Abstract
While macrofossil remains of herbs and flowers have disintegrated beyond identification, pollen survives the digestive process and can be identified, sometimes to species level, and is hence an important tool in determining and reconstructing animal diets.
This study compares macroscopic plant parts and pollen remains in faeces from grouse. While macroscopic remains reflect the quantity of bark, twigs, berries and leaves in the birds' food, the pollen also reflects other food, such as catkins, flower buds with developed pollen, flowers and pollen attached to leaves.
Both techniques reflect that Betula and Salix are the most important food for grouse in early Spring. Pollen analysis shows that later in Spring, after snow-melt, the birds are attracted to different plants, firstly, it is taxa within the Ericales and their fruits from the previous year. The nutrient value of food in periods with more or less snow cover is based on the environmental conditions of the previous year. Later on, different herbs, including insect-pollinated plants rich in nectar/sugar, are recorded in the pollen but are not reflected in the macro study, and are therefore lost in most calculations and not discussed. Nutrient content calculations based on Spring and Summer flowers need to be compared with the environmental and climate conditions of the current year.
The grouse has received much scientific attention (e.g. Bergerud & Gratson, Citation1988; Martin et al., Citation1995; Hannon & Dobush, Citation1997). The supply of plant food during Winter, Spring, and early Summer is of special importance for the grouse. Adaptive strategies and macroscopic studies of the food of grouse show that birch (Betula spp.) and willow (Salix spp.) are the main dietary components during the Winter (e.g. Huempfner & Tester, Citation1988; Bergerud & Gratson, Citation1988; Gardarsson, Citation1988). The study by Jenkins et al. (Citation1963) concluded that over 90% of the birds' diets in the Spring was heather shoots – mostly woody cells. In Norway, similar studies by Holmboe (Citation1924), Nordhagen (Citation1928), Lid & Meidell (Citation1933), Myrberget (Citation1979, Citation1981, Citation1988), and Blom (Citation1980) show that the food in Winter consists mainly of Betula catkins, but that available shoots and winter-green parts of different Ericales taxa are also eaten (Myrberget, Citation1979; Blom, Citation1980), again woody elements. The potential importance of pollen and flowers of herbs with nutrient-rich content, like nectar, has hardly been discussed. Does the selective use of only macro-remains exclude important information about bird feeding?
In the present study, pollen analysis is used as an additional method in the study of macroscopic remains on the same material collected on the island of Tran⊘y, Senja, north Norway. The results are discussed in relation to the time of the yield (food) production versus the time of sampling.
Geography
The early Spring situation is suggested to be one of the main reasons why a part of the population of grouse (Lagopus lagopus subsp. lagopus L.) on the island of Senja moves to the nearby and smaller island Tran⊘y in March–April to breed (Myrberget, Citation1972). Different studies at this island have dealt with the flora and vegetation (e.g. Myrberget, Citation1972, Citation1975; Myrberget & Erikstad, Citation1976) and the grouse (Myrberget, Citation1972, Citation1975, Citation1979; Blom Citation1980). The peri-coastal island Tran⊘y (ca. 1.27 km2) (69°09’N, 17°25’E) is situated in the Senja municipality in Troms county (). The vegetation is a mosaic woodland (about 44%) mainly dominated by Betula pubescens Ehrh.; heaths (25%); bogs and fens (8%); shoreline communities (3.5%); and cultivated areas (one farm) (12.5%). A list of plants has been made by Myrberget & Erikstad Citation(1976), and a vegetation map is available for the island (Olav Hesjedal in: Myrberget, Citation1975). Trees, such as Sorbus aucuparia L., Alnus incana (L.) Moench, Salix spp., Populus tremula L., and some Pinus sylvestris L. are present. Stands of Picea abies (L.) H. Karst have been planted. The dwarf shrub Empetrum hermaphroditicum (Lange ex Hagerup) Böcher dominates the ground vegetation together with significant occurrences of Vaccinium, Eriophorum, and Carex species.
Figure 1. Survey map of the studied island of Tran⊘y, Troms county, northern Norway.
A pollen calendar for Tran⊘y has not been made. The nearest one, from the city of Troms⊘, 80 km NE of the area, documents the start of pollen dispersal at the end of May with Alnus (A. incana) and Salix spp. [The earliest records are from 1984 (Ramfjord Citation1991)]. Following shortly after, Betula and Juniperus begin flowering. The first herbs start to flower in the second half of June (Ramfjord, Citation1986: figure 6; Ramfjord, Citation1991: figure 7), and Ericales still later. The vegetation season at Tran⊘y typically starts 2–3 weeks before Troms⊘ (Karl-Dag Vorren, pers. comm.) and the results should be discussed in relation to the expected phenology for the local area.
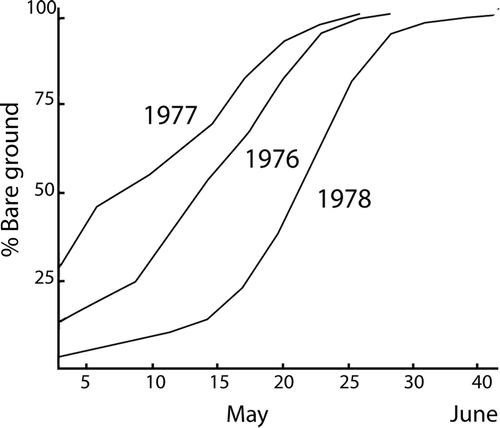
The island is, for this region, known to have an early spring. However, snow covers most of the area until May. Before 10 May in general, the snow covers more than 75% of the area, and after 15 May, less than 50% (). Later on more and more snow-free areas develop (Blom, Citation1980).
Figure 2. Course of snow melt on Tran⊘y, 1976–1978. Mean values for all vegetation types (after Blom, Citation1980).
Material and methods
The present analysis is based on faecal samples from grouse droppings collected by R. Blom on the island of Tran⊘y in 1976. The fieldwork covered the years 1976, 1977, and 1978, with sampling during April–May each year, and analysis of macro remains are presented and discussed in Blom (Citation1980). Sufficient material for pollen analysis was only available for the year 1976, and the 16 samples cover the period from 3 to 31 March 1976, with one to four samples a day.
Pollen analysis is a well known technique, and has also been used on the faeces, stomach contents and coprolite material of different mammals (e.g. Tikhomirov & Kuprianova, Citation1954; King, Citation1977; Moe, Citation1983; Justesen, Citation1992; Bjune, Citation2000). With pollen analysis, pollen and spores can be identified to family and/or genus level, in some cases also to species level.
In this study, the laboratory procedures follow standard methods and available keys for spore and pollen identification (Fægri & Iversen, Citation1989). Two bird droppings were used in each preparation.
Figure 3. Joint diagrams with identified and selected pollen taxa (marked P) and macro remains. Pollen calculations are in % of the total pollen sum; for spores: % of sum pollen + selected taxon; the hollow silhouettes denote a 10 × exaggeration of the percentage values shown in black. Macrofossil remains are presented as histogram bars in number of remains found (after Blom, Citation1980). (Complete diagram of pollen analysis available on request). A. Trees and shrubs; B. Herbs.

Results
The amount of pollen found varied significantly, both during the recorded period, and also between faeces collected on the same day. The rapid changes and peaks are explained as individual behaviour and interest and presence of food (Moe, Citation1983). In the earliest samples in May (sample 1 and 2), the food was dominated by Betula pollen. In samples 3 and 4, the diet has changed to also include pollen of Pinus and Juniperus. In addition, the birds have used herb remnants from the year before containing pollen. Old flowers are possible as food, as well as pollen attached to leaves. It is of interest that the recorded taxa all are insect pollinated. In the samples from 13 May (spectra 5 and 6) the results obviously reflect birds feeding within different types of habitat with more fresh food: in Betula stands and on Empetrum-heathland. Later on, Vaccinium species with flowers and nectar must have been attractive food. A similar situation is seen on 24 and 29 May where different plant taxa have been the selected food, such as different Ericales, Poaceae, Cyperaceae, and Potentilla. Most of the pollen herb taxa reflect known attractive fodder plants for domestic animals, and variation and differences reflect the use of different habitats (e.g. Moe, Citation1983).
In the present study, vegetation types as Betula-woodland, Salix shrub, mire and different kinds of heathland and meadow vegetation are likely to be used for available food. Of special interest are insect-pollinated taxa where the pollen itself is attractive food alone or together with nectar (Fægri & van der Pijl, Citation1966), taxa such as Filipendula cf. ulmaria (L.) Maxim., Potentilla, Asteraceae, Campanula cf. rotundifolia L., Cerastium, Cornus suecica L., Dryas octopetala L., Lotus cf. corniculatus L., and Primula cf. scandinavica.
Based on the pollen content, three main periods can be distinguished: Period 1 (sampled between 3–16 May 1976), characterised by dominant values of Betula pollen together with lower values of Sorbus, Juniperus, some Ericales/Empetrum (), and pollen of different herbs like Cyperaceae, Filipendula and Poaceae (); Period 2 (16–29 May) is characterised by lower values of tree pollen and changing values of Ericales/Vaccinium and herb pollen; and Period 3 (29–31 May) by pollen of Vaccinium, Cyperaceae and herbs, as well as other pollen of Potentilla-type.
Discussion
The macrofossil content of the faeces corresponds with the pollen data in the early period 1 (); with remains of Betula twigs and catkins found together with remains of Salix and different species within the Ericales. The high values of Betula-pollen between 3 and 16 May () reflect a more permanent stay by the birds inside the Betula vegetation, and the attractive food needs to have been mainly fresh or Betula catkins from the previous season, while the fresh herb vegetation on the ground had hardly started to flower because of the local snow cover (Blom, Citation1980). The results clearly show that Betula twigs and catkins had been the major food resource during the first part of the studied period, when there was a minimum of bare ground.
The herb pollen found in Period 1 requires a comment. Pollen of Filipendula, Ranunculaceae, and Rumex acetosa-type, and spores of Polypodiaceae (), can hardly reflect any new spring vegetation, but rather plant debris from the year before (1975). Such pollen may occur both in melting water, mud, and soil. Some taxa within Cyperaceae (e.g. Eriophorum), however, on optimal exposed sites may have started to produce some attractive ‘fresh’ food in that year such as shoots and flowers. On the other hand, the macrofossil study demonstrates the use of leaves, berries, and stems of different Ericales taxa supported by a minimum of pollen of the same species. The pollen which is found therefore may have been one-year-old pollen attached to berries and leaves. Also the berries are from the previous year. The first indication that the grouse have eaten flowers with fresh or new pollen is dated to 13 May (Empetrum). Some days later, pollen of Vaccinium is also recorded, indicating the onset of flowering. By mid-May, the use of Eriophorum flowers may have been of importance.
During Period 2, with more than 50% bare ground, the pollen composition shows an interesting change. The dominant use of pollen-rich Betula catkins has ended (Ramfjord, Citation1986, Citation1991), but still some Betula tissue is found. Salix, which flowers somewhat later than Betula species, is now recorded both in the pollen and macrofossil remains. It should be kept in mind that both the pollen of Salix and also Alnus (A. incana) would have been produced the year before.
Significant amounts of both pollen and macrofossil remains of Ericales are found, and have probably been the main food source. The macrofossil values of Poaceae flowers and most of the herb remains represent flowers from the previous year. The normal flowering time for Poaceae in this area starts one to two months later (Ramfjord, Citation1986, Citation1991). The pollen analysis alone, however, in this period includes several taxa which may have been of special importance and which may have grown to some extent as a part of the early spring flora in that year, such as Potentilla, Primula (P. cf. farinosa), Rosaceae, and, later on, also Rubus chamaemorus. Increased food availability due more species present makes the grouse select the plants with the best taste and highest forage quality.
In Period 3 most of the area is free of snow, and the two methods correspond with high values of Ericales. Concerning the herbs, the macrofossil data are available but mostly unidentified. The pollen diagram informs us that in addition to Vaccinium, different herbs such as Filipendula (F. ulmaria), Potentilla, Rubus chamaemorus, and Lotus (cf. corniculatus) have been attractive as food (Myrberget, Citation1975). The food supply is dominated by fresh Spring/early-Summer attractive food.
The lack of identification of herbs in the macrofossil study during Period 2 and especially during Period 3 reduces the usefulness of such data alone as a basis for quantitative calculations. The pollen data contribute a wider set of bird-menu parameters including nutrient-rich plants, especially from mid-May onwards.
Only during the late spring onwards is the food based on the yield of that actual year. Fresh pollen is rich in nutrients. Insect-pollinated taxa are even more attractive fodder because in most cases they contain energy-rich floral or extra floral nectar (e.g. Fægri & Iversen, Citation1989; Fægri & van der Pijl, Citation1966). Attractive herbs which belong to these groups, and which are recorded, are of special interest for energy and nutrient supply for different kinds of animal (e.g. Moe, Citation1983).
The finding that some of the plants eaten during the winter and in the spring are based on production from the previous Summer/Autumn (Miller & Watson, Citation1978: 296) is of special importance. First of all, trees and shrub taxa with catkins, shoots, berries, and all edible parts have been produced the year before.
While available Winter and early Spring diet are based on environmental and climatic conditions in the year before, the late Spring, Summer and Autumn diet are more a result of the conditions in that year. Later on, the climate and duration of the snow cover, certainly varying from year to year (), are important factors in general (e.g. Lance, Citation1978) determining the transition from Betula to Ericales in the birds' diets.
Conclusion
Both macrofossil and pollen analysis are important methods for studying the diet for grouse and terrestrial animals. The macrofossil studies reflect a certain number of woody taxa and are useful for a volumetric quantification of the food. The pollen analysis, as an easy method, includes in a more qualitative way a larger number of pollen taxa, of which the herb taxa during late spring and the earlier summer season enlarge the picture of the local bird diet. A proper energy supply calculation needs to include nutrient-rich herbs as well as available woody taxa. The study of pollen and herbs with intra- and extra-floral nectars during spring and early summer is of special interest and needs to be included to give a more proper energy and nutrient calculation. Pollen analysis could therefore be of special interest and contribute important elements to the discussion about selection and changes of diet. Pollen analysis alone is, however, not a good basis for the quantification of food. The methods are therefore supplementary.
Use of macro and micro analysis of bird droppings and obviously, if possible, direct observation techniques on feeding birds, will give a better understanding of the selection of plant food and energy supply. In this study lack of sufficient field data prevent us presenting proper energy and nutrient calculations.
Acknowledgements
We would like to thank Yngvar Hagen and Odd Jacobsen for allowing us to use unpublished data and material, Eldar Gaare, NINA for help with literature, Karl-Dag Vorren, Ingvar Byrkjedal, Vigdis Vandvik for valuable data, discussion and comments to the manuscript, and James Greig and John Birks for improving the language. Special thanks are due to Prof. Sigmar Bortenschlager of the Institut für Botanik der Universität Innsbruck for making working facilities available for DM. We are grateful to the two anonymous reviewers and Richard Bradshaw for their helpful comments. This is publication No. A227 from the Bjerknes Centre for Climate Research.
References
- Bergerud , A. T. and Gratson , M. W. , eds. 1998 . Adaptive strategies and population ecology of northern grouse , Minneapolis, MN : Univ. Minn. Press .
- Bjune , A. E. 2000 . Pollen analysis of faeces as a method of demonstrating seasonal variations in the diet of Svalbard reindeer (Rangifer tarandus platyrhynchus) . Polar Res. , 19 : 183 – 192 .
- Blom , R. 1980 . Spring foods of the grouse, Lagopus L. lagopus on Tran⊘y, northern Norway , Bergen : Univ. Bergen. MSc Thes .
- Fægri , K. and Iversen , J. 1989 . Textbook of pollen analysis , 4th , Edited by: Fægri , K. , Kaland , P. E. and Krzywinski , K. Chichester/New York/Brisbane/Toronto/Singapore : J. Wiley & Sons .
- Fægri , K. and van der Pijl , L. 1966 . The principles of pollination ecology , Oxford : Pergamon Press .
- Gardarsson , A. 1988 . “ Cyclic population changes and some related events in rock ptarmigan in Iceland ” . In Adaptive strategies and population ecology of northern grouse , Edited by: Bergerud , A. T. and Gratson , M. W. 300 – 329 . Minneapolis, MN : Univ. Minn. Press .
- Hannon , S. J. and Dobush , G. 1997 . Pairing status of male ptarmigan: Is polygony costly to males? . Anim. Behav. , 53 : 369 – 380 .
- Holmboe , J. 1924 . Hvad liryperne lever av i Norge , Bergen : Bergens Mus. Årbok 1922–23, Naturvitensk. Rekke 5 .
- Huempfner , R. A. and Tester , J. R. 1988 . “ Winter arboreal feeding behaviour of ruffed grouse in East-Central Minnesota ” . In Adaptive strategies and population ecology of northern grouse , Edited by: Bergerud , A. T. and Gratson , M. W. 122 – 157 . Minneapolis, MN : Univ. Minn. Press .
- Jenkins , D. , Watson , A. and Miller , G. R. 1963 . “ Population studies on red grouse, Lagopus lagopus scoticus (Lath.) in north-east Scotland ” . In J. Anim. Ecol Vol. 36 , 97 – 122 .
- Juggins , S. 2003 . C2. A program for analysing and visualising palaeoenvironmental data. Vers. 1.3 , Newcastle, , UK : Sch. Geogr. Pol. Sociol. Newcastle Univ .
- Justesen , J. 1992 . Pollenanalyser av fekalier som metode i vegetasjons – og diettunders⊘kelser , Bergen : Univ. Bergen. MSc Thes .
- King , F. B. 1977 . An evaluation of the pollen contents of coprolites as environmental indicators . J. Ariz. Acad. Sci. , 12 : 47 – 52 .
- Lance , A. N. 1978 . Territories and the food plant of individual red grouse: II. Territory size compared with an index of nutrient supply in heather . J. Ecol. , 47 : 307 – 313 .
- Lid , J. and Meidell , O. 1933 . The food of Norwegian grouse chicks (Lagopus lagopus L.) . N. Mag. Naturvidensk. B , 73 : 75 – 114 .
- Martin , K. , Horn , A. G. and Hannon , S. J. 1995 . The calls and associated behaviour of breeding ptarmigan in Canada . Wilson Bull. , 107 : 496 – 509 .
- Miller , G. R. and Watson , A. 1978 . Territories and the food plant of individual Red Grouse: I. Territory size, number of mates and brood size compared with the abundance, production and diversity of heather . J. Ecol. , 47 : 293 – 305 .
- Moe , D. 1983 . Palynology of sheep's faeces: Relationship between pollen content, diet and local pollen rain . Grana , 22 : 105 – 113 .
- Myrberget , S. 1972 . “ Fluctuations in a North Norwegian population of grouse ” . In Int. Ornithol. Congr., The Hague 1970, Proc , 15th , Edited by: Voous , K. H. 107 – 120 . Leiden : E. J. Brill .
- Myrberget , S. 1975 . Vegetasjonskartlegging av Tran⊘ya i Senja , Trondheim : Dir. Vilt & Ferskvanfsk. Tran⊘y Feltst. Report .
- Myrberget , S. 1979 . Winter food of grouse in two Norwegian areas . Medd. Nor. Viltforsk. , 3 : 1 – 32 .
- Myrberget , S. 1981 . Diet of grouse Lagopus lagopus chicks on a coastal island . Fauna Norv., S.C. Cinclus , 4 : 58 – 63 . Trondheim: NINA
- Myrberget , S. 1988 . “ Demography of an island population of ptarmigan in Northern Norway ” . In Adaptive strategies and population ecology of Northern Grouse , Edited by: Bergerud , A. T. and Gratson , M. W. 379 – 419 . Minneapolis, MN : Univ. Minn. Press .
- Myrberget , S. and Erikstad , K. E. 1976 . Liste over karplanter funnet på Tran⊘ya i Senja , Trondheim : Dir. Vilt & Ferskvanfsk. Tran⊘y Feltst. Report .
- Nordhagen , R. 1928 . Rypeaar og Bæraar , Bergen : Bergens Mus. Årbok, Naturvitensk. Rekke, 2 .
- Ramfjord , H. 1985 . Registrering av pollen og sporer i 1984 , Trondheim : Dept. Bot. NTNU. Rep .
- Ramfjord , H. 1986 . Registrering av pollen og sporer i 1985 , Trondheim : Dept. Bot. NTNU. Rep .
- Ramfjord , H. 1991 . Registrering av pollen og sporer i 1990 , Trondheim : Dept. Bot. NTNU. Rep .
- Tikhomirov , B. A. and Kuprianova , L. A. 1954 . Investigation of pollen from plants remains found in the stomach of the Beresovka-mammoth. St. Petersburg: Imp. Acad. Sci. Mem. Ser. , 7 ( 42 ) (in Russian)