Abstract
A morphological study of three different types of dispersed Circumpolles pollen grains from the Toarcian‐Aalenian of the Fuentelsaz section (Spain) was performed using conventional wide field and confocal laser scanning microscopy (CSLM). The CSLM data allowed scanning electron microscope‐like three‐dimensional reconstructions to be made and showed that the most part of the studied pollen have taxonomic features characteristic of the Mesozoic genus Classopollis. At the species level, Classopollis torosus and Classopollis classoides were identified. The third form studied is characterized by the presence of Ubisch bodies adhering to the pollen wall.
A confocal laser scanning and conventional wide field light microscopy study of Classopollis from the Toarcian‐Aalenian of the Fuentelsaz section (Spain)
Palynological studies have often relied on the use of techniques based on wide field light microscopy, electron microscopy (scanning electron microscopy [SEM] or transmission electron microscopy [TEM]), and more recently, confocal laser scanning microscopy (CSLM) (Salih et al., Citation1997). The technique used depends on the aim pursued, the time available, and the material to be examined. Despite its limitations, conventional light microscopy remains the basic tool used in palynological investigations. Electron microscopy affords the highest resolution (Taylor, Citation1999), but the destruction of material analysed using TEM and the time needed to perform ultrastructural studies has led some researchers to use alternative techniques such as CSLM (Shute et al., Citation1996; Feist‐Burkhardt & Pross, Citation1999; Hochuli & Feist‐Burkhardt, Citation2004). The latter is a powerful imaging technique that allows high resolution optical sections and three‐dimensional SEM‐like reconstructions of small‐scale samples to be made. Previous micropalaeontological studies have highlighted the interest and promise of CSLM for morphological analysis in a variety of organisms including ostracods (Birkmann & Lundin, Citation1996), radiolarians (O'Connor, Citation1996), acritarchs (Yang et al., Citation1998) and dinoflagellate cysts (Feist‐Burkhardt & Pross, Citation1999). Palynological studies of terrestrial palynomorphs, the first of which were performed by Scott and Hemsley Citation(1991) and Salih et al. Citation(1997), and more recently by Banks et al. Citation(2006), have provided interesting results.
This paper reports a detailed study of Toarcian‐Aalenian Circumpolles pollen grains including Classopollis from the Fuentelsaz section (Spain) using CSLM and wide field light microscopy. The genus Classopollis Pflug, Citation1953 emend. Reyre, Citation1970 nom. cons., non Malyavkina, Citation1949 (Traverse, Citation2004; Skog, Citation2005) is a well‐known Mesozoic pollen type that was produced by conifers of the extinct family Cheirolepidiaceae (Taylor & Alvin, Citation1984). The genus was initially instituted by Pflug Citation(1953) and later emended by Pocock and Jansonius Citation(1961) and Reyre Citation(1970). As currently circumscribed the genus is defined by the presence of a distal pore, a proximal trilete scar, a two‐layered exine, and an equatorial band (not necessary thickened) with or without striations and allows for a considerable variation in sculptures and structures. The approach of Reyre Citation(1970) allows the inclusion of forms of pollen with morphological variations, as well as others, which do not show the full set of features characterising the genus. For example, it allows species with sculptural processes, such as Classopollis echinatus Burger, Citation1966, or Classopollis caratinii Reyre, Citation1970, to be included in the genus. However, despite the abundance of Classopollis in Mesozoic rocks and the large number of taxonomic studies dealing with its complex and varying morphology (e.g., Pocock & Jansonius, Citation1961; Burger, Citation1965, Citation1966; Médus & Reyre, Citation1966; Boltenhagen, Citation1968, Citation1973; Reyre, Citation1970; Taugourdeau‐Lantz, Citation1971; Pocock et al., Citation1990), it often remains difficult to identify its member species.
Given the particular morphological features shown by Classopollis pollen species, their important role in Mesozoic assemblages, and their abundance in the Mesozoic rocks of the Iberian Peninsula (Gomez et al., Citation2001; Barrón & Azêredo, Citation2003; Barrón et al., Citation1999, Citation2006), Classopollis is ideal for investigating the potential of CSLM as tool in palynological analyses. Special interest was paid to pollen ultrastructure reported in SEM and TEM studies (e.g., Pettitt & Chaloner, Citation1964; Reyre, Citation1970; Srivastava, Citation1976; Taylor & Alvin, Citation1984). This paper is the first to report CSLM observations of Classopollis pollen grains.
Material and methods
Geological setting
The Fuentelsaz section, recently defined as the Global Boundary Stratotype Section and Point (GSSP) of the Toarcian–Aalenian Boundary (Cresta et al., Citation2001), is a site of special interest in the central sector of the Rama Castellana (Castilian Branch) of the Iberian Range (Figure ). It is situated about 0.5 km north of the village of Fuentelsaz in the Province of Guadalajara, Spain and was described by Goy et al. Citation(1994, Citation1996), and Cresta et al. Citation(2001). Compared to other sections of the Iberian Range, this section is well‐developed (it is some 41.5 m thick) and shows remarkable continuity of Toarcian‐Aalenian deposits, which are composed of marls with interbedded limestones forming irregular rhythmic alternations. The section includes the upper part of the Turmiel Formation (Goy et al., Citation1976) (levels 1 to 153), and the basal portion of the Casinos Formation (Gómez & Goy, Citation1979) (levels 154 to 205). Together, these deposits represent the transgressive and regressive depositional cycle LJ‐4 described by Gómez & Goy Citation(2005).
Figure 1 Geological scheme of the Mesozoic outcrops in the Fuentelsaz area(FZ) and the location of the sites mentioned in the text.
The Upper Toarcian and Lower Aalenian rocks of the Fuentelsaz section yield abundant remains of marine and continental fossils such as brachiopods, bivalves, foraminifera, ostracods, palynomorphs and calcareous nanofossils, including a well‐preserved succession of ammonoid assemblages (Goy et al., Citation1994, Citation1996; Cresta et al., Citation2001).
Sample preparation and microscopy protocols
Samples were prepared by acid treatment (HCl, HF) at high temperature (Wood et al., Citation1996) and mounted on microscope slides using glycerine jelly. Due to the autofluorescence properties of palynomorphs, the samples were not stained with a signal‐enhancing agent (Gijzel, Citation1967). The palynological content of 69 stratigraphic levels were investigated by transmitted light wide field microscopy using a Leica Laborlux D microscope coupled to a Wild MPS45 camera.
For the CSLM study, selected sporomorphs were studied using a Biorad 1024 CSLM system [Zeiss Advanced Imaging Microscopy (A. I. M.), Jena, Germany] coupled to a Nikon microscope and driven by Leica Confocal Scanware software, at the Centre for Microscopy and Cytometry of the Complutense University of Madrid (Spain). An oil immersion objective with magnification of ×63 and a numerical aperture of 1.4 was used. The excitation source was a helium/neon mixed gas laser emitting at 488, 594 and 633 nm coupled to an acousto‐optical beam splitter system and a prism spectrophotometer detector. The channel providing the best result for each palynomorph was selected and the signal‐to‐noise ratio improved by time averaging over eight repetitions for each optical section. Each of the latter had a size of 512×512 pixels; each pixel was associated with an 8‐bit intensity grey value ranging from 0 to 255. The distances between each section were chosen taking into account the size of the specimens and the required level of precision (0.2–0.5 µm). VG Studio Max 1.2 software (Volume Graphics GmbH, Heidelberg, Germany) was used to process the image stacks and to generate three‐dimensional (3D) renderings of the whole grains.
Results
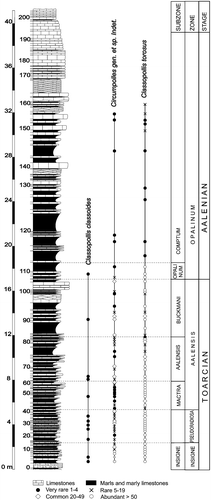
Pollen grains belonging to the genus Classopollis were abundant throughout the Fuentelsaz section (Figure ), and were predominant in the assemblages corresponding to the Upper Toarcian rocks of the Insigne Zone (levels FZ1 to FZ16).
Figure 2 Stratigraphic succession of the Fuentelsaz section and the relative distribution of theClassopollis species.
The use of CSLM plus conventional wide field microscopy allowed three major types of Circumpolles including Classopollis to be distinguished:
• Type A. Pollen grains mostly observed individually, circular in polar view (Figure ), and more or less elliptic in equatorial view (Figure ); equatorial diameter (E): 20–32 µm and length of the polar axis (P): 18–21 µm. Proximal side with a small trilete mark sometimes forming an ill‐defined sinuous fold or a small triangular area. Distal hemisphere flattened; a marked circular pore (Figure ) visible in polar view. Exine of both hemispheres 1–2 µm thick with psilate ornamentation and an alveolate structure. Equatorial band 2–3 µm thick, with ill‐defined endostriae running parallel around the equatorial plain. Hemispheres clearly separated by a pronounced rimula.
CSLM clearly shows the structure of the pollen wall (Figure ). The diameter and length of the elements making up the sexine are smaller around the distal pore and at the level of the rimula (Figure ); at the equatorial band they are distinctly larger. In tangential view the anastomosing system and the structure of the endostriae are revealed (Figure ). The confocal sections allow specification of variations in the pollen wall thickness: 0.6–1 µm at the distal pole area and rimula, 1.2–2.2 µm over both hemispheres, and 2.1–2.7 µm at the equatorial girdle. In the 3D rendering (Figure ) and CSLM maximum projection of the sections (Figure ), the psilate nature of the sculpture is distinct. Hairy strands arising from the proximal polar area (Figure ) have been observed on some specimens.
These grains appear sporadically throughout the Upper Toarcian and lowermost Aalenian (Opalinum Zone).
• Type B. Pollen grains frequently observed in tetrads (Figure ). Monads oblate, and more or less elliptical in equatorial view; Equatorial diameter (E): 26–29 µm and length of the polar axis (P): 20–26 µm; spherical, distally monoporate, and with a trifid tetrad scar in the proximal polar area. Exine scabrate, thicker in the equatorial area than in the polar area, and forming a girdle with 5–10 endostriae (Figure ). Rimula clearly visible (Figure ).
Rotation of the specimens by CSLM revealed the distal pore and the proximal scar on the same folded side of the studied specimen (Figure ). In optical sections the exine varies in thickness from 0.8 to 1.3 µm. As in the case of Type A specimens, the structure of the sexine appeared alveolate.
Type B is the most abundant form of Circumpolles pollen in the Fuentelsaz section; it is particularly common in the late Toarcian assemblages corresponding to the Insigne Zone (levels FZ1 to FZ16) where it is numerically dominant. It is also well represented with percentages of more than 30% in the Pseudoradiosa Zone (levels FZ17 to FZ42), and in the lower and middle part of the Aalensis Zone (levels FZ43 to FZ77 and FZ103 to FZ105). It is also a conspicuous component of the palynological assemblages of the lowermost Aalenian (Opalinum Zone, levels FZ107 to FZ111).
The descriptions of the pollen grains attributed to Type A and B based on conventional light microscopy and CSLM agree with the emended diagnosis of Classopollis classoides (Pflug, Citation1953) Pocock & Jansonius, Citation1961 and Classopollis torosus (Reissinger, Citation1950) Balme, Citation1957, respectively. The precise, systematic separation of these species remains controversial. Authors such as Couper Citation(1958) and Cornet and Traverse Citation(1975) suggested that Classopollis torosus should be treated as synonymous with Classopollis classoides. However, Pocok and Jansonius Citation(1961), Burger Citation(1965, Citation1966) and Srivastava Citation(1976) consider the apparent resemblance between these taxa to be insufficient to affirm their synonymy since the loss of the type material precludes any precise and objective comparison. The synonymy of these two species proposed by Couper Citation(1958) and Cornet and Traverse Citation(1975) is thus still uncertain. Our results suggest, however, that CSLM is a useful tool for investigating the similarities and differences of these taxa.
The CSLM study confirmed, in both Type A and B pollen, the presence of the seven features indicated by Pocock and Jansonius Citation(1961) for Classopollis classoides: (a) a circular distal pore (b) a proximal trilete scar, (c) a two‐layered exine including (d) a spheroid nexine (continuous over whole area of the pollen grain) and (e) a variable and more complex sexine some 1–2 µm thick showing (f) a rimula and (g) an equatorial thickening differentiated into “four or more annular bands … which may or may not be quite pronounced”. However, the Type B grains showed a thinner scabrate exine than those of type A, relating them more to Classopollis torosus (Reissinger, 1959) Balme, Citation1957.
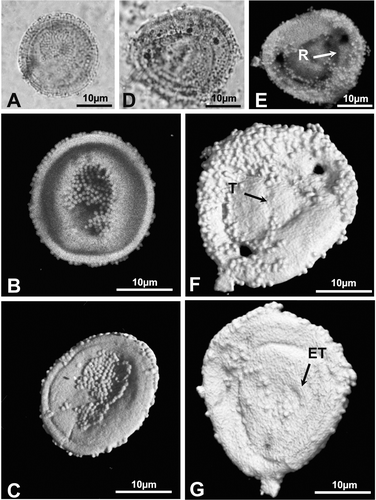
• Type C. Pollen grains observed individually (Figure ) or as tetrads (Figure ). Monads, oblate to spherical, in polar view circular to elliptic, in equatorial view elliptical; Equatorial diameter (E): 22–31 µm, length of the polar axis (P): 20–28 µm; the polar areas are usually flattened at the poles. Distally monoporate with a proximal scar that is not always very pronounced (Figure ). Hemispheres separated by a well defined rimula (Figure ). Exine two‐layered and equally thick in all areas; nexine thin and quite homogeneous. The exine is scabrate, alveolate, 0.6–1.3 µm thick and covered by abundant Ubisch bodies, 0.6–1.4 µm in diameter, either isolated or in groups (Figure ).
The equatorial thickening (1.2–1.5 µm) showed no visible striations. The distal operculum‐like structure, up to 7 µm, appears sometimes bordered by an exinal thickening (Figure ).
This type of pollen grains occurs throughout the section, and is abundant in the Insigne Zone (levels FZ1 to FZ16). Above the Pseudoradiosa Zone, it gradually decreases in abundance and is very rare in the Aalenian.
Figure 3 A–H. Type A: Classopollis classoides. (A) Light microscope (LM) photomicrograph of an individual pollen grain of C. classoides in polar view; (B) Confocal scanning laser microscope (CSLM) photomicrograph of the same grain obtained by maximum projection of the optical slice stack along the vertical axis. Note that the white spots (black arrow) correspond to sexine elements; (C) LM photomicrograph of another specimen of C. classoides in equatorial view; (D & E). CSLM partial maximum projection of sections of the same grain as in (C). Each composite view represents a 0.8 µm‐thick section of the grain. Note the anastomosing system (black arrow) formed by the sexine elements; (F) CSLM full maximum projection of the same grain shown in (C). Note the well defined rimula and equatorial endostriae; (G) CSLM detail of the same grain as in (C). Note the thinning of the sexine at the level of the rimula (arrow). (H) Three‐dimensional (3‐D) rendering based on CSLM sections of the same grain as in (C). Note the psilate nature of the sculpture and the emerging lips of the trilete scar.
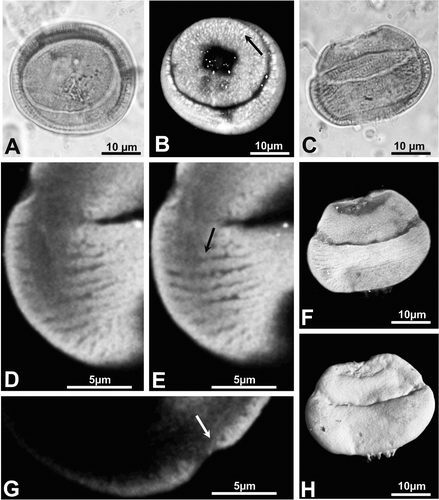
Figure 4 A, B & D, E. Type B: Classopollis torosus. C. Type C: Circumpolles gen. et sp. indet. (R – rimula; T – trilete scar; P – distal pore). (A) LM photomicrograph of a tetrad of pollen grains attributed to C. torosus; (B) LM photomicrograph of a single pollen grain of C. torosus; (C) LM photomicrograph of a tetrad of pollen grains attributed to Circumpolles gen. et sp. indet.; (D) 3‐D rendering based on CSLM sections of the same grain as in (B). Note the scabrate nature of the sculpture; (E) 3‐D rendering based on CSLM sections of the same grain as in (B) but showing its back side. Note the pore and trilete scar.
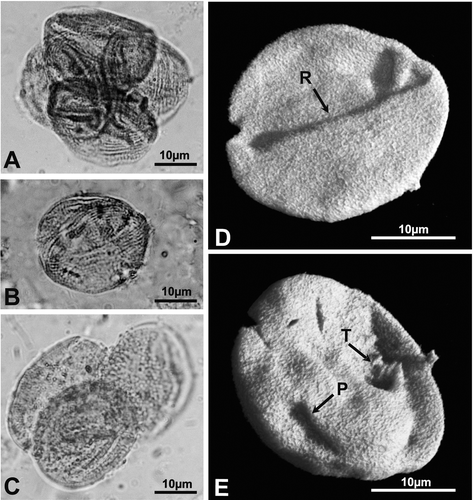
Figure 5 A–G. Type C: Circumpolles gen. et sp. indet. (R – rimula; T – trilete scar; ET – thickened annulus). (A) LM photomicrograph of an individual pollen grain attributed to Circumpolles gen. et sp. indet.; (B) CSLM maximum vertical projection of sections of the same grain; (C) 3‐D rendering based on CSLM sections of the same grain showing the proximal side; (D) LM photomicrograph of another specimen of Circumpolles gen. et sp. indet.; (E) CSLM maximum vertical projection of sections of the same grain; (F) 3‐D rendering based on CSLM sections of the same grain viewed from its proximal side. The triangular area represents the trilete scar; (G) 3‐D rendering based on CSLM sections of the same grain viewed from its distal side. The pore has a thickened annulus.
Discussion
Methodology
Our studies of Classopollis show that CSLM is a powerful tool for morphological analyses of fossil pollen grains. The promptness of analysis compared to SEM and TEM, the possibility of making direct observations on strew slides, and the improved contrast and resolution (especially with respect to depth) compared to that obtained with light microscopy, make it an effective technique filling the gap between traditional wide field light microscopy and electron microscopy. The technique allows 3D reconstructions of the pollen grain without further sample preparation than that required for light microscopy. Together, light microscopy and CSLM are complementary, the former allowing overview observations, the latter providing more structural details.
The exclusive use of electron microscopy for taxonomic purposes has favoured the proliferation of discriminating features, sometime invisible when using light microscopy. This has lead to an increase in the number of published species that may not be possible to recognise in traditional palynological analyses. For example, Reyre Citation(1970) and Reyre et al. Citation(1970) described 15 new species of Classopollis on the basis of micromorphological ornamentation data provided by an impressive volume of SEM evidence. Despite the accuracy of the thorough ultrastructural analyses made, many of these species remain very difficult, or even impossible, to differentiate by light microscopy (Taugourdeau‐Lantz, Citation1971; Boltenhagen, Citation1973).
At low magnifications (×1000 or ×3000) CSLM images are almost as informative as SEM images, but CSLM is clearly less informative at higher magnifications. Similarly, the comparison of optical sections obtained by CSLM (Figures & ) with ultrathin sections obtained by classical TEM techniques (Pettitt & Chaloner, Citation1964; Taylor & Alvin, Citation1984) shows a large difference in resolution in favour of the latter. Thus, the morphological features observed by electron microscopy are commonly invisible in conventional wide field light microscopy and even CSLM. Combinations of light microscopy, SEM and TEM should therefore be used (Leffingwell et al., Citation1970; Doyle et al., Citation1975; Schrank & Mahmoud, Citation2002).
Despite the higher resolutions achieved by electron microscopy techniques, their use in taxonomic studies remains very limited (Zippi, Citation1991), largely because electron microscopy requires time‐consuming sample preparation. Further, it cannot be used to examine problematic forms permanently mounted on strew slides. Certainly, new studies of old spore type collections, as advocated by Jansonius Citation(1981), would be easier to undertake using CSLM and the initiative of Feist‐Burkhardt, Henderson and McLachlan performed on a dinoflagellate cysts collection from the Natural History Museum of London (http://www.nhm.ac.uk/research‐curation/projects/duxbury/) should therefore be encouraged.
Taxonomic aspects of the present results
The morphological analysis of the Type C specimens of Fuentelsaz suggests they may be related to “gemmate” Classopollis‐type pollen grains identified by Médus & Reyre Citation(1966) and Médus Citation(1969, Citation1970) from the Lower‐Middle Jurassic of Southern Europe and North Africa. Médus Citation(1969) noted that the Jurassic “gemmate” forms resemble the Upper Triassic Granuloperculatipollis rudis Venkatachala and Góczán, Citation1964 but differ in having striae. Later, he (Médus, Citation1970) instituted the monotypic genus Gemmulina and described Gemmulina jurassica but omitted to specify diagnosis nor provide illustrations. Furthermore, the absence of detailed ultrastructural studies of these previously described Jurassic forms prevents detailed comparisons.
In a similar way, a conclusive assimilation of the type C specimens of Fuentelsaz to the Upper Triassic species Granuloperculatipollis rudis remains premature. Further, this species is a recognized biomarker in numerous Norian‐Rhaetian sections from India (Bharadwaj, Citation1969), Eastern North America (Fowell & Olsen, Citation1993; Fowell et al., Citation1994; Hounslow et al., Citation2004), Northern Europe (Orbell, Citation1973; Warrington, Citation1974; Warrington & Harland, Citation1975; Schuurman, Citation1977) and Central Europe (Venkatachala & Góczán, Citation1964; Venkatachala, Citation1966; Schulz, Citation1967; Morbey, Citation1975; Schuurman, Citation1979) reputed to not range up the limit Triassic/Jurassic.
For these reasons, it is not possible to assign the Type C specimens to a particular genus and the open nomenclature Circumpolles gen. et sp. indet. has to be applied. A further comparative study involving the analysis of ultrastructural features of Late Triassic and Early Jurassic materials are needed.
Conclusions
The combination of CSLM and conventional wide field light microscopy was found to be adequate for the analysis of morphological types of Classopollis pollen from the Early Jurassic of Spain. On the basis of these morphological studies, the species C. classoides and C. torosus were identified. A third morphotype characterized by the presence of an operculum‐like structure and numerous Ubisch bodies adhered to its surface was described and attributed to Circumpolles gen. et sp. indet.
CSLM allowed the rapid, non‐destructive, 3D characterization of the Classopollis pollen grains, permitting the distinction of internal wall structures and ill‐defined morphological features that cannot be discerned by light microscopy alone. Although CSLM is insufficiently precise for ultrastructural studies at high magnification, it is a helpful tool in taxonomic studies.
Acknowledgements
This work was financed by the projects CGL 2005‐01765/BTE and CGL 2005‐04574/BTE of the Ministerio de Educación y Ciencia (Spain), and by the Basler Stiftung für biologische Forschung (Switzerland). We would like to thank Drs. A. Sahuquet and S. Baghdiguian (Laboratoire de Dynamique Moléculaire des Interactions Membranaires, Université Montpellier II, France), N. Lautredou‐Audouy (Centre Régional d'Imagerie Cellulaire/IURC, Montpellier, France) and Drs. Luis Miguel Alonso Colmenar and Alfonso Cortés Peña (Centro de Microscopía y Citometría, Universidad Complutense de Madrid, Spain) for their help in the confocal microscopy studies. The authors are grateful to Drs. D. Batten and R. Zetter for their helpful comments and corrections to an earlier version of this article. This paper is dedicated to the memory of Dr. Dominique Jolly (Université de Montpellier II), an international leader in palynology, and our colleague and friend.
References
- Balme , B. E. 1957 . Spores and pollen grains from the Mesozoic of Western Australia , Sydney : Commonwealth Scientific and Industrial Research Organization (Australia) . Coal Res. Sect. CSIRO. Tech. Commun. 25
- Banks , H. , Feist‐Burkhardt , S. and Klitgaard , B. 2006 . The unique pollen morphology of Duparquetia (Leguminosae: Caesalpinioideae): Developmental evidence of aperture orientation using confocal microscopy. . Ann. Bot. , 98 : 107 – 115 .
- Barrón , E. and Azêredo , A. C. 2003 . Palynology of the Jurassic (Callovian‐Oxfordian) succession from Pedrogão (Lusitanian Basin, Portugal). Palaeoecological and palaeobiogeographical aspects. . N. Jahrb. Geol. Paläontol. Abh. , 227 : 259 – 286 .
- Barrón , E. , Comas‐Rengifo , M. J. and Trincão , P. 1999 . Estudio palinológico del tránsito Pliensbachiense/Toarciense en la Rambla del Salto (Sierra Palomera, Teruel, España). . Cuad. Geol. Ibérica , 25 : 171 – 187 .
- Barrón , E. , Gómez , J. J. , Goy , A. and Pieren , A. P. 2006 . The Triassic‐Jurassic boundary in Asturias (Northern Spain): Palynological characterization and facies. . Rev. Palaeobot. Palynol. , 138 : 187 – 208 .
- Bharadwaj , D. C. 1969 . Palynological succession through the Mesozoic era in India. . J. Palynol. (India) , 2 : 85 – 94 .
- Birkmann , H. and Lundin , R. F. 1996 . Confocal microscopy; potential applications in micropaleontology. . J. Palaeontol. , 70 : 1084 – 1087 .
- Boltenhagen , E. 1968 . Révision du Genre Classopollis Pflug. . Rev. Micropaléontol. , 11 : 29 – 44 .
- Boltenhagen , E. 1973 . Quelques espèces du genre Classopollis (Pflug) Reyre du Crétacé Supérieur du Gabon. . Rev. Micropaléontol. , 16 : 205 – 213 .
- Burger , D. 1965 . Some new species of Classopollis from the Jurassic of Netherlands. . Leidse Geol. Mededel. , 33 : 63 – 69 .
- Burger , D. 1966 . Palynology of uppermost Jurassic and lowermost Cretaceous strata in the eastern Netherlands. . Leidse Geol. Mededel. , 35 : 209 – 276 .
- Cornet , B. and Traverse , A. 1975 . Palynological contributions to the chronology and stratigraphy of the Hartford Basin in Connecticut and Massachusetts. . Geosci. Man , 11 : 1 – 33 .
- Couper , R. A. 1958 . British Mesozoic microspores and pollen grains. . Palaeontographica B , 103 : 75 – 179 .
- Cresta , S. , Goy , A. , Ureta , S. , Arias , C. , Barrón , E. , Bernad , J. , Canales , M. L. , García Joral , F. , Garcia Romero , E. , Gialanella , P. R. , Gómez , J. J. , Gonzalez , A. , Herrero , C. , Martinez , G. , Osete , M. L. , Perilli , N. and Villalain , J. J. 2001 . The global boundary stratotype section and point (GSSP) of the Toarcian‐Aalenian boundary (Lower‐Middle Jurassic). . Episodes , 24 : 166 – 175 .
- Doyle , J. A. , Van Campo , M. and Lugardon , B. 1975 . Observations on exine structure of Eucommiidites and Lower Cretaceous angiosperm pollen. . Pollen Spores , 17 : 429 – 486 .
- Feist‐Burkhardt , S. and Pross , J. 1999 . Morphological analysis and description of Middle Jurassic dinoflagellate cyst marker species using confocal laser scanning microscopy, digital optical microscopy, and conventional light microscopy. . Bull. Centr. Rech. Elf Explor.‐Prod. , 22 : 103 – 145 .
- Fowell , S. J. and Olsen , P. E. 1993 . Time calibration of Triassic/Jurassic microfloral turnover, eastern North America. . Tectonophysics , 222 : 361 – 369 .
- Fowell , S. J. , Cornet , B. and Olsen , P. E. 1994 . Geologically rapid Late Triassic extinctions: Palynological evidence from the Newark Supergroup. . Geol. Soc. Am. Sp. Pap. , 288 : 197 – 206 .
- Gijzel , P. V. 1967 . Autofluorescence of fossil pollen and spores with special reference to age determination and coalification. . Leidse Geol. Medd. , 40 : 263 – 317 .
- Gomez , B. , Martín‐Closas , C. , Méon , H. , Thévenard , F. and Barale , G. 2001 . Plant taphonomy and paleoecology in the lacustrine Uña delta (Late Barremian, Iberian Ranges, Spain). . Palaeogeogr. Palaeoclimatol. Palaeoecol. , 170 : 133 – 148 .
- Gómez , J. J. and Goy , A. 1979 . Las unidades litoestratigráficas del Jurásico medio y superior en facies carbonatadas del Sector Levantino de la Cordillera Ibérica. . Est. Geol. , 35 : 17 – 57 .
- Gómez , J. J. and Goy , A. 2005 . Late Triassic and Early Jurassic palaeogeographic evolution and depositional cycles of the Western Tethys Iberian platform system (Eastern Spain). . Palaeogeogr. Palaeoclimatol. Palaeoecol. , 222 : 77 – 94 .
- Goy , A. , Gómez , J. J. and Yébenes , A. 1976 . El Jurásico de la Rama Castellana de la Cordillera Ibérica (Mitad Norte). Unidades litoestratigraficas . Est. Geol. , 32 : 391 – 423 .
- Goy , A. , Ureta , S. , Arias , C. F. , Canales , L. M. , García Joral , F. , Herrero , C. , Martínez , G. and Perilli , N. 1994 . The Fuentelsaz section (Iberian range, Spain), a possible stratotype for the base of the Aalenien Stage. . Serv. Geol., Nazl. Miscellanea , 5 : 1 – 3 .
- Goy , A. , Ureta , S. , Arias , C. F. , Canales , M. L. , García Joral , F. , Herrero , C. , Martínez , G. and Perilli , N. 1996 . Die Toarcium‐Aalenium‐Grenze im profile Fuentelsaz (Iberische Ketten, Spanien). . Geol. Landesamt Baden‐Württemberg , 8 : 43 – 52 .
- Hochuli , P. and Feist‐Burkhardt , S. 2004 . A boreal early cradle of Angiosperms? Angiosperm‐like pollen from the Middle Triassic of the Barents Sea (Norway). . J. Micropalaeontol. , 23 : 97 – 104 .
- Hounslow , M. W. , Posen , P. E. and Warrington , G. 2004 . Magnetostratigraphy and biostratigraphy of the Upper Triassic and lowermost Jurassic succesion, St. Audrie's Bay, UK. . Palaeogeogr. Palaeoclimatol. Palaeoecol. , 213 : 331 – 358 .
- Jansonius , J. 1981 . Linnean nomenclature Universal language of taxonomists and the sporae dispersae (with a commentary on Hughes' proposal). . Taxon , 30 : 438 – 448 .
- Leffingwell , H. A. , Larson , D. A. and Valencia , M. J. 1970 . A study of the fossil pollen Wodehouseia spinata. I. Ultrastructure and comparisons to selected modern taxa. II. Optical microscopic recognition of foot layers in differentially stained fossil pollen and their significance. . Bull. Can. Petrol. Geol. , 18 : 238 – 262 .
- Malyavkina , V. S. 1949 . Identification of spores and pollen of the Jurassic and Cretaceous , Leningrad (St. Petersburg)/Moskva : VNIGRI. N. Ser. Contrib. 33 .
- Médus , J. 1969 . Étude morphologique des grains de pollen appartenant au groupe des circumpolles (Pflug) Klaus dans le Jurassique de l'Aquitaine occidentale. . Geobios , 2 : 81 – 88 .
- Médus , J. 1970 . Contribution à la classification des grains de pollen du groupe des circumpolles (Pflug) Klaus. . Pollen Spores , 12 : 205 – 216 .
- Médus , J. and Reyre , Y. 1966 . Contribution à l'étude des grains de pollen appartenant au genre de forme Classopollis (Pflug) Pocock et Jansonius. . C. R. Acad. Sci. (Paris) Ser. D , 262 : 2703 – 2706 .
- Morbey , S. J. 1975 . The palynostratigraphy of the Rhaetian stage, upper Triassic in the Kendelbachgraben, Austria. . Palaeontographica B , 152 : 1 – 75 .
- O'Connor , B. 1996 . Confocal Laser Scanning Microscopy: A new technique for investigating and illustrating fossil Radiolaria. . Micropaleontology , 42 : 395 – 402 .
- Orbell , G. 1973 . Palynology of the British Rhaeto‐Liassic. . Bull. Geol. Surv. G. B. , 44 : 1 – 44 .
- Pettitt , J.‐M. and Chaloner , W. G. 1964 . The ultrastructure of the Mesozoic pollen grain Classopollis. . Pollen Spores , 6 : 611 – 620 .
- Pflug , H. 1953 . Zur Entstehung und Entwicklung des Angiospermiden pollens in der Erdgeschichte. . Palaeontographica B , 95 : 60 – 171 .
- Pocock , S. J. and Jansonius , J. 1961 . The pollen genus Classopollis Pflug, 1953. . Micropaleontology , 7 : 85 – 153 .
- Pocock , S. J. , Vasanthy , G. and Venkatachala , B. S. 1990 . Pollen of circumpolles – An enigma or morphotrends showing evolutionary adaptation. . Rev. Palaeobot. Palynol. , 65 : 179 – 193 .
- Reissinger , A. 1950 . Die “Pollenanalyse,” ausgedehnt auf alle Sedimentgesteine der geologischen Vergangenheit. . Palaeontographica B , 90 : 99 – 126 .
- Reyre , Y. 1970 . Stereoscan observations of the pollen genus Classopollis Pflug 1953. . Palaeontology , 13 : 303 – 322 .
- Reyre , Y. , Kieser , G. and Pujol , C. 1970 . Interet stratigraphique de quelques espèces du genre Classopollis (Pflug) Reyre. . Rev. Micropaléontol. , 13 : 146 – 154 .
- Salih , A. , Jones , A. S. , Bass , D. and Cox , G. 1997 . Confocal imaging of exine as a tool for grass pollen analysis. . Grana , 36 : 215 – 224 .
- Schrank , E. and Mahmoud , M. S. 2002 . Barremian angiosperm pollen and associated palynomorphs from the Dakhla Oasis area, Egypt. . Palaeontology , 45 : 33 – 56 .
- Schulz , E. 1967 . Sporenpaläontologische Untersuchungen rëtoliassischer Schichten im Zentralteil des Germanischen Beckens. . Paläontol. Abh. B , 2 (3) : 427 – 633 .
- Schuurman , W. M. L. 1977 . Aspects of Late Triassic Palynology. 2. Palynology of the “Grès et schiste à Avicula contorta” and “argiles de Levallois” (Rhaetian) of Northeastern France and Southern Luxemburg. . Rev. Palaeobot. Palynol. , 23 : 159 – 253 .
- Schuurman , W. M. L. 1979 . Aspects of Late Triassic palynology. 3. Palynology of latest Triassic and earliest Jurassic deposits in the Northern Limestone Alps in Austria and Southern Germany, with special reference to a palynological characterisation of the Rhaetian stage in Europe. . Rev. Palaeobot. Palynol. , 27 : 53 – 75 .
- Scott , A. C. and Hemsley , A. R. 1991 . Comparison of new microscopical techniques in the study of fossil spore wall ultrastructure. . Rev. Palaeobot. Palynol. , 67 : 133 – 139 .
- Shute , C. , Hemsley , A. R. and Strother , P. 1996 . Reassessment of dyads contained in a Late Silurian Rhynophytoid sporangium. . Sp. Pap. Palaeontol. , 55 : 137 – 145 .
- Skog , J. E. 2005 . Report of the Committee for Fossil Plants. . Taxon , 54 : 827
- Srivastava , S. K. 1976 . The fossil pollen genus Classopollis. . Lethaia , 9 : 437 – 457 .
- Taugourdeau‐Lantz , J. 1971 . A propos du genre Classopollis (Pflug) Pocock & Jansonius, un phénomène nouveau. . Rev. Micropaléontol. , 14 (1) : 50 – 54 .
- Taylor , T. N. 1999 . “ The ultrastructure of fossil pollen and spores. ” . In Fossil plants and spores: Modern techniques , Edited by: Jones , T. P and Rowe , N. P . 126 – 131 . London : Geol. Soc .
- Taylor , T. N. and Alvin , K. L. 1984 . Ultrastructure and development of Mesozoic pollen: Classopollis. . Am. J. Bot. , 71 : 575 – 587 .
- Traverse , A. 2004 . Proposal to conserve the fossil pollen morphogeneric name Classopollis against Corollina and Circulina. . Taxon , 53 : 847 – 848 .
- Venkatachala , B. S. 1966 . Mesozoic operculate pollen and their morphology. . Palaeobotanist , 15 : 98 – 101 .
- Venkatachala , B. S. and Góczán , F. 1964 . The spore‐pollen flora of the hungarian “Kössen facies”. . Acta Geol. Hung. , 8 : 202 – 228 .
- Warrington , G. 1974 . Studies in the palynological biostratigraphy of the British Trias. I. Reference sections in West Lancashire and North Somerset. . Rev. Palaeobot. Palynol. , 17 : 133 – 147 .
- Warrington , G. and Harland , R. 1975 . Palynology of the Trias and Lower Lias of the Larne borehole. . Bull. Geol. Surv. G. B. , 50 : 37 – 49 .
- Wood , G. D. , Gabriel , A. M. and Lawson , J. C. 1996 . “ Palynological techniques – Processing and microscopy. ” . In Palynology: Principles and applications. Vol. 1–3 , Edited by: Jansonius , J and McGregor , D. C . 29 – 50 . College Station, TX : AASP Found .
- Yang , W. P. , Zhang , H. C. and Xu , F. M. 1998 . A tentative study on the ultrastructure of the Middle Permian acritarch Micrhystridium from western Yunnan, China. . Chin. Sci. Bull. , 43 : 1823 – 1826 .
- Zippi , P. A. 1991 . SEM and light microscope mounting and specimen location technique for same‐specimen study of palynological strew mounts. . Micropaleontology , 37 : 407 – 413 .