect capsule, winged seeds, and a common type of pollen with three apertures. A wide inter‐ and infraspecific variation, both in apomorphic and plesiomorphic palynological characters, were, however, observed (Table & Figures , & ). Exine sculpture varies from finely verrucate‐rugulate to coarsely rugulate within the section. Infraspecific variation in exine sculpture occur; e.g. exine sculpture of E. campanulatus and E. campanulatus var. palibinii is finely verrucate‐rugulate (Figure ), but that of E. campanulatus var. longilobus is moderately to coarsely rugulate (Figure ). Pollen characters of E. campanulatus (e.g. 3‐(4)‐colpor(oid)ate grains, finely verrucate‐rugulate to moderately to coarsely rugulate exine sculpture) are similar to those observed for E. chinensis and E. deflexus of the section Enkiantella (Table ). Anderberg Citation(1994) also reported similarities among these three species. Few abnormal grains with coarsely rugulate exine sculpture with transversely striate rugulae were found in E. campanulatus (Figure ). Anderberg Citation(1994) considered E. campanulatus as a variable species; it includes a number of taxa earlier recognized as separate species, e.g. E. longilobus (Nakai) Ohwi, E. rubicandus Matsum. & Nakai, E. sikokianus (Nakai) Ohwi, E. kikuchi‐masaoi Mochizuki. The results of this palynological study do not support this circumscription of E. campanulatus. Enkianthus sikokianus, in particular, is distinct from the other taxa in having minute grains with thinner exine, mesocolpial exine sculpture with more densely spaced granules, and apocolpial exine sculpture with sparsely spaced granules and perforations (Figure ). When considering the differences in length of inflorescence and pedicels, as well as in number of flowers between E. sikokianus and E. campanulatus (Yamazaki, Citation1993), E. sikokianus should be recognized as a separate species. Similar infraspecific variation of exine sculpture was also found between E. cernuus (Figure ) and E. cernuus f. rubens (Figure ) and variation within the same taxa; E. campanulatus (Figure ), E. cernuus f. rubens (Figure ), E. sikokianus (Figure ) and E. nudipes (Figure ) respectively, was also observed. The palynological characteristics of E. cernuus f. rubens (e.g. indistinct ora, exine sculpture finely rugulate to scabrate with minute granules‐spinules) showed significant differences among the other members of this section (Table & Figure ).
Section Andromedina
Contains two species, E. nudipes and E. subsessilis, and is the sister‐group of section Enkianthus (Anderberg, Citation1994). Both species have racemose inflorescence, 4‐aperturate pollen, amb relatively angular, exine sculpture coarsely rugulate to coarsely rugulate‐psilate, smooth anther, urceolate corolla, glabrous pedicel, and wingless seeds. The exine of the pollen in species of Andromedina is somewhat thicker than that of other subgenera (Ueno, Citation1950). Enkianthus subsessilis and E. nudipes share many palynological characters with those of sect. Enkianthus, especially E. perulatus (Figure ). Enkianthus nudipes has 4‐(5)‐aperturate pollen, medium size, exine sculpture coarsely rugulate‐psilate and flowers with urceolate corolla, similar to those of E. perulatus (Table & Figures , ), and E. nudipes and E. perulatus could form a taxonomical bridge/link between the sections Andromedina and Enkianthus.
Pollen morphology has been studied for three species, E. perulatus, E. quinqueflorus and E serotinus, from the section Enkianthus (Anderberg, Citation1994). All taxa have more or less coriaceous leaves, and umbellate inflorescence. This section possesses the most stable palynological character states compared to those of other sections in the genus (Figure ). Pollen are medium sized, oblate spheroidal, 4‐5‐aperturate, amb relatively circular and exine sculpture moderately to coarsely rugulate‐psilate, and these characters might be more derived within the genus. The Chinese species, E. quinqueflorus and E. serotinus, are closely similar to each other in regards to external morphology and differ only in minor details (Hsu, Citation1982; Anderberg, Citation1994; Fang & Stevens, Citation2005). This is also supported by the pollen data. Enkianthus quinqueflorus and E. serotinus have flowers with a tubular corolla probably derived from ancestors with urceolate corollas, i.e. the condition in the E. perulatus, and in the section Andromedina. Anderberg Citation(1994) considered the section Enkianthus as a monophyletic specialized group within the genus, comprising all taxa with flowers in umbels, 4‐5‐aperturate pollen, and increasingly smooth anther.
Figure 5 Palynological data incorporated on phylogenetic tree ofEnkianthus by Anderberg Citation(1994); C. – Clethra; E. – Enkianthus. For character states see Table .
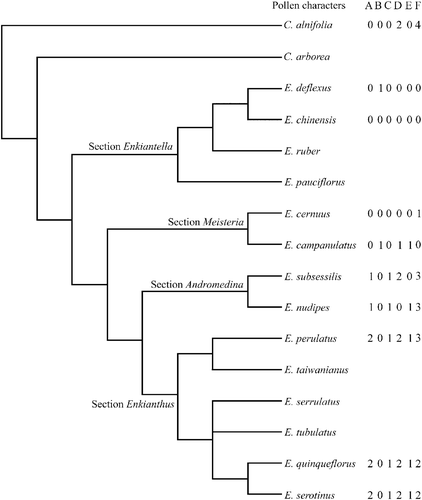
Phylogenetic significance of pollen characters
Ueno Citation(1950) observed 3‐aperturate pollen for E. campanulatus and E. cernuus from the section Meisteria, and Anderberg Citation(1994) reported 3‐aperturate pollen in both the sections Enkiantella and Meisteria. However, pollen with (2)‐3‐(4) apertures were found in the section Enkiantella and 3‐(4) apertures in the section Meisteria in this study (Table ), though the frequency of 2‐ or 4‐aperturate pollen grains was low. Anderberg Citation(1994) found pollen with frequent presence of five apertures only for one species, E. perulatus, in the section Enkianthus, as E. taiwanianus seems to be synonymous with E. perulatus (Li et al., Citation1998, p.17) while Ueno Citation(1950) did not observe any 5‐aperturate grains. According to our observation, 5‐aperturate pollen occurs commonly in section Enkianthus and few 5‐aperturate pollen are also found in E. nudipes of the section Andromedina. Furthermore, Ikuse Citation(2001) reported the occurrence of 5‐colporate grains in E. subsessilis of the section Andromedina. So the position of character state change (character 18.2) in Anderberg (Citation1994, Fig. 9 & App. 3) should be modified. Four apertures generally characterize the sect. Andromedina, and 4‐5 apertures characterize sect. Enkianthus.
No subdivision in pollen types corresponding to the sections could be identified. However, some morphological trends were noticed in pollen, in regards to aperture number, size, shape, and exine sculpture and based on pollen features two distinct groups are distinguished; “prolate 3‐aperturate pollen” characterizing the sections Enkiantella and Meisteria and the other, “oblate 4‐5‐aperturate pollen” characterizes the sections Andromedina and Enkianthus, (Table ). Pollen in sections Enkiantella and Meisteria is commonly 3‐aperturate, prolate spheroidal to prolate (P/E 1.06–1.48), L/P 0.69–0.84, with relatively thinner exine and exine sculpturing with a tendency to granulate. Pollen in the sections Andromedina and Enkianthus is 4‐5‐aperturate, commonly oblate spheroidal (P/E 0.95–0.97), L/P 0.56–0.63, with relatively thicker exine and exine sculpturing with a tendency to coarsely rugulate‐psilate with granules (Table ). However, it seems difficult to differentiate between sections of the same group on the basis of palynological characters only.
The species of sections Enkiantella and Meisteria have plesiomorphic characters such as racemose inflorescence, campanulate and actinomorphic flower, and 3‐aperturate pollen grains. Anderberg Citation(1994) concluded that the ancestors of Ericaceae were similar to Enkianthus chinensis or E. campanulatus in many respects. Therefore, 3‐aperturate, minute, and prolate spheroidal to prolate pollen grains with finely verrucate‐rugulate exine sculpture with minute granules may possibly be a plesiomorphic character in the genus. The species of the section Enkianthus, on the other hand, have umbellate inflorescence and 4‐5‐aperturate, medium, and oblate spheroidal pollen with exine sculpture coarsely rugulate with minute to mostly without granules, and are regarded as advanced. Anderberg Citation(1994) found a similar trend, whereby racemes evolved into umbels in the inflorescence of Enkianthus.
Evidence suggests that the major trend in exine sculpture is from finely verrucate‐rugulate through coarsely rugulate to coarsely rugulate‐psilate with distinct to faint granules in the genus Enkianthus (Figure ). However, since a wide infrageneric variation of exine sculpture is observed in the Ericaceae and related groups; e.g. psilate to verrucose in (Zhang & Anderberg, Citation2002), we have to consider the character states of exine sculpture very carefully.
It is interesting that by adding palynological characters to the phylogenetic analysis of Anderberg Citation(1994), the tree topology was generally changed within the sections Enkiantella and Meisteria, while no change was observed in the Andromedina + Enkianthus clade. This might be due to more stable character states in the Andromedina + Enkianthus clade. Species composition of sect. Meisteria might require further research including the study of more specimens and combined analysis of morphological and molecular data in order to clarify the validity and systematic position within the genus Enkianthus.
Dichotomous key to pollen of the Enkianthus taxa examined:
-
1a. Pollen grains commonly 3‐aperturate, colpi acute towards end, P/E 1.06–1.48, L/P 0.69–0.84, sculpture tending to granulate ‐‐‐‐‐‐‐‐‐‐ 2
-
2a. Sculpture fine verrucate‐rugulate or moderate to coarse rugulate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 3
-
3a. (2)‐3‐colporate, endoaperture lalongate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ E. chinensis
-
3b. 3‐(4)‐colpor(oid)ate, endoaperture circular/lalongate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ E. deflexus
-
2b. Sculpture fine verrucate‐rugulate to coarse rugulate‐psilate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 4
-
4a. Sculpture granulate to coarse rugulate‐psilate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 5
-
5a. Sculpture granulate to Scabrate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 6
-
6a. Aperture colporate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 7
-
7a. Prolate spherical, endoaperture lalongate ‐‐‐‐‐ E. cernuus
-
7b. Subprolate, endoaperture circular ‐‐‐‐‐‐‐‐‐ E. sikokianus
-
6b. Aperture colpor(oid)ate‐‐‐‐‐‐‐‐‐‐ ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ E. cernuus f. rubens
-
5b. Sculpture moderate rugulate to coarse rugulate‐psilate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 8
-
8a. Coarse rugulate‐psilate, subprolate ‐‐‐‐‐‐‐ E. campanulatus var. palibinii
-
8b. Moderate to coarse rugulate, prolate spheroidal ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ ‐‐‐‐‐‐E. campanulatus var. longilobus
-
4b. Sculpture fine verrucate‐rugulate to coarse rugulate, subprolate/prolate, endoaperture lalongate/circular ‐‐‐‐‐‐‐‐‐‐ E. campanulatus
-
1b. Pollen grains 4‐5‐aperturate, colpi slightly tapering or obtuse towards end, P/E 0.95–1.07, L/P 0.56–0.63, sculpture tending to coarse rugulate‐psilate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 9
-
9a. Commonly 4‐aperturate, amb relatively angular, aperture colporate ‐‐‐‐‐‐‐‐‐‐‐‐‐‐ 10
-
10a. Mediae, prolate spheroidal ‐‐‐‐‐‐ ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ E. nudipes
-
10b. Minute, oblate spheroidal ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ E. subsessilis
-
9b. Commonly 4‐5‐aperturate, amb relatively circular, aperture colpor(oid)ate ‐‐‐‐‐‐‐‐ 11
-
11a. Aperture colporate, endoaperture lalongate ‐‐‐‐‐‐‐‐‐‐‐‐‐ E. quinqueflorus
-
11b. Aperture colpor(oid)ate, endoaperture lalongate/circular ‐‐‐‐‐‐‐‐‐‐‐‐‐ 12
-
12a. Sculpture moderate to coarse rugulate with minute granules ‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐ E. serotinus
-
12b. Sculpture coarse rugulate with faint very minute or without granules ‐‐‐‐‐‐‐‐‐‐‐‐ E. perulatus
Conclusions
Our observations in Enkianthus pollen show that the pollen is monadinous, oblate spheroidal to prolate in shape, 3‐ to 5‐colpor(oid)ate and exine sculpture varies from granulate to coarsely rugulate‐psilate, with faint to distinct minute granules. It is concluded that taxa having 3‐aperturate, minute, and prolate spheroidal pollen in the sections Enkiantella and Meisteria are probably plesiomorphic, while taxa with 4‐5‐aperturate, medium, and oblate spheroidal pollen in the sections Andromedina and Enkianthus might be more derived within Enkianthus. A close relationship between E. chinensis and E. deflexus is strongly supported by palynological characters. The Andromedina + Enkianthus clade is also supported as a monophyletic group with shared derived palynological characters. Among the infrageneric classifications, Anderberg’s system best suits the present result.
Specimens investigated
∗ = Light microscope; ∗∗ = Scanning electron microscope; • = Taxa with intermediate morphological characters.
Section Enkiantella Palibin
E. chinensis Franch. Flora of S.W. CHINA: NE Upper Burma and SE Tibet (Ex. Herb.Hort. Bot. Reg. Edin), no date. Forrest 30465 (TI); (∗, ∗∗). C China, W Hukeh, no date. E. H. Wilson 1002 (E 00201100); (∗∗).
E. deflexus (Griff.) Schn. JAPAN: Tokyo, cultivated, 24.05.1981. T. Yamazaki 2537 (TI 1326237); (∗, ∗∗). BHUTAN: Tzatogang (3200 m)–Dotanang (2500 m), 27.05.1967. H. Hara et al. 21810 (TI); (∗). CHINA: W. Yunnan, Huadianba Farm, W of N end of Cangshan, alt. 2900 m. 19.05.1981. Unknown 0814 (E 00201099)•; (∗, ∗∗). NW Yunnan, Mountain of Wei‐Hsi, alt. 3080 m, no day. 07.1928. J. F. Rock 17142 (E 00201101)•; (∗∗).
Section Meisteria Palibin
E. campanulatus (Miq.) Nichols. JAPAN: Sendai‐shi, 24.05.1977. H. Takahashi 511(SAPS); (∗, ∗∗). Honshu, Kaga, Mt. Hakusan, no day. 07.1889. C. Sukawa s.n. (SAPS); (∗, ∗∗). Honshu, Shinano, near Omagari‐Zorin‐Zigyosyo, cultivated plant, 10.06.1961. M. Tatewaki, K. Ito et. T. Nigi s.n. (SAPS); (∗, ∗∗). Sapporo‐shi, Hokkaido University, Botanic Garden, cultivated, 02.06.2004. Sarwar & H. Takahashi s.n. (SAPS); (∗∗).
E. campanulatus (Miq.) Nichols var. longilobus (Nakai) Makino. JAPAN: Bungo, Mt. Kuju, 11.06.1928. Z. Tashiro s.n. (TI); (∗, ∗∗).
E. campanulatus (Miq.) Nichols var. palibinii (Craib) Bean. JAPAN: Honshu, Shimotsuke, Nikko, the pass of Sanno, 23.06.1961. M. Tatewaki s.n. (SAPS); (∗, ∗∗).
E. cernuus (Sieb. et Zucc.) Makino. JAPAN: Shikoku, 21.6.1984. M. Takahashi 1827 (SAPT); (∗, ∗∗).
E. cernuus (Sieb. et Zucc.) Makino f. rubens (Maxim.) Ohwi. JAPAN: Honshu, Kanagawa, Hakone, Mt. Kami‐Futago, 04.06.1967. M. Tohyama s.n. (SAPS); (∗, ∗∗). Honshu, Chichibu‐Musashi, 17.05.1901. S. Matsuda s.n. (SAPS); (∗, ∗∗). Honshu, Kii, Koyasan, 09.06.1929. K. Numajiri s.n. (SAPS); (∗∗). Honshu, Wakayama, Kozagawa‐cho, Hirai, 20‐24.05.1970. T. Kumata s.n. (SAPS); (∗∗).
E. sikokianus (Palib.) Ohwi. JAPAN: Honshu, Kii, Upper Koga, Tamanotami, 13.05.1937. T. Onooka 27206 (SAPS); (∗, ∗∗). Honshu, Wakayama Exp. For., 03.06.1937. T. Onooka 27245 (SAPS); (∗∗).
Section Andromedina Palibin
E. nudipes (Honda) Ohwi. JAPAN: Honshu, Wakayama Exp. For. 03.05.1937. T. Onooka 27182 (SAPS); (∗, ∗∗). Honshu, Kii, Mt. Meshimori, 30.04.1928. T.Nakajima s.n. (SAPS); (∗∗).
E. subsessilis (Miq.) Makino. JAPAN: Honshu, Utsunomiya–shi, 28.05.1982. H. Ohashi et al. 11824 (SAPT); (∗, ∗∗). Honshu, Kodzuke, Mt. Tanigawa, 21.06.1956. M. Tohyama s.n. (SAPS); (∗∗). Honshu, Nikko, Yagenbori, 07.07.1903. H. Takeda s.n. (SAPS); (∗∗).
Section Enkianthus (Section Euenkianthus Palibin)
E. perulatus (Miq.) C.K. Schn. JAPAN: Sendai‐shi, cultivated, 02.05.1977. H. Takahashi 509 (SAPS); (∗, ∗∗). Sapporo‐shi, Hokkaido University, Botanic Garden, cultivated, 18.05.2004. Sarwar & H. Takahashi s.n. (SAPS); (∗∗).
E. quinqueflorus Lour. CHINA: Flora of Kwangtung, Herb. Canton Christian, Coll. No. 13271, 13.03.1967. F. A. McClure s.n. (TI); (∗, ∗∗).
E. serotinus Chun & Fang. CHINA: Kwangtung, Chiow‐lung, Ta‐mao‐shan, alt. 300–700 m. 16.01.1964. M. Togashi & G. Murata 8032 (TI); (∗, ∗∗).
Acknowledgements
Authors wish to express their sincere thanks to the Directors and Curators of Herbaria: E, SAPS, SAPT and TI for allowing us to examine and/or send the specimens on loan and sample polliniferous materials. Particular thanks are due to Ms. Aoyama Yuriko and Mr. Toshiaki Ito of Electron Microscopy Laboratory, Graduate School of Agriculture, Hokkaido University for their technical assistance and cooperation during the SEM study and photography of pollen grains. The first author (A. K. M. G. S.) is thankful to Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan for the Monbukagakusho Scholarship during the period of this study. We appreciate the help of Prof. Dr Else Marie Friis; Julie Cantrill, and two anonymous reviewers for their linguistic check and many helpful comments on the manuscript.
References
- Anderberg , A. A. 1993 . Cladistic interrelationships and major clades of the Ericales. . Pl. Syst. Evol. , 184 : 207 – 231 .
- Anderberg , A. A. 1994 . Cladistic analysis of Enkianthus with notes on the early diversification of the Ericaceae. . Nord. J. Bot. , 14 : 385 – 401 .
- Anderberg , A. A. , Rydin , C. and Källersjö , M. 2002 . Phylogenetic relationships in the order Ericales s.l.: analysis of molecular data from five genes from the plastid and mitochondrial genomes. . Am. J. Bot. , 89 : 677 – 687 .
- Erdtman , G. 1960 . The acetolysis method. A revised description. . Sven. Bot. Tidskr. , 54 : 561 – 564 .
- Erdtman , G. 1986 . Pollen morphology and plant taxonomy. Angiosperms , Leiden : E. J. Brill .
- Fang , R. and Stevens , P. F. 2005 . “ Enkianthus ” . In Flora of China Edited by: Wu , Z and Raven , P. H . 243 – 245 . Vol. 14 (Apiaceae through Ericaceae) Beijing: Science Press & St. Louis, MO: Mo. Bot. Gard. Press
- Hara , H. 1948 . Enumeratio Spermatophytarum Japonicarum , 1. Tokyo : Iwanami Shoten .
- Hara , H. 1966 . “ Taxonomic comparison between corresponding taxa of spermatophyta in eastern Himalaya and Japan. ” . In The flora of Eastern Himalaya , Edited by: Hara , H . 627 – 657 . Tokyo : Univ. Tokyo .
- Holmgren , P. K. , Holmgren , N. H. and Barnett , L. C. 1990 . Index Herbariorum, Part I: The Herbaria of the World , 8 , Bronx, NY : N. Y. Bot. Gard .
- Hsu , T. Z. 1982 . Classification, distribution and phylogeny of the genus Enkianthus. . Acta Bot. Yunn. , 4 : 355 – 362 . (in Chinese with English abstract)
- Ikuse , M. 1956 . Pollen grains of Japan , Tokyo : Hirokawa Publ. Co . (in Japanese)
- Ikuse , M. 2001 . Pollen grains of Japan , 2 , Tokyo : Hirokawa Publ. Co . (in Japanese)
- Judd , W. S. and Kron , K. A. 1993 . Circumscription of Ericaceae (Ericales) as determined by preliminary cladistic analyses based on morphological, anatomical and embryological features. . Brittonia , 45 : 99 – 114 .
- Kaplan , S. M. and Mulcahy , D. L. 1971 . Mode of pollination and floral sexuality in Thalictrum. . Evolution , 25 : 659 – 668 .
- Kron , K. A. 1996 . Phylogenetic relationships of Empetraceae, Epacridaceae, Ericaceae, Monotropaceae, Pyrolaceae: Evidence from nuclear ribosomal 18s sequence data. . Ann. Bot. (London) , 77 : 293 – 303 .
- Kron , K. A. 1997 . Phylogenetic relationships of Rhododendroideae (Ericaceae). . Am. J. Bot. , 84 : 973 – 980 .
- Kron , K. A. and Chase , M. 1993 . Systematics of the Ericaceae, Empetraceae, Epacridaceae and related taxa based upon rbcL sequence data. . Ann. Mo. Bot. Gard. , 80 : 735 – 741 .
- Kron , K. A. , Judd , W. S. , Stevens , P. F. , Crayn , D. M. , Anderberg , A. A. , Gadek , P. A. , Quinn , C. J. and Luteyn , J. L. 2002 . Phylogenetic classification of Ericaceae: molecular and morphological evidence. . Bot. Rev. , 68 : 335 – 423 .
- Kurosawa , K. 1991 . SEM photographs of pollen of angiosperms , Osaka : Osaka Mus. Nat. Hist . (in Japanese)
- Lewis , W. H. 1964 . Oldenlandia corymbosa (Rubiaceae). . Grana Palynol. , 5 : 330 – 341 .
- Li , H. L. , Lu , S. Y. , Yang , Y. P. and Tseng , Y. H. 1998 . “ Ericaceae. ” . In Flora of Taiwan , Edited by: Huang , T. C , Li , H.‐L , Liu , T.‐S , Koyama , T and de Vol , C. E . Taipei : Dept. Bot. Natl Taiwan Univ . Vol. 4 (2nd ed.)
- Nakamura , J. 1980 . Diagnostic characters of pollen grains of Japan , Vol. I & II , Osaka : Osaka Mus. Nat. Hist . (in Japanese)
- Palibin , J. V. 1899 . Revisio generis Enkianthus Lour. . Scripta Bot. Horti Univ. Imp. Petropolit. , 15 : 1 – 18 .
- Punt , W. , Blackmore , S. , Nilsson , S. and Le Thomas , A. 1994 . Glossary of pollen and spore terminology , Utrecht : LPP Found . LPP Contrib. Ser. 1
- Takahashi , H. 1987 . Pollen morphology and its taxonomic significance of the Monotropoideae (Ericaceae). . Bot. Mag. Tokyo , 100 : 385 – 405 .
- Ueno , J. 1950 . On Enkianthus: A classification of the genus Enkianthus based upon the characters of pollen grains and crystals in the leaf. . J. Inst. Polytechn. Osaka City Univ. Ser. D , 1 : 55 – 62 .
- Walker , J. W. and Doyle , J. A. 1975 . The bases of angiosperm phylogeny: Palynology. . Ann. Mo. Bot. Gard. , 62 : 664 – 723 .
- Wang , F. , Chien , N. , Zhang , Y. and Yang , H. 1995 . Pollen flora of China , 2 , Beijing : Science Press . (in Chinese)
- Yamazaki , T. 1993 . Enkianthus, Flora of Japan , Edited by: Iwatsuki , K , Yamazaki , Y , Boufford , D. E and Ohba , H . Vol. IIIa , 45 – 47 . Tokyo : Kodansha .
- Zhang , X. P. and Anderberg , A. A. 2002 . Pollen morphology in the ericoid clade of the order Ericales, with special emphasis on Cyrillaceae. . Grana , 41 : 201 – 215 .