
Abstract
The modern agricultural landscape is characterised by large areas of monoculture, including fields of mass-flowering crops. These may benefit bees by providing abundant forage, but may cause harm if the fields are treated with insecticides. Here, we investigated the potential exposure of honeybee colonies to pesticides in flowering oilseed rape (OSR) in England using a combination of aerial photographs and geographical modelling. Aerial photographs were used to detect and map fields of blooming OSR in eight areas across England where OSR is grown. We determined the distance to the nearest OSR field for randomly chosen positions (i.e., potential hive locations) using a digital map made of each area. The proportion of colonies within foraging range to the nearest OSR field in each area was then calculated using published data on honeybee foraging distances obtained by waggle dance decoding. We found that the distribution of distances to the nearest OSR field varied greatly among study areas, ranging from almost all colonies being within range to almost all being out of range (6–79% were within median foraging distance). In one study area, we examined changes over a 4-year period and found little year-to-year variation due to crop rotation (23–36% within median foraging distance). Our specific results show that exposure in England to OSR and to any chemicals used in treatment varies greatly among areas. Overall, our methodology provides a novel way of quantifying the exposure of bee colonies or other central place foragers to a particular crop or land-type at a landscape level.
El paisaje agrícola moderno se caracteriza por extensas áreas de monocultivo, incluyendo campos de cultivos de floración masiva. Éstos pueden beneficiar a las abejas al proporcionar alimento abundante, pero pueden causar daño si los campos son tratados con insecticidas. Aquí se investigó la posible exposición de las colonias de abejas de miel a la floración de la colza (OSR por sus siglas en inglés) en Inglaterra mediante el uso de una combinación de fotografías aéreas y de modelado geográfico. Se usaron fotografías aéreas para detectar y mapear campos de OSR en ocho áreas de 20 x 20 km a lo largo de Inglaterra donde se cultiva la colza. Se determinó la distancia al campo de OSR más cercano desde posiciones escogidas aleatoriamente (es decir, localizaciones potenciales de colmenas) utilizado un mapa digital hecho para cada área. La proporción de colonias dentro del rango de pecoreo del campo de colza más cercano se calculó entonces a partir de los datos publicados sobre las distancias de pecoreo de las abejas melíferas, obtenidas mediante la decodificación de la danza de las abejas. Encontramos que la distribución de distancias al campo de colza más cercano varió mucho entre las áreas de estudio, oscilando desde casi todas las colonias dentro del rango, a casi todas fuera del rango (6% -79% se encontraban dentro de la distancia media de pecoreo). En una de las áreas de estudio, examinamos los cambios en un período de 4 años, encontrando poca variación de año en año debido a la rotación de cultivos (23% - 36% dentro de la distancia media de pecoreo). Nuestros resultados específicos muestran que la exposición en Inglaterra a la colza y a los productos químicos utilizados en su tratamiento, varía mucho entre zonas. En general, nuestra metodología proporciona una nueva forma de cuantificar, al nivel de paisaje, la exposición de las colonias de abejas u otros pecoreadores clave, a un cultivo o uso de suelo particular.
Introduction
The agricultural landscape has seen dramatic modifications in recent decades. (Graf, Müller, Korner, Jenny, & Jenni, Citation2014; Klein Goldewijk & Battjes, Citation1997; Robinson & Sutherland, Citation2002). Major changes include the conversion of natural ecosystems to agricultural land and the intensification of existing farm land (Lambin et al., Citation2001). More subtle changes include the phasing in and out of different crops. In England, the area planted as oilseed rape (OSR) increased from c. 218,000 to 742,000 hectares from 1983 to 2012, while cereals have declined by 727,000 hectares, largely due to reductions in barley (DEFRA, Citation2014). In the UK, 3% of the whole country, 4% of all agricultural land and 12% of arable land was planted to OSR in 2012. OSR is also grown widely in other countries, with the UK yield being only 4% of the world total (DEFRA, Citation2014; FAO, Citation2012).
Changes in crop type can affect wildlife. For example, bird diversity and abundance decrease when the vegetation structure of agricultural land is simplified (Breitbach, Laube, Steffan-Dewenter, & Böhning-Gaese, Citation2010), as would occur when orchards are replaced by grassland or crops. Flower-visiting insects may be harmed by the loss of hay meadows, most of which have been eliminated in the UK since the Second World War (Fuller, Citation1987), and the reduced use of clovers and legumes in pastures (Robinson & Sutherland, Citation2002).
The replacement of cereals by mass-flowering crops such as OSR may potentially confer advantages. Previous studies have shown that mass-flowering crops can enhance the abundance of pollinators because they provide additional pollen and nectar resources for pollinators (Holzschuh, Dormann, Tscharntke, & Steffan-Dewenter, Citation2013; Riedinger, Renner, Rundlöf, Steffan-Dewenter, & Holzschuh, Citation2014; Westphal, Teffan-Dewenter, & Tscharntke, Citation2003). But flowering crops may also harm pollinators if they are treated or contaminated with pesticides, especially insecticides, when they are in bloom (Sandrock et al., Citation2014). Both effects, positive and negative, are likely to be especially important to bees: pollen and nectar from flowers are the main food for both adults and larvae (Michener, Citation2000). Most bees build nests in which brood is reared on pollen and nectar brought to the nest by adult bees, either workers in social species or nest foundresses in solitary and primitively eusocial species (Michener, Citation2000). Larger bees may be particularly able to profit from mass-flowering crops, as they can forage at greater distances (Greenleaf, Williams, Winfree, & Kremen, Citation2007). In addition, social bees like the honeybee may also be able to recruit nestmates. Honeybees use the waggle dance to direct nestmates to rewarding food sources (Couvillon, Citation2012; von Frisch, Citation1946). Other bee species may use the flower scents and signals to alert nestmates to the presence of food, and some also use pheromones to guide nestmates to specific locations (Dornhaus & Chittka, Citation1999; Dornhaus & Chittka, Citation2001). Recruitment may allow the social bees to exploit resources at larger distances (Beekman & Ratnieks, Citation2000; Greenleaf et al., Citation2007), and recruitment allows these central place foragers to exploit the resources efficiently (Biesmeijer & Ermers, Citation1999; Seeley, Camazine, & Sneyd, Citation1991). However, greater efficiency could potentially increase their exposure to insecticides. In addition, pesticides may accumulate in the nest and in stores and lead to an exposure of developing larvae in turn (Sandrock et al., Citation2014).
Because bees are more likely to visit nearby than distant flowers, their exposure to flowering crops will depend on the distance from the nest to the field (Couvillon, Schürch, & Ratnieks, Citation2014a). However, few studies have attempted to quantify the exposure of bee colonies or other central place foragers to a particular crop or land-type at a landscape level. More specifically, we hypothesised that honeybees, who forage at greater distances than most other flower-visiting insects (Ratnieks and Shackleton, Citation2015), would vary in their access to a specific crop – OSR – across England on a per area basis. Such data would be useful to the establishment and maintenance of honeybee colonies.
In this study, we used aerial photographs to map flowering fields of OSR in eight areas, one in each of the eight agricultural regions of England. In one of these areas, we had aerial photographs for 4 consecutive years, 2011–14. We then used computer simulations and previously published data on honeybee foraging range to estimate the proportion of random locations within a landscape that were within foraging range of the nearest OSR field. Overall, our analyses show that there is great variation in potential “exposure” to OSR. In some areas, most locations within the landscape were within foraging range, whereas in one particular region, very few locations were within foraging range. There was no important effect of crop rotation on exposure in the area studied for 4 years.
Materials and methods
Mapping fields of flowering OSR in the eight agricultural regions of England
OSR in bloom with its abundant yellow petals is a dramatic sight. As a result, fields of OSR can easily be detected by eye, including from aerial photographs (e.g., Figure (B)) (Garbuzov, Ouvillon, Schürch, & Ratnieks, Citation2015). To map OSR across England, we made a stratified sample with one study area in each of the eight subregions of England used by Department for Environment, Food & Rural Affairs (DEFRA) for agricultural statistics. In choosing the study areas, we initially examined aerial photographs in Google Earth for the presence of OSR fields. If oilseed rape was present, we then mapped the fields of OSR in a subregion of approximately
using the polygon tool in Google Earth. In total, we mapped approximately 3200 km2. Table lists the eight regions and the nearest town to each mapped area. Figure (A) displays the extent of the sampled fields.
Figure 1. (A) Great Britain showing London (black) and the eight English agricultural regions (grey: North East (NE), North West (NW), Yorkshire and the Humber (Y & H), East Midlands (EM), West Midlands (WM), East of England (E), South East (SE), South West (SW)). Numbers in the regions correspond to the region numbers in Table . (B) 2014 aerial photograph of oilseed rape fields near Brighton in the South East study area. (C) Mapped oilseed rape (OSR) fields from the aerial survey that includes B; OSR fields are the grey and black polygons, with the black polygon denoting the fields shown in B; dots are uniform-randomly distributed point locations, which represent simulated honeybee hives, within a 5 km radius (dotted line) from the centre of the SE study area, from each of which distances to the nearest OSR fields were determined (note, for clarity only 200 point locations are shown but 1000 were used in the simulations).
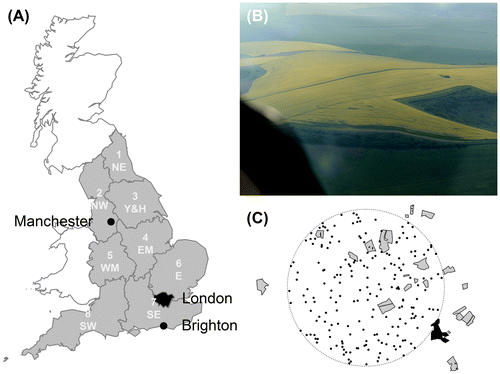
Table 1. The eight agricultural regions of England within each of which we studied one area of c.  (see also Figure (A); the numbers by the region names correspond to the labels in the figure). The region names are those used by DEFRA (Department for Environment, Food & Rural Affairs). The proportion of arable land planted with OSR per region is based on DEFRA statistics for 2010. Also given are the nearest settlements, when the aerial photographs were taken, and the image provider. For four of the eight regions, the imagery date was unavailable.
(see also Figure (A); the numbers by the region names correspond to the labels in the figure). The region names are those used by DEFRA (Department for Environment, Food & Rural Affairs). The proportion of arable land planted with OSR per region is based on DEFRA statistics for 2010. Also given are the nearest settlements, when the aerial photographs were taken, and the image provider. For four of the eight regions, the imagery date was unavailable.
Additionally, for 2011 and 2012 in the South East of England study area, which was in the area around our laboratory, fields planted with OSR were identified from our own previous aerial photography and marked as on the Sussex Habitat Framework, provided to us by the Sussex Wildlife Trust for East Sussex locations (Garbuzov et al., Citation2015). For 2013, we used Google Earth, and for 2014 we made additional aerial photographs ourselves by renting a light aircraft (Garbuzov et al., Citation2015). We exported the OSR fields as shape files and used the maptools package to load them into R. These fields were then mapped out in Google Earth using the polygon tool. All Google Earth-based maps were exported as Keyhole Markup Language (KML) files and imported into R for further processing.
Using Google Earth may introduce a systematic bias in the assessment of where OSR is grown. For our own aerial flight, we could choose the flight days so that they were taking place while OSR was in full bloom. In contrast, when the Google Earth aerial photography was taken was not under our control. We circumvented this problem by choosing areas within each region where OSR was obviously in full bloom, except for the South East area where we had to use the area for which we had aerial photos for previous years, 2011 and 2012. Fortunately, the Google Earth photos for the South East study area were taken during OSR bloom, although judging from the lower intensity of the yellow colour, it was late bloom.
Estimation of distance to nearest OSR field
We imported all shape and KML files into R using the maptools and rgdal package where they were re-projected onto the UK Grid coordinates if necessary. In R, we then created random point locations with uniform distribution, which represent honeybee hives, within a 5-km radius from the centre of the subregion (see Figure (C)). Neither the colonies of honeybees nor bumblebee nests are uniformly randomly distributed (BBKA, Citation2006; Osborne et al., Citation2008), and apiaries are more densely located in some parts of the country (SE & SW) when compared to other parts (NE & NW, Data from National Bee Unit BeeBase). But our study is primarily measuring the exposure of the landscape to mass-flowering crops. Additionally, it also gives a good idea of exposure of central place foragers that are randomly distributed relative to OSR fields. We think this is valid, especially if OSR fields are also considered to be randomly distributed as well. We used rejection sampling to exclude random points that were further than 5 km or within an OSR field. We excluded locations (i.e., hives) within OSR fields because apiaries are not generally set up within arable fields. In addition, bumble bees seldom nest in arable fields (Dave Goulson, pers. comm.). Constraining the locations to a 5 km radius allowed us to avoid edge effects for our sampled points, as the mapped subregion was considerably larger, . With the gDistance function from the rgeos package, we calculated the distance between each randomly generated point and the nearest point of the nearest OSR field. This is a reasonable measure of exposure to OSR. Bees are economically rational foragers, and so the nearest field will be more attractive, as less time and energy are used flying to it (Schmid-Hempel, Citation1987; Schmid-Hempel & Schmid-Hempel, Citation1987, Seeley, Citation1994). In addition, a typical OSR field is many hectares and so is a major source of forage. Previous research involving the decoding of waggle dances has demonstrated that foraging on OSR is predominantly on the nearest OSR field (Garbuzov et al., Citation2015). We used 1000 locations per study area. To check that this was appropriate, we sampled distances from one of our maps multiple times with different sample sizes. Once the number of random locations reached 100, the mean distance estimated stabilised, indicating that 1000 was more than sufficient (data not shown).
Results
Distances to mass-flowering OSR varies greatly among study areas
Median distances to the nearest OSR field for random locations varied greatly among the 8 areas, ranging from 301 m in the East Midlands area to 2903 m in the North West area (Figure (A)). From a variety of methods, it is known that honeybees will forage as far as c. 10–12 km from the hive (Beekman & Ratnieks, Citation2000; Ratnieks, Citation2007). But previous research that decoded honeybee waggle dances has shown that the actual foraging distances of honeybees in spring in the Brighton area are well below this maximum range with a median of 647 m, interquartile range of 364–1305 m, and with 95% of foraging within 2114 m (Couvillon, Schürch, & Ratnieks, Citation2014b; Garbuzov, Schürch, & Ratnieks, Citation2014). Other studies confirm that foraging distances in spring are low (Beekman & Ratnieks, Citation2000). The decreased foraging distance is most likely because forage in the spring is more readily available, so the bees do not have to fly as far to collect forage. The median and 95% quantile are shown on Figure as horizontal dashed and dotted lines, respectively, and show what proportion of the point locations in the study landscape are within honeybee foraging range of the nearest OSR field.
Figure 2. (A) Differences in distances from randomly distributed point locations within the mapped areas, one per English agricultural region, to the nearest OSR field. Boxes show the medians (horizontal black bar) and interquartile ranges (whole box). Whiskers extend to the extreme value or 1.5 times the interquartile range, whichever is lower. The lower horizontal dashed line is the median spring honeybee foraging distance (647 m) and the upper horizontal dashed line is the 95%-quantile (2114 m) from published data on honeybees in the South East study area (Couvillon et al., 2014b). Additionally, the numbers given at the intersection of each box with these lines give the exact percentages of random points within the median and 95%-quantile spring foraging distance, respectively. (B) Median distances of randomly distributed point locations to the nearest OSR field decrease with the proportion of arable land used for OSR in that region (data from DEFRA, 2014). The labels give an abbreviation of the region name corresponding to the x-axis label in A, and the region number as given in Table . The grey curve depicts a back-transformed least-squares fit from a linear model with the logit-transformed proportion planted as OSR as a predictor and distance as response.
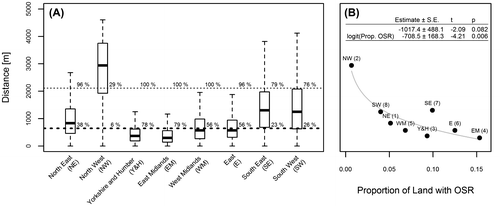
Comparing the median spring foraging distance with the median distance to the nearest OSR fields shows that there is considerable variation in potential exposure to OSR. The most extreme cases are the study areas in the North West and East Midlands (Figure (A)). In the former, only 5.7% of the locations are at a distance to the nearest field that is below median foraging range, and only about 30% are within the 95% quantile. In the latter, almost all locations, 81.4%, are below median foraging distance and all, 100%, are within the 95% quantile. Averaging across all regions, 45.9% of random locations in the landscape are within median honeybee spring foraging distance to the nearest OSR field and 84.5% are within the 95% quantile (Figure (A)).
A linear model confirmed that the differences in distances of these random points co-vary significantly with the logit-transformed proportion of arable land that is used for growing OSR in the region from which our study areas were chosen (Figure (B)).
Distances to OSR vary little across years within a study area
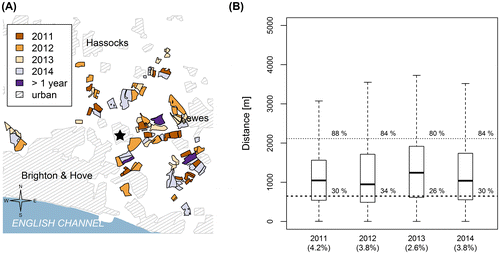
To help control pests and for other agronomic reasons, OSR is usually grown in a crop rotation (Rathke, Christen, & Diepenbrock, Citation2005). For example, in the South East study area, where OSR fields were mapped over four consecutive years, 2011–2014, only 5% of fields were used more than once to grow OSR (Figure (A)). Thus, even if the overall acreage of OSR were unaffected, the changing spatial pattern of the OSR fields could affect the distances from random points to the nearest OSR field. However, the overall effect of this is minor (Figure (B)).
Figure 3. (A) Crop rotation for oilseed rape fields in the South East study area around the city of Brighton and Hove, Sussex, United Kingdom. Shown are the locations of the Laboratory of Apiculture and Social Insects (star), and OSR fields for 2011, 2012, 2013 and 2014. Location data of urban areas were provided by Digimap, a mapping service from EDINA, the national data centre based at the University of Edinburgh. In 2011, 2012 and 2014 OSR fields were mapped on the Sussex Habitat Framework using our own aerial surveys. In 2013, we used aerial photographs available through Google Earth to locate OSR fields and map them onto the Sussex Habitat Framework. (B) Year-to-year differences in distances from randomly distributed points to the nearest mapped OSR fields for the Brighton study area in the South East region (see also Figure (A)). Shown are the medians (black bar) and the interquartile range (box). The whiskers extend to the extreme value or maximally to 1.5 times interquartile range. The wide dashed line represents median spring foraging distance and the thin dashed line is the 95%-quantile of foraging distances. Additionally, the percentages given at the intersection of each box with these lines give the percentages of random points within the median and 95%-quantile Spring foraging distance, respectively. We also give the proportion of land planted with OSR for the 4 years in the x-axis label.
Discussion
Our results show that the simulated exposure of honeybee colonies to OSR varies widely across the 8 study areas. In some areas almost all the colonies would be within foraging range of at least one OSR field, whereas in others only a minority will have easy access to this mass flowering crop during spring foraging. The results also show that the distance to the nearest OSR field co-varies with the proportion of arable land planted with spring blooming OSR (Figure (A)).
Additionally, in the South East study area the results show little variation in the mean random distances to the nearest OSR field even though field locations changed and the area planted to OSR varied from 2.6 to 4.2% of the total area. In all 4 years, the median distance to the nearest OSR field was considerably higher than the median spring foraging distance indicated by waggle dances (Figure (B)).
Our study used aerial photographs obtained from Google Earth for all regions, except for the three years OSR was mapped from our own aerial surveys in the South East study area. The fact that median distances to the nearest field in the South East study area were highest for the year 2013, when we were using Google Earth photos, is consistent with the possibility that we may not have been able to identify all OSR fields (i.e., by missing some that had gone out of bloom) compared to our analysis when we had access to our own aerial survey. However, there was little difference among years (Figure (B)), so that if any bias does exist, it should be very small. In addition, our main conclusion from the multi-year comparison in the South East study area, that there is little year-to-year difference, holds whether or not 2013 is included.
The question of how far mass-flowering OSR is located from hives is not only of academic interest, but also of economic relevance (Kluser, Neumann, Chauzat, & Pettis, Citation2011). For a colony, feeding on OSR during spring could boost food collection and colony development, as it does in other bees (Holzschuh et al., Citation2013; Westphal et al., Citation2003; Westphal, Teffan-Dewenter, & Tscharntke, Citation2009). Improved spring build-up will likely influence parameters such as swarming (Seeley & Visscher, Citation1985), the size of the workforce in summer (Riedinger et al., Citation2014), the amount of honey stored to see the colony through the winter (Seeley & Visscher, Citation1985) and other fitness-relevant measures. The presence or absence of OSR in the vicinity of an insect colony in spring could therefore determine how big the pollinator community is in summer.
On a grimmer note, there is evidence that honeybees and other pollinators suffer lethal and sub-lethal effects after exposure to neonicotinoids under laboratory conditions (Gill, Ramos-Rodriguez, & Raine, Citation2012; Henry et al., Citation2012, Sandrock et al., Citation2014). For this reason, the European Union imposed a 2-year moratorium against the use of neonicotinoid seed dressings on mass-flowering crops starting in December 2013 (EU, Citation2013). However, it remains unclear how much neonicotinoid is actually taken up by flower-visiting insects, which determines the severity of the consequences to insects (Carreck & Ratnieks, Citation2013), and there is also a lack of evidence assessing just how much the populations of honeybees have been affected (Cresswell, Citation2011). As for the beneficial effects of mass-flowering crops, for honeybees and other central place foragers, the amount of neonicotinoids will also depend on their choices of foraging locations, and more distant locations are less visited (Couvillon et al., Citation2014a; Garbuzov et al., Citation2015). Our data show that the exposure of bees to neonicotinoids will almost certainly vary greatly from place to place. Across the whole of England, approximately 50% of the landscape is within honeybee foraging range to the next OSR field based on the median spring foraging distance.
So far we have dealt only with the honeybee, for which good data on both foraging distances in our region and fields of OSR are available (Couvillon et al., Citation2014b; Garbuzov et al., Citation2015). Our findings are also applicable to other bees, as most bees are central place foragers. Bumblebees are a common and important group for which foraging distance estimates are available. However, in contrast to honeybees, these data are not available year round, and are usually collected in summer when the worker populations of bumblebee colonies are greatest (Knight et al., Citation2005; Osborne et al., Citation1999). However, deducing from these data, the potential foraging range of bumblebees seems to be similar to honeybees, so that the potential exposure might be similar. Other bee species with smaller body sizes presumably have smaller foraging ranges, so that the potential exposure of other bees is probably less than that for honeybees (Greenleaf et al., Citation2007). More data on spring foraging distances are required to make conclusive inferences regarding the exposure to OSR of bumble and other bees.
We used random locations to represent “hives”, which gives a picture of the general exposure of the landscape as a whole, and for central place foragers that are randomly distributed in relation to OSR fields. Our methodology could also be used to estimate actual exposure based on known colony or apiary locations. Although it is not compulsory to register hives in England, the National Bee Unit (NBU), which is under the control of DEFRA and the Welsh Government, does maintain records of hive locations in its BeeBase database. In October 2015, we were able to obtain from the NBU the number of apiaries per English county. A quick calculation showed that, not surprisingly, apiary densities vary across England, with the SE and SW possessing substantially more apiaries than the NE and NW. However, as foraging on OSR will depend on the interplay between OSR field locations, apiary locations, and forage availability, which determines how far honeybees are likely to go to find food, and because relative density does not tell us about actual locations, our study using random hive locations must by necessity remain the first step. An obvious future step is to compare the results of the random location method with actual locations. Comparison to actual beehive locations would then show whether our estimates of exposure based on random locations have any consistent bias.
Our estimates of honeybee foraging on OSR is based on springtime foraging, as a majority of OSR in the UK is planted to bloom in April and May, which is a time of year when forage is relatively more abundant, particularly for our study site, but possibly for temperate regions in general (Couvillon et al., Citation2014b). This means that during the spring, when spring OSR is blooming, the honeybees may have alternative sources to choose, such as dandelions (Garbuzov et al., Citation2015). It was also shown that, in the springtime, honeybees may not even visit OSR fields that are located 1200 m or beyond (Garbuzov et al., Citation2015).
However, it is also possible for OSR to be planted to bloom in July. High summer is a time of year that may be characterised by foraging dearth for honeybees and other flower-visiting insects (Couvillon, Nsome, Quah, & Schürch, Citation2014; Couvillon et al., Citation2014b). Therefore, it may be that our estimates of honeybees foraging on OSR would change if the mass-flowering crop was in bloom during the summer: fewer fields of OSR would probably be in bloom (lower % area), but bees would most likely be foraging at a greater distance. Further experimental work is needed to determine honeybee use of OSR during the summer and whether this leads to a lower or greater overall exposure.
In this study, we used the human eye to detect OSR fields from aerial photographs. Attempts have been made to automatically classify land-use data from satellite images (Müller, Böttcher, Meyer-Schatz, & Kage, Citation2008). With its bright yellow fields of flowers and intense bloom, OSR is a good candidate for automated detection. Current challenges in using remote sensing and machine learning to identify crops include the availability of data of sufficient spectral, temporal and spatial resolution and the capacities to recognise field boundaries despite drop shadows and other confounding patterns when using self-organising maps (Kohonen, Citation2001). When satellites with suitable sampling frequency and the right sensors become available (e.g., yellow sensor with 570–590 nm wavelength (Yates & Steven, Citation1987) and weekly sampling given the likelihood of cloud cover, especially in England), our methods in conjunction with remote sensing data may, in the future, be developed to survey wide areas and to extend our understanding of the extent to which bee colonies are exposed to mass-flowering crops such as OSR.
Acknowledgements
We thank the Sussex Wildlife Trust for providing the Sussex Habitat Framework data, Mihail Garbuzov for making available OSR maps for 2011 and 2012, and the National Bee Unit BeeBase for providing information about apiary densities per county.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- BBKA. (2006). Choosing an apiary site. www.bbka.org.uk/files/library/choosing_an_apiary_site_b11_1306865400.pdf
- Beekman, M., & Ratnieks, F. L. W. (2000). Long-range foraging by the honey-bee, Apis mellifera L. Functional Ecology, 14, 490–496.10.1046/j.1365-2435.2000.00443.x
- Biesmeijer, J. C., & Ermers, M. C. W. (1999). Social foraging in stingless bees: How colonies of Melipona fasciata choose among nectar sources. Behavioral Ecology and Sociobiology, 46, 129–140.10.1007/s002650050602
- Breitbach, N., Laube, I., Steffan-Dewenter, I., & Böhning-Gaese, K. (2010). Bird diversity and seed dispersal along a human land-use gradient: High seed removal in structurally simple farmland. Oecologia, 162, 965–976.10.1007/s00442-009-1547-y
- Carreck, N., & Ratnieks, F. L. W. (2013). Will neonicotinoid moratorium save the bees? Research Fortnight, 415, 20–22.
- Couvillon, M. J. (2012). The dance legacy of Karl von Frisch. Insectes Sociaux, 59, 297–306.10.1007/s00040-012-0224-z
- Couvillon, M. J., Fensome, K. A., Quah, S. K. L., & Schürch, R. (2014). Summertime blues: August foraging leaves honey bees empty-handed. Communicative & Integrative Biology, 7, e28821.10.4161/cib.28821
- Couvillon, M. J., Schürch, R., & Ratnieks, F. L. W. (2014a). Dancing bees communicate a foraging preference for rural lands in high-level agri-environment schemes. Current Biology, 24, 1212–1215.10.1016/j.cub.2014.03.072
- Couvillon, M. J., Schürch, R., & Ratnieks, F. L. W. (2014b). Waggle dance distances as integrative indicators of seasonal foraging challenges. PLoS ONE, 9, e93495.10.1371/journal.pone.0093495
- Cresswell, J. (2011). A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees. Ecotoxicology, 20, 149–157.10.1007/s10646-010-0566-0
- DEFRA. (2014). Land use on agricultural holdings on 1 June, 2014. Author.
- Dornhaus, A., & Chittka, L. (1999). Insect behaviour: Evolutionary origins of bee dances. Nature, 401, 38–38.10.1038/43372
- Dornhaus, A., & Chittka, L. (2001). Food alert in bumblebees (Bombus terrestris): Possible mechanisms and evolutionary implications. Behavioral Ecology and Sociobiology, 50, 570–576.10.1007/s002650100395
- EU. (2013). Commission implementing regulation (eu). In Commission, E. (Ed.) no 485/2013.
- FAO. (2012). Food outlook. Rome: Food and Agriculture Organization of the United Nations.
- Fuller, R. M. (1987). The changing extent and conservation interest of lowland grasslands in England and Wales: A review of grassland surveys 1930–1984. Biological Conservation, 40, 281–300.10.1016/0006-3207(87)90121-2
- Garbuzov, M., Couvillon, M. J., Schürch, R., & Ratnieks, F. L. W. (2015). Honey bee dance decoding and pollen-load analysis show limited foraging on spring-flowering oilseed rape, a potential source of neonicotinoid contamination. Agriculture, Ecosystems and Environment, 203, 62–68.10.1016/j.agee.2014.12.009
- Garbuzov, M., Schürch, R., & Ratnieks, F. L. W. (2014). Eating locally: Dance decoding demonstrates that urban honey bees in Brighton, UK, forage mainly in the surrounding urban area. Urban Ecosystems, 18, 411–418.
- Gill, R. J., Ramos-Rodriguez, O., & Raine, N. E. (2012). Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature, 491, 105–108.
- Graf, R., Müller, M., Korner, P., Jenny, M., & Jenni, L. (2014). 20% loss of unimproved farmland in 22 years in the Engadin, Swiss Alps. Agriculture, Ecosystems & Environment, 185, 48–58.
- Greenleaf, S., Williams, N., Winfree, R., & Kremen, C. (2007). Bee foraging ranges and their relationship to body size. Oecologia, 153, 589–596.10.1007/s00442-007-0752-9
- Henry, M., Béguin, M., Requier, F., Rollin, O., Odoux, J.-F., Aupinel, P., … Decourtye, A. (2012). A common pesticide decreases foraging success and survival in honey bees. Science, 336, 348–350.10.1126/science.1215039
- Holzschuh, A., Dormann, C., Tscharntke, T., & Steffan-Dewenter, I. (2013). Mass-flowering crops enhance wild bee abundance. Oecologia, 172, 477–484.10.1007/s00442-012-2515-5
- Klein Goldewijk, C., & Battjes, J. (1997). A hundred year (1890–1990) database for integrated environmental assessments (HYDE, version 1.1). Bilthoven: National institute of public health and the environment (RIVM).
- Kluser, S., Neumann, P., Chauzat, M.-P., & Pettis, J. S. (2011). UNEP emerging issues: Global honey bee colony disorder and other threats to insect pollinators. Geneva: United Nations Environment Programme.
- Knight, M. E., Martin, A. P., Bishop, S., Osborne, J. L., Hale, R. J., Sanderson, R. A., & Goulson, D. (2005). An interspecific comparison of foraging range and nest density of four bumblebee (Bombus) species. Molecular Ecology, 14, 1811–1820.10.1111/mec.2005.14.issue-6
- Kohonen, T. (2001). Self-organizing maps. Berlin: Springer.10.1007/978-3-642-56927-2
- Lambin, E. F., Turner, B. L., Geist, H. J., Agbola, S. B., Angelsen, A., Bruce, J. W., … Xu, J. (2001). The causes of land-use and land-cover change: Moving beyond the myths. Global Environmental Change, 11, 261–269.10.1016/S0959-3780(01)00007-3
- Michener, C. D. (2000). The bees of the world. Baltimore: JHU Press.
- Müller, K., Böttcher, U., Meyer-Schatz, F., & Kage, H. (2008). Analysis of vegetation indices derived from hyperspectral reflection measurements for estimating crop canopy parameters of oilseed rape (Brassica napus L.). Biosystems Engineering, 101, 172–182.10.1016/j.biosystemseng.2008.07.004
- Osborne, J. L., Clark, S. J., Morris, R. J., Williams, I. H., Riley, J. R., Smith, A. D., … Edwards, A. S. (1999). A landscape-scale study of bumble bee foraging range and constancy, using harmonic radar. Journal of Applied Ecology, 36, 519–533.10.1046/j.1365-2664.1999.00428.x
- Osborne, J. L., Martin, A. P., Shortall, C. R., Todd, A. D., Goulson, D., Knight, M. E., … Sanderson, R. A. (2008). Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. Journal of Applied Ecology, 45, 784–792.
- Rathke, G. W., Christen, O., & Diepenbrock, W. (2005). Effects of nitrogen source and rate on productivity and quality of winter oilseed rape (Brassica napus L.) grown in different crop rotations. Field Crops Research, 94, 103–113.10.1016/j.fcr.2004.11.010
- Ratnieks, F. L. W. (2007). How far do honeybees forage? The Beekeepers Quarterly, 89, 26–28.
- Ratnieks, F. L. W., & Shackleton, K. (2015). Does the waggle dance help honey bees to forage at greater distances than expected for their body size? Frontiers in Ecology and Evolution, 3.
- Riedinger, V., Renner, M., Rundlöf, M., Steffan-Dewenter, I., & Holzschuh, A. (2014). Early mass-flowering crops mitigate pollinator dilution in late-flowering crops. Landscape Ecology, 29, 425–435.10.1007/s10980-013-9973-y
- Robinson, R. A., & Sutherland, W. J. (2002). Post-war changes in arable farming and biodiversity in Great Britain. Journal of Applied Ecology, 39, 157–176.10.1046/j.1365-2664.2002.00695.x
- Sandrock, C., Tanadini, L. G., Pettis, J. S., Biesmeijer, J. C., Potts, S. G., & Neumann, P. (2014). Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success. Agricultural and Forest Entomology, 16, 119–128.10.1111/afe.2014.16.issue-2
- Schmid-Hempel, P. (1987). Efficient nectar-collecting by honeybees I. Economic models. The Journal of Animal Ecology, 56, 209–218.10.2307/4810
- Schmid-Hempel, P., & Schmid-Hempel, R. (1987). Efficient nectar-collecting by honeybees II. Response to factors determining nectar availability. The Journal of Animal Ecology, 56, 219–227.10.2307/4811
- Seeley, T. D. (1994). Honey bee foragers as sensory units of their colonies. Behavioral Ecology and Sociobiology, 34, 51–62.10.1007/BF00175458
- Seeley, T. D., Camazine, S., & Sneyd, J. (1991). Collective decision-making in honey bees: How colonies choose among nectar sources. Behavioral Ecology and Sociobiology, 28, 277–290.
- Seeley, T. D., & Visscher, P. K. (1985). Survival of honeybees in cold climates: The critical timing of colony growth and reproduction. Ecological Entomology, 10, 81–88.10.1111/j.1365-2311.1985.tb00537.x
- von Frisch, K. (1946). Die Tänze der Bienen [The Dances of the Bees]. Österreichische Zoologische Zeitschrift, 1, 1–148.
- Westphal, C., Steffan-Dewenter, I., & Tscharntke, T. (2003). Mass flowering crops enhance pollinator densities at a landscape scale. Ecology Letters, 6, 961–965.10.1046/j.1461-0248.2003.00523.x
- Westphal, C., Steffan-Dewenter, I., & Tscharntke, T. (2009). Mass flowering oilseed rape improves early colony growth but not sexual reproduction of bumblebees. Journal of Applied Ecology, 46, 187–193.10.1111/jpe.2009.46.issue-1
- Yates, D. J., & Steven, M. D. (1987). Reflexion and absorption of solar radiation by flowering canopies of oil-seed rape (Brassica napus L.). The Journal of Agricultural Science, 109, 495–502.10.1017/S0021859600081703