
ABSTRACT
The West Palearctic species of Rhoptromeris are revised. A total of 11 species are recognised as valid in this region, including four newly described species: Rhoptromeris dichromata sp. nov., Rhoptromeris koponeni sp. nov., Rhoptromeris leptocornis sp. nov. and Rhoptromeris macaronesiensis sp. nov. Eucoila luteicornis Ionescu, 1959 is synonymised with Rhoptromeris heptoma (Hartig, 1840) syn. nov. A checklist of the Holarctic Rhoptromeris is presented and an identification key to the West Palearctic species is provided. www.zoobank.org/urn:lsid:zoobank.org:pub:8164332C-93E2-4E3F-A408-F5FF5DFB366E
Introduction
Rhoptromeris is a genus of small wasps of the characteristic figitid subfamily Eucoilinae. They are usually less than 2 mm long, and as far as known are koinobiont larval-pupal parasitoids of Chloropidae (Diptera).
The genus belongs to the tribe Trichoplastini Kovalev Citation1989, where it shares with Nanocthulhu Buffington, 2012, Angustacorpa Quinlan, 1988, Stentorceps Quinlan, 1984 and Trichoplasta Benoit, 1956 the lateral bridges on the pronotal plate and the modified second flagellomere of the male’s antenna. Rhoptromeris can be further distinguished by the sloping scutellum, narrow wings and marginal cell with both abscissae of approximately the same size (Nordlander Citation1978). Although readily identifiable by such morphological characters, the monophyly of the genus is not satisfactorily demonstrated, and phylogenetic studies are still required to better elucidate relationships between the genera of Trichoplastini (Buffington et al. Citation2007; Forshage et al. Citation2008).
Rhoptromeris has a cosmopolitan distribution with the exception of the Neotropical region (Forshage and Nordlander Citation2008). The centre of diversity for the genus is in the Afrotropical region, where most of the described species occur (Quinlan Citation1986). The European fauna is second in numbers of described species and was revised by Nordlander (Citation1978), who presented a redescription of the genus, provided information about its biology and dealt with a number of nomenclatural issues. Nordlander and Grijpma (Citation1991) later described one more species, Rhoptromeris strobigena Nordlander and Grijpma, Citation1991, from northern Europe.
Females of most Rhoptromeris search for hosts on grass or low-growing vegetation (Nordlander Citation1978). The exception is R. strobigena, which attacks Hapleginella (Diptera: Chloropidae) and Gaurax spp. (Diptera: Chloropidae) in cones of Larix and Abies (Nordlander and Grijpma, Citation1991). The parasitoid habits of Rhoptromeris heptoma Hartig, 1941 have been extensively studied, as the species attacks Oscinella frit (L.) (Diptera: Chloropidae), a common pest of cereals (Nordlander Citation1978).
In recent years, an important contribution to entomological studies in Sweden has been made by The Swedish Malaise Trap Project (SMTP), a large-scale inventory programme, connected with the Swedish Museum of Natural History and the Swedish Taxonomy Initiative, to increase the knowledge of insect diversity in the country. The collecting efforts sponsored by the project resulted in thousands of samples and millions of insect specimens from several regions of Sweden, including a large number of Rhoptromeris. The current study is one of the many results of this project.
The aim of the present study is to revise the West Palearctic species of Rhoptromeris.
Material and methods
List of depositories
| BMNH | = | The Natural History Museum, London, United Kingdom |
| CAS | = | California Academy of Sciences, San Francisco, USA |
| MGAB | = | Muzeul National de Istorie Naturala ‘Grigore Antipa’, Bucuresti, Romania |
| MNCN | = | Museo Nacional de Ciencias Naturales, Madrid, Spain |
| MZLU | = | Museum of Zoology, Lund University, Lund, Sweden |
| NHRS | = | Naturhistoriska Riksmuseet, Stockholm, Sweden |
| OLML | = | Oberösterreichisches Landesmuseum, Linz, Austria |
| SLU | = | Sveriges Lantbruksuniversitet, Uppsala, Sweden |
| ZIN | = | Zoological Institute – Russian Academy of Sciences, St. Petersburg, Russia |
| ZMUN | = | University of Oslo, Zoological Museum, Oslo, Norway |
Specimens were also obtained on loan from the following private collections:
| Coll. Koponen | = | Coll. Martti Koponen, Otava, Finland |
| Coll. Munk | = | Coll. Thorkild Munk, Aarhus, Denmark (Now in the Zoological Museum, University of Copenhagen, Copenhagen, Denmark) |
| Coll. Ribes | = | Coll Antoni Ribes Escolà, Torres de Segre, Spain (Now in BMNH) |
A number of samples were obtained through the SMTP. These were invaluable for the study and represent a substantial part of the material from which some of the new species are described.
Descriptions are based on observation of specimens made with Leica MZ9.5 and Leica MZ16 stereo microscopes. Names of morphological structures are based on Fontal-Cazalla et al. (Citation2002), Nordlander (Citation1978) and Nordlander and Grijpma (Citation1991), with additions and modifications by the present authors.
Photographs were taken either with a Canon EOS 7D camera coupled with a MP-E 65 mm lens or with an Infinity X 32 camera attached to a Olympus SZX12 microscope. The software Zerene Stacker was used for stacking images.
Scanning electron microscopy (SEM) images were taken with Zeiss Supra35 VP FEG equipment from the Earth Sciences department of Uppsala University, Sweden.
Measurements were made directly using the stereo microscopes or from photographs.
Systematic treatment
Checklist of Holarctic species of Rhoptromeris
References to all nomenclatural acts not proposed in this paper can be found in Nordlander and Grijpma (Citation1991) and Forshage et al. (Citation2013).
Each valid species is accompanied by an indication of the biogeographical region where it occurs. HA = Holarctic, NA = Nearctic, ePA = East Palearctic, wPA = West Palearctic.
Rhoptromeris Förster, 1869
Rhoptromeris Förster, 1869 type Cothonaspis eucera Hartig, 1841 (= R. heptoma)
Miomoera Förster, 1869 type Miomoera aberrans Förster, 1869 (= R. heptoma)
Hexamerocera Kieffer, 1901 type Eucoila rufiventris Giraud, 1860
Striatellia Belizin, 1966 type Striatellia armeniaca Belizin, 1966
Rhoptromeris acuminata (Belizin, 1968) ePA
Odonteucoila acuminata Belizin, 1968
Rhoptromeris aperta (Kieffer, 1908) NA.
Eucoila (Rhoptromeris) aperta Kieffer, 1908
Rhoptromeris armeniaca (Belizin, 1966) wPA
Striatellia armeniaca Belizin 1966
Rhoptromeris carinata Ionescu, 1969 wPA
Rhoptromeris dichromata sp. nov. wPA
Rhoptromeris heptoma (Hartig, 1840) wPA
Cothonaspis heptoma Hartig, 1840
Cothonaspis biscapa Hartig, 1840
Cothonaspis eucera Hartig, 1841
Cothonaspis tristis Hartig, 1843
Eucoila nodosa Giraud, 1860
Eucoila parvula Thomson, 1862
Miomoera aberrans Förster, 1869
Eucoela (Rhoptromeris) aequalis Kieffer, 1901
Eucoela (Rhoptromeris) graciliclava Kieffer, 1902
Eucoela (Rhoptromeris) eucera var. subnuda Kieffer, 1904
Eucoela (Rhoptromeris) eucera var. pulchricornis Kieffer, 1904
Rhoptromeris widhalmi Kurdjumov, 1912
Eucoila luteicornis Ionescu, 1959 syn. nov. Type examined by MF in MGAB.
Rhoptromeris koponeni sp. nov. wPA
Rhoptromeris leptocornis sp. nov. wPA
Rhoptromeris macaronesiensis sp. nov. wPA
Rhoptromeris miniata (Belizin, 1968) ePA
Pseudeucoila (Hexamerocera) miniata Belizin, 1968
Rhoptromeris nigriventris Nordlander, Citation1978 HA
Rhoptromeris rufiventris (Giraud, 1860) wPA
Eucoila rufiventris Giraud, 1860
Rhoptromeris strobigena Nordlander & Grijpma, Citation1991 wPA
Rhoptromeris villosa (Hartig, 1840) wPA
Cothonaspis villosa Hartig, 1840
Glauraspidia parva Thomson, 1877
Key to the West Palearctic species of Rhoptromeris
Based on the keys by Nordlander (Citation1978) and Nordlander and Grijpma (Citation1991)
Rhoptromeris armeniaca is not included in the key since the poor condition of the only known specimen prevents the visualisation of important diagnostic characters. Its species identity is discussed in the systematic treatment below.
1. Metapleura with an anteroventral cavity (–), posterior margin not depressed medially, or very weakly so .... 2
– Metapleura without an anteroventral cavity (), posterior margin strongly depressed medially .. 7
2. The foveae of the pronotal plate confluent in the middle, forming a transverse furrow (–). Compound eyes and ocelli relatively large... 3
– The foveae of the pronotal plate separated by a medial bridge (). Compound eyes and ocelli relatively small 5
3. Antennae very long, in males 2× body length, in females clearly longer than body (,). Female antennae with an indistinct club of 7–8 elongate segments hardly wider than pre-club segments, each several times longer than wide. Posterior incision of metapleura present (). Wings relatively narrow, marginal cell elongate Rhoptromeris leptocornis sp. nov.
– Antennae less elongate, in males less than twice body length, in females same size or shorter than body, with a more or less distinct club, club segments only a few times longer than wide. Posterior metapleural incision absent (,). Wings relatively broad 4
4. Mesoscutum humped, marginal cell with vein Rs longer than 2r. Female antennae shorter than body, with pre-club flagellomeres brown. Club distinct, 6-segmented, each segment compact, only slightly longer than wide (). Male flagellomeres relatively short, stout () Rhoptromeris strobigena
– Mesoscutum only weakly arched, marginal cell almost 3 times as long as deep, veins 2r and Rs subequal in length. Female antennae as long as body, pre-club flagellomeres yellow or light brown. Club indistinct, 7-segmented, segments relatively elongate, about twice as long as wide (). Male flagellomeres more elongate () Rhoptromeris koponeni sp. nov.
5. Male antennae completely dark, flagellomeres stout, F2 strongly widened and clearly longer than F1 (, ). Femora of fore and middle legs expanded, especially on females (). Female club distinctly 7-segmented (). Legs relatively short Rhoptromeris heptoma
(cf. also the rare Rhoptromeris carinata, known only from a handful of specimens collected in Romania. The species exhibits remarkably short legs and antennae and exceptionally elongate metasoma.)
– Male antennae with F1 and F2 usually lighter than remaining flagellomeres, F2 not much wider than other flagellomeres. Both sexes with relatively long legs, femora of fore and middle legs slender 6
6. Dorsal scutellar surface with reticulate sculpture (), marginal cell elongate, approximately 3 times as long as wide. Female antenna 0.8–0.9× body length, F3 relatively long, often as long as F2 and F4, indistinct club of 7 or 8 segments (). Central club segments about 2.5 times longer than wide. Male F2 distinctly curved Rhoptromeris dichromata sp. nov.
– Dorsal scutellar surface smooth or only weakly sculptured, marginal cell shorter, approximately 2 times as long as wide. Female antenna 0.6–0.7× body length, F3 relatively short, usually much shorter than F2 or F4, distinct 6-segmented club (). Central club segments about 1.5 times longer than wide. Male F2 weakly curved Rhoptromeris villosa
7. The lateral foveae of the pronotal plate separated by a medial bridge (). Scutellar foveae of normal size, scutellar dorsal surface with reticulate sculpture in the posterior part. Scutellum higher, more projected posteriorly, less strongly sloping, with a distinct dorsolateral border. Mesoscutum relatively humped. Compound eyes relatively large Rhoptromeris nigriventris
– The lateral foveae of the pronotal plate confluent in the middle (–). Scutellar foveae very narrow (), scutellar dorsal surface largely smooth. Scutellum remarkably short, low, strongly sloping. Mesoscutum relatively flat. Compound eyes relatively small 8
8. Posterior incision of metapleura present (,) Rhoptromeris macaronesiensis sp. nov.
– Posterior incision of metapleura absent (,) Rhoptromeris rufiventris
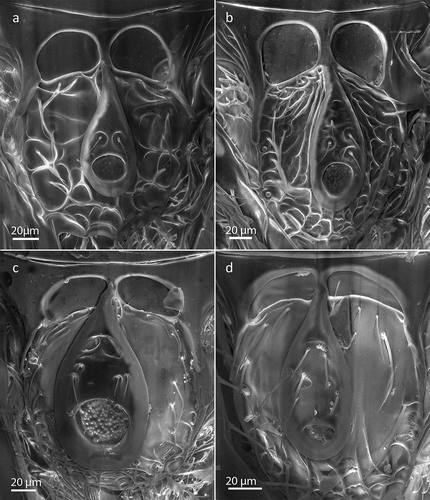
Figure 1. Scanning electron microscopy (SEM) images of the metapleura of (a) Rhoptromeris dichromata sp. nov., (b) R. koponeni sp. nov., (c) R. leptocornis sp. nov. and (d) R. macaronesiense sp. nov. Vertical arrows point to the anterior cavity of the metapleura (present in a, b and c, absent in D) while diagonal arrows indicate the posterior incision of the metapleura (present in c and d, absent in a and b).

Figure 2. Scanning electron microscopy (SEM) images of the pronotal plate of (a) female Rhoptromeris dichromata sp. nov., (b) male R. leptocornis sp. nov., (c) female R. koponeni sp. nov., and (d) female R. macaronesiensis sp. nov.

Figure 3. Habitus of Rhoptromeris dichromata sp. nov. (a: female, b: male), Rhoptromeris koponeni sp. nov. (c: female, d: male), Rhoptromeris leptocornis sp. nov. (e: female, f: male) and Rhoptromeris macaronesiesis sp. nov. (g: female, h: male).

Figure 4. Antennae of (a) female Rhoptromeris dichromata sp. nov., (b) female R. strobigena, (c) female R. villosa, (d) female R. koponeni sp. nov., (e) female R. leptocornis sp. nov., (f) female R. nigriventris, (g) female R. rufiventris, (h) female R. macaronesiensis sp. nov., (i) female R. heptoma, and (j) male R. heptoma.

Figure 5. Habitus of Rhoptromeris heptoma (a: female, b: male), Rhoptromeris villosa (c: female, d: male), Rhoptromeris strobigena (e: female, f: male), Rhoptromeris nigriventris (g: female, h: male) and Rhoptromeris rufiventris (i: female).

Figure 6. Scanning electron microscopy (SEM) images of the scutellum of (a) female Rhoptromeris dichromata sp. nov., (b) female R. koponeni sp. nov., (c) male R. leptocornis sp. nov., and (d) female R. macaronesiensis sp. nov.
Descriptions
Rhoptromeris armeniaca (Belizin), 1966
Diagnosis
Known from a single female deposited in ZIN. Type specimen small, brown, with slender build. Scutellum partly obscured by a glued wing, scutellar plate very narrow, dorsal surface of scutellum smooth. Anteroventral cavity of metapleura present. First flagellomere subequal to second, third flagellomere smaller than first two, fourth subequal to third. Front and middle legs missing.
The poor condition of the type made its study inconclusive. The specimen is most similar to R. villosa due to the small, slender body and narrow scutellar plate, but differs in the proportion of antennal segments. No specimens similar to the holotype were found among the material examined from Western and Northern Europe or from the Macaronesian islands. We suggest that it is either a representative of an eastern species or an aberrant specimen of R. villosa.
Distribution
The holotype and only known specimen was collected in Armenia.
Biology
Rhoptromeris armeniaca is probably a parasitoid of Chloropidae, as are other known Rhoptromeris species.
Rhoptromeris carinata Ionescu, 1969
Diagnosis
Species known from female holotype, one paratype and two additional specimens, all deposited in MGAB. Body small, pale, stout, antennae and legs remarkably short. Globular head with broad genae. Anteroventral metapleural cavity present, posteroventral incision of metapleura absent. Elongate metasoma with lateral hairy ring. Holotype wings damaged, paratype wings missing.
The stout body, short appendages and antennal characters suggest this species is most similar to Rhoptromeris heptoma or possibly a synonym of it. No specimens similar to the holotype were found among the material examined from Western and Northern Europe or from the Macaronesian islands. We suggest that the known individuals are either representatives of an eastern or rare species, or perhaps even aberrant specimens of R. heptoma.
Distribution
All four known specimens were collected in Romania.
Biology
Rhoptromeris carinata is probably a parasitoid of Chloropidae, as are other known Rhoptromeris species.
Rhoptromeris dichromata sp. nov.
(,)
Diagnosis
Head and mesosoma dark brown or black, female metasoma lighter, usually with reddish or orange tinge ventrally, 1.5–1.6 mm (), male metasoma brown. The species can be distinguished from R. heptoma or R. villosa by the long and narrow marginal cell, elongate antennae and more strongly sculptured scutellum, and from R strobigena or R. koponeni by the long and narrow marginal cell, the medial bridge on the pronotal plate and the smaller eyes. Specimens show considerable variation in size of female flagellomeres, height of scutellum and sculpture of the dorsal scutellar surface. It is possible that future molecular studies or analysis of additional material will reveal that the species as conceived here can be split into two or more distinct species.
Description
Compound eyes and ocelli small. Female antenna 0.8–0.9× body length, 13-segmented, scape, pedicel and first three flagellomeres light brown, flagellomeres 4–13 brown. Flagellomeres subequal in size. Indistinct club of 8 articles, bearing rhinaria, not much wider than pre-club flagellomeres. Male antennae approximately 1.6× body length. Flagellomeres elongate, brown, F1 and F2 sometimes lighter. F2 moderately bent, forming a weak ‘S’ shape when viewed from above. F2 from 1.5 to almost 2 times the length of F1.
Mesonotum smooth, with a few scattered setae, weakly arched. Pronotal plate with lateral bridges, foveae large and separated by a medial bridge (). Lateral aspect of pronotum with a few scattered setae. Scutellar plate drop-shaped (), sometimes more elongate, surface flat, with a large posterior glandular pit and small setal pits anteriorly, a few scattered setae. Scutellar foveae of normal size and depth, sometimes more shallow. Lateral bars sometimes with a few striae basally. Dorsal scutellar surface with ridges forming a reticulate pattern, more or less radiating from scutellar plate. Dorsal surface with a line of setae along its rim, separated from lateral surface by a carina. Lateral surface mostly smooth, with a few ridges. Scutellum rather short. Mesopleural line present as a ridge. Metapleura glabrous, except for 2 or 3 long setae on dorsal region. Anteroventral cavity present, round (), filled with hair. Posterior margin straight, with a few weak ridges running anterodorsally. Posterior incision of metapleura absent.
Propodeum pubescent laterally, with tuft of hair above metapleural ventral callus. Propodeal carinae straight, converging dorsally.
Metasoma darker on its dorsal portion, in females often much lighter ventrally, with a yellowish or reddish tinge. Hairy ring with a small dorsal opening, dense, especially in females.
Legs yellow or light brown. Metacoxae with a small posteroproximal tuft of short hair.
Wings relatively narrow, hyaline, pubescent, apex rounded, hair fringe not particularly long. Marginal cell closed or diffusely closed, narrow, elongate, 3 times as long as deep, veins Rs and 2r straight, the same size or with Rs slightly longer. Venation light brown, accessory veins sometimes very lightly indicated by colouration.
Etymology
In allusion to the often yellowish or reddish metasoma which contrasts with the black head and mesosoma.
Distribution
Europe: specimens seen from Austria, Denmark, Finland, Norway, Slovenia, Sweden.
Holotype
Female: SWEDEN, Västerbotten, Vindelns kommun, Kulbäckslidens f.-park, Kulbäcken, meadow, birch wood on fine alluvial sediments, (64.190217°N 19.605700°E), 1–18 August 2003, Swedish Malaise Trap Project (NHRS).
Paratypes
(4 females): SWEDEN: Småland, Nybro kommun, Bäckebo, Grytsjöns naturreservat, 56.931407°N 16.085536°E, Old moist haymaking meadow in forest edge, 27 June–2 July 2005, Swedish Malaise Trap Project (NHRS) (1 female, NHRS); Småland, Nybro kommun, Bäckebo, Grytsjöns naturreservat,56.921656°N 16.101228°E, Old aspen forest in boulder terrain, 2–12 July 2005, Swedish Malaise Trap Project (NHRS) (1 female, NHRS); Halland, Halmstad kommun, Gårdshult, Buskastycket, Moist hay meadow (56.694883°N 13.150500°E) 1–25 April 2004, Swedish Malaise Trap Project (NHRS) (1 female, NHRS); Västerbotten, Vindelns kommun, Kulbäcken meadow, birch wood on fine alluvial sediments, (64.190217°N 19.605700°E), 5–20 August 2004, Swedish Malaise Trap Project (NHRS) (1 female, NHRS).
Additional material
AUSTRIA: Niederösterreich Lunz/See 47.85°N 15.0333°E, 5–14 August 2005 leg H. Malicky (1 female, OLML), FINLAND: Southern Finland, Mikkelin mlk 26 June 1983 M. Koponen leg (1 female, coll Koponen), Savonia australis, Mikkelin mlk 6840:507 28 July 1996 M. Koponen leg (1 male, coll Koponen), Kuusamo, Kuusamo 7371:609 3 July 1998 M. Koponen leg (2 males, coll Koponen) Ks, Kuusamo 30 June 1979 M. Koponen leg (1 male, coll Koponen) Ostrobothnia media, Lappajärvi 7014:327 31 July 1995 (1 male, coll Koponen) Southern Finland, Ristiina 25 June 1983 M. Koponen leg (1 male, coll Koponen); SLOVENIA: Radovljica 2–13 August 1978 malaise trap, Phragmites swamp Lars Huggert (2 females, NHRS); SWEDEN: Småland, Högsby kommun, Hornsö, kronopark, birch fen (57.006550°N 16.109350°E), 20–30 June 2004 (coll. Event ID 340), Swedish Malaise Trap Project (NHRS) (1 female, NHRS); Södermanland, Huddinge kommun, Sofielunds återvinningsanläggning, pine forest with garbage (59.176533°N 17.993850°E), 30 June–13 July 2004, Swedish Malaise Trap Project (NHRS) (1 female, NHRS); Uppland, Älvkarleby kommun, Båtfors. Pine forest with blueberry (60.46065°N 17.317817°E), 14 June–04 July 2005, Swedish Malaise Trap Project (NHRS) (2 females, NHRS); Uppland, Uppsala kommun, Ekdalens naturreservat, tall herbs and young trees mixed with old oaks (59.971517°N 18.354983°E), 7–21 July 2003, Swedish Malaise Trap Project (NHRS) (1 female, NHRS); Skåne, Skäralid, SW of Lierna N56° 01ʹ E13° 13ʹ 18–30 August 1994. Leg M. Söderlund (1 female, MZLU); Västerbotten, Hällnäs, 18 August 1982 Leg K.J. Hedqvist (1 female, MZLU); Hälsingland, Älgesjön, RN689312/152201 Leg Erik Sahlin, Fönsterfälla 15 May–15 June 2002 (2 females, NHRS); Hälsingland, Älgesjön, RN689312/152201 Leg Erik Sahlin, Fönsterfälla 24 July–11 September 2002 (1 female, NHRS).
Biology
Specimens were collected from the end of June to August in mostly forested habitats (both coniferous and deciduous forests, as well as forest edges towards meadows). The relatively narrow wings suggest that the species is not a very good flier and possibly attacks hosts living on low vegetation. Probably a parasitoid of Chloropidae (Diptera), like other known Rhoptromeris species.
Rhoptromeris heptoma (Hartig, 1840)
(,)
Diagnosis
Common species easily identified by the enlarged femora in the fore and middle legs and by antennal characters.
Redescription
Compound eyes relatively small. Male antennae dark, flagellomeres stout, F2 strongly enlarged and clearly longer than F1. Female antennae with F1 subequal to F2 and F3 subequal to F4, the first pair longer than the second. Pre club flagellomeres yellow or light brown, club distinct, 7-segmented, flagellomeres dark. Pronotal fovea separated by a medial bridge. Scutellar dorsal surface weakly sculptured. Anteroventral cavity of metapleura present, posterior incision of metapleura absent. Legs rather short, femora of fore and middle legs expanded. Wings relatively narrow.
Distribution
Very widespread species. Records listed here follow Fauna Europaea, Nordlander (Citation1978) and Quinlan (Citation1986) (African records), with additions by the authors. AFRICA: Democratic Republic of Congo, South Africa. ASIA: China. EUROPE: Austria, British Islands, Croatia, Czech Republic, Denmark, Estonia, Finland, France, Germany, Hungary, Italy (including Sardinia), Lithuania, Norway, Poland, Romania, Russia (uncertain: ‘USSR’ records), Slovakia, Slovenia, Spain, Sweden, Switzerland, The Netherlands, Ukraine.
Biology
Rhoptromeris heptoma is one of the main parasitoids of the agricultural pest Oscinella frit (Linnaeus, 1758). Details about the interaction of R. heptoma with its host are given by Nordlander (Citation1978). The species is readily collected in grassy areas (Nordlander Citation1978).
Rhoptromeris koponeni sp. nov.
(,)
Diagnosis
Head and body chestnut brown to dark brown, metasoma often somewhat paler, 1.2–1.6 mm, marginal cell elongate, approximately 3 times as long as wide. The species is most likely to be confused with R. strobigena, from which it can be distinguished by the longer marginal cell and less humped mesonotum, and by the antennae, which are considerably longer in females and with more elongate flagellomeres in males.
Description
Female antenna the same size as body, 13-segmented, scape, pedicel, flagellomeres 1–3 yellow or light brown, flagellomeres 4–13 brown. Flagellomeres subequal in size, F1 slightly shorter. Indistinct club of 7 articles, bearing rhinaria, each not much wider than pre-club flagellomeres, F4 often a transitional shape and colour. Male antenna 1.8× body size, 15-segmented, light brown or brown. Flagellomeres 1 and 2 somewhat paler, flattened on outer side, second flagellomere longer than neighbour articles, moderately curved, with rhinaria on about two-thirds of its extent. Compound eyes and ocelli large.
Mesonotum, smooth, with a few scattered setae, weakly arched. Pronotal plate with lateral bridges, foveae confluent in the middle forming a transverse furrow ().
Scutellar plate elongate, elliptical, flat to slightly convex with a distinct margin and large posterior glandular pit. A few smaller pits present anteriorly, sometimes with a seta in the middle. Foveae of average size and depth. Lateral bars longitudinally striate basally. Dorsal scutellar surface with striation irradiating from scutellar plate () and running more or less longitudinally, sculpture sometimes irregular near the rim. Dorsal surface with a line of setae along its rim, separated from lateral surface by a carina.
Mesopleural line present as a ridge. Lateral surface of metapleura glabrous, except for 2 or 3 long setae on dorsal region. Anteroventral cavity round, filled with hair (). Surface mostly smooth, sometimes 1 or 2 small ridges arising from posterior margin and running anterodorsally. Posterior margin weakly depressed above ventral callus. Posterior incision of metapleura absent.
Propodeum, more or less pubescent laterally, with tuft of hair behind posteroventral depression on metapleura. Propodeal carinae straight, converging dorsally.
Hairy ring often incomplete dorsally, interruption larger in males. Dense in females, more sparse in males.
Legs yellow, metacoxae elongate, with a small posteroproximal tuft of short hair.
Wings broad, hyaline, pubescent, hair fringe not particularly long. Marginal cell closed, rather deep, elongate, almost 3 times as long as deep, vein 2r sometimes slightly arched, subequal to or slightly shorter than Rs, Rs straight. Venation brown, some accessory veins lightly coloured.
Etymology
Named after the Finnish hymenopterist Martti Koponen, who collected and loaned, from his personal collection, a large number of Rhoptromeris specimens studied for this work.
Distribution
Europe: specimens seen from Czech Republic, England, Finland, Ireland, Scotland, Sweden.
Holotype
Female: FINLAND: Ristiina, 8 July 1978, M. Koponen (coll. Koponen).
Paratypes
(5 females, 4 males): CZECH REPUBLIC: Bohemia mer Vráž u Písku, 28 May 1994, V. Barták (1 male, OLML). FINLAND: SOUTHERN FINLAND, Mikkelin mlk, 20 June 1980, M. Koponen, (1 male, coll. Koponen), Mikkelin mlk, 11 July 1981, M. Koponen (1 male, coll. Koponen), Ristiina, 4 July 1980, M. Koponen (1 female, coll. Koponen); Uusimaa, Espoo, 10 August 1980, M. Koponen (1 female, coll. Koponen); Savonia australis: Mikkelin mlk 6839:506 26 June 1999 M. Koponen leg (1 female, coll. Koponen). SWEDEN: Småland, Nybro kommun, Bäckebo, Grytsjöns naturreservat, 56.931407°N 16.085536°E, old moist haymaking meadow in forest edge, 2–12 July 2005, Swedish Malaise Trap Project (2 females, 1 male, NHRS).
Additional material
FINLAND: SOUTHERN FINLAND, Mikkelin mlk, 15 June 1975 M. Koponen leg (1 male, coll. Koponen), Mikkelin mlk, 1 August 1981 M. Koponen leg (1 female, coll. Koponen); Ostrobothnia media: Lappajärvi 7020:331 31 July 1995 M. Koponen leg (1 male, coll. Koponen); Nylandia: Nurmijärvi 6704:381 9 July 1992 M. Koponen leg (1 female, coll. Koponen); Savonia borealis: Sonkajärvi 7082:521 8 July 2000 M. Koponen leg (1 male, coll. Koponen); Tavastia borealis: Hankasalmi museo 6909:3466 11 July 2009 M. Koponen leg (1 male, coll. Koponen); SWEDEN Skåne, Revinge s:n, N Krankesjön (VB0440 7470) 23 June 1993 (14.45–16.00) Moist birch stand, surrounded by oak; sweepnetting. Leg Michael Söderlund (1 male, MZLU), Ivö 9 June 1949 Leg Tord Nyholm? (1 female, SLU), Småland, Nybro kommun, Bäckebo, Grytsjöns naturreservat, 56.931407°N 16.085536°E, Old moist haymaking meadow in forest edge, 2–12 July 2005, Swedish Malaise Trap Project (1 male, NHRS).
Biology
Specimens collected from end of May to beginning of August. In Sweden, specimens were captured in a meadow along a forest edge. One specimen was collected in a boggy birch grove surrounded by oak. Probably a parasitoid of Chloropidae (Diptera), like other Rhoptromeris species.
Rhoptromeris leptocornis sp. nov.
(,)
Diagnosis
Species with characteristic colour pattern of dark brown head, mesonotum and scutellum contrasting with light or chestnut brown remainder of mesosoma (,). Colour contrast less evident in very dark or pale specimens. Body length 1.0–1.5 mm, marginal cell elongate, vein 2r somewhat longer than Rs. Distinguishable from other European species by the very long antennae which are longer than the body on females and twice as long as the body on males, large scutellar plate and presence of posteroventral cavity on metapleuron. Studied specimens from Turkey and Greece were smaller and paler than those from other areas.
Description
Antennae filiform, elongate, approximately 1.3× body size in females, 2.0× in males. Female antenna 13-segmented with scape and pedicel yellow or light brown, first two flagellomeres yellow, flagellomeres 3–13 light brown or brown. Flagellomeres several times longer than wide and subequal in length, except for first flagellomere somewhat shorter. Indistinct club of 7–8 segments only slightly wider than pre-club flagellomeres, rhinaria often hard to distinguish. Male antenna 15-segmented with scape, pedicel and first two flagellomeres yellow or light brown, flagellomeres 3–15 gradually darker until light brown to brown. Second flagellomere modified, longer than neighbour articles, somewhat flattened, moderately curved. Compound eyes and ocelli large.
Mesonotum, smooth, glabrous, moderately arched. Pronotal plate with lateral bridges, foveae confluent in the middle forming a transverse furrow (). A small lateral incision on either side of the furrow, which could possibly be mistaken for a laterally open fovea as in the majority of other genera of Eucoilinae.
Scutellar plate elevated, large, drop-shaped, posteriorly protruding to hind margin of scutellum. Dorsal surface flat to slightly concave, with a distinct margin, a few scattered setae and a very large posterior glandular pit. Scutellar foveae relatively narrow, shallow (). Lateral bars smooth. Dorsal scutellar surface smooth or longitudinally striate, striation stronger posteriorly. Dorsal surface with a line of setae along its rim, separated from lateral surface by a carina. Lateral surface smooth. Scutellum strongly sloping posteriorly.
Mesopleural line present. Lateral surface of metapleura glabrous, except for 2 or 3 long setae on dorsal region. Anteroventral cavity round, filled with hair, sometimes completely enclosed by sclerite. Surface smooth except for oblique ridge running anterodorsally from approximately one-third the height of posterior margin. Posterior margin with a circular incision below the ridge (posterior incision of metapleura (). Area around incision often somewhat depressed. Posterior corner of metapleura pointed, sometimes slightly upcurved.
Propodeum more or less pubescent laterally, tuft of hair behind posteroventral metapleural cavity. Propodeal carinae slightly converging dorsally, sometimes weakly arched on ventral portion.
Metasoma chestnut brown to dark brown, darker on its posteroventral half. Hairy ring often incomplete dorsally, dense in females, sparse in males.
Legs long and slender, yellow, sometimes light brown. Metacoxae elongate, with a small posteroproximal tuft of short hair.
Wings relatively narrow, hyaline, pubescent, apex rounded with a rather long hair fringe. Marginal cell closed, narrow, elongate, at least twice as long as deep, vein 2r somewhat longer than Rs. Venation yellow or light brown, accessory veins visible as spectral veins.
Etymology
From the greek ‘lepto-’ meaning thin, slender, and the latin ‘-cornis’, horns. Refers to the thin, elongate antennae.
Distribution
Western Palearctic: specimens seen from Canary Islands, Croatia, France, Germany, Greece, Madeira archipelago, Norway, Spain and Turkey. A large number of specimens were collected at the Macaronesian islands, while only isolated individuals were seen from the continent.
Holotype
Female: MADEIRA ISLAND – Portugal, Ribeiro Serrao, Proj. Meia Serra, 6 October 2004, Plant: Erica scoparia, No beat: 2(40) (MNCN).
Paratypes
(46 females, 30 males): CANARY ISLANDS: TENERIFE, El Batan, 26 October 1996, M. Báez (1 male, coll Koponen); La Laguna, Anaga Mts, Las Mercedes-Wald, 889 m, 28.5303°N 16.2878°W, 16 October 2008 (1 female, OLML); Puerto de la Cruz, Taoro, 11 December 1986, M. Koponen (1 male, coll Koponen). MADEIRA ISLAND: Funchal, Curral das Romeiros, (500–600 m), 8 December 1991, M. Koponen (1 female, coll. Koponen); Funchal, Curral das Romeiros, (500–600 m), 22 December 1995, M. Koponen (1 female, 1 male, coll. Koponen); Funchal, Old Monte Gardens, 550 m, 5 November 1996, M. Koponen (28 females, 20 males, coll Koponen); Funchal, Palheiro Ferreiro, (550–600 m), 22 December 1995, M. Koponen (1 female, 3 males, coll. Koponen); Funchal, Quinta do Pomar, (600 m), 22 December 1995, M. Koponen, (5 females, 1 male, coll. Koponen); Jardim da Serra, 10 December 1991, M. Koponen, (1 male, coll. Koponen); Monte, (550–750 m) 17 December 1994, M. Koponen (3 females, 2 males, coll. Koponen); Monte, (600 m) 8 November 1996, M. Koponen (1 female, coll. Koponen); Monte, (600 m) 3 November 1996, M. Koponen (2 females, coll. Koponen); Ribeiro Serrao, 6 October 2004 (3 females, MNCN).
Additional material
CROATIA: Zadar, Borik, 15 July 1961, Stellan Erlandsson (1 male, SLU). GREECE: Kerkini Lake National Park, yellow pan trap, Megalohori site, 24–29 August 2008, Gordon Ramel (1 female, NHRS); NORWAY: Oslo, Ekebergskråningen, 29 May–27 June 2007, A. Endrestøl/Lars Ove Hansen (1 female, ZMUN); SPAIN: Alcoletge, Lleida, 31T CG01, 160 m 19 July 2007 ‘Sweeping on grasses and flowering vegetation, lowland Populus riparian forest’ Leg Antoni Ribes (2 males, coll Ribes); La Mitjana, Lleida 31T CG01, 145 m, 22 June 2007 ‘Sweeping on flowering Dorycnium pentaphyllum’ Leg Antoni Ribes (1 male, coll Ribes); TURKEY: Side. 1–12 June 1992 Gardens and pasture. Light trap Leg Gösta Gillerfors (1 female, MZLU).
Biology
Specimens were collected between June and August on the continent and from October to December on Madeira and Canary Islands, at elevations ranging between 89 and 889 m. In Madeira, individuals were swept from Erica scoparia, Origanum, Quercus and Vaccinium, while in Spain specimens were captured on Dorycnium pentaphyllum and among grasses and flowering vegetation in a riparian forest. Probably a parasitoid of Chloropidae (Diptera), as are other Rhoptromeris species.
Rhoptromeris macaronesiensis sp. nov.
(,h)
Diagnosis
Body brown, head somewhat darker than body and metasoma with a rufous tinge anteroventrally, legs yellow, 1.0–1.1 mm. The species is most similar to R. rufiventris, from which it can be separated by the distinct round incision on the posterior metapleural margin.
Description
Female antennae approximately 0.8× body size, 13-segmented, scape and pedicel yellow or light brown, flagellomeres gradually darker until brown. Indistinct club of 6 segments, rhinaria sometimes hard to distinguish. Male antenna 15-segmented with scape and pedicel yellow or light brown, flagellomeres brown, rather slender. Second flagellomere modified, not much longer than third, weakly curved. Compound eyes and ocelli small.
Mesonotum smooth, with a few scattered setae, weakly arched. Pronotal plate with lateral bridges, foveae confluent in the middle forming a transverse furrow ().
Scutellar plate elongate, eliptical. Dorsal surface flat, with a distinct margin, a few scattered setae and a relatively small posterior glandular pit. Scutellar foveae narrow, shallow (). Lateral bars smooth. Dorsal scutellar surface smooth anteriorly and with light striation posteriorly. Dorsal surface with a line of setae along its rim, separated from lateral surface by a carina. Lateral surface smooth. Scutellum very short, sloping posteriorly.
Mesopleural line present. Lateral surface of metapleura smooth, glabrous, except for 2 or 3 long setae on dorsal region. Anteroventral cavity absent. Posterior incision of metapleura present (). Area around the incision sometimes more depressed than remainder of metapleura.
Propodeum more or less pubescent laterally, tuft of hair behind posteroventral metapleural cavity. Propodeal carinae slightly converging dorsally, sometimes weakly arched.
Metasoma with a rufous tinge anteroventrally. Hairy ring incomplete or much more sparse dorsally, more dense in females.
Metacoxae rather elongate, with a small posteroproximal tuft of short hair.
Wings relatively narrow, hyaline, pubescent, apex rounded with a rather long hair fringe. Marginal cell closed or diffusely closed, 3 times as long deep, vein 2r somewhat longer than Rs. Venation light brown, accessory veins not visible.
Etymology
In reference to the fact that most studied specimens of this species were collected from the Macaronesian islands.
Distribution
West Palearctic: specimens seen from Canary Islands, Denmark, Madeira archipelago.
Holotype
Male: MADEIRA: Monte, 600 m, 3 November 1996, M. Koponen (coll Koponen).
Paratypes
(6 females, 3 males): CANARY ISLANDS: La Palma, Breña Baja, Los Cancajos Hac. S. Jorge, 22 February 1997, M. Koponen (1 female, coll Koponen). MADEIRA: Monte, 550–750 m, 17 December 1994, M. Koponen (1 female, coll Koponen); Babosas, 550 m, 7 December 1991, M. Koponen (1 male, coll Koponen); Romeiros, 11 May 1980, M.W.R. de V. Graham coll (1 male, BMNH (E) 1995-177); Romeiros, 20 July 1982, M.W.R. de V. Graham coll (1 male, BMNH (E) 1995-177); Curral dos Romeiros, 20 August 1982, M.W.R. de V. Graham coll (1 female, BMNH (E) 1995-489); Cruzinhas do Faisl, 11 August 1987, on lime trees, M.W.R. de Graham coll (1 female, BMNH (E) 1995-489); Romeiros, 11 May 1980, E.M. Graham, M.W.R. de V. Graham coll (1 female, BMNH (E) 1995–177); Lombada dos Vacas, 11 August 1985, M.W.R. de V. Graham coll (1 female, BMNH (E) 1995-177).
Additional material
DENMARK: Jylland, Grene 3 July 1984 (1 female, coll Munk).
Biology
Specimens collected in November, December, February, May, July and August in Madeira and Canary Islands, height between 550 and 750 m. One specimen collected on lime trees). The only individual from Denmark was captured in July. Probably a parasitoid of Chloropidae (Diptera), as is known for other Rhoptromeris species.
Rhoptromeris nigriventris Nordlander, Citation1978
(,h)
Diagnosis
This species shares with R. rufiventris and R. macaronesiensis sp. nov. the lack of the anteroventral cavity of the metapleura. It can be distinguished by the pronotal foveae separated by a medial bridge, larger eyes, broader wings and higher scutellum with relatively broad scutellar plate.
Several studied specimens exhibited a mix of diagnostic characters of both R. nigriventris and R. rufiventris, and more extensive molecular studies could be useful to better elucidate the relationship between these species.
Redescription
Compound eyes relatively large. Antennae rather long in both sexes. Male F2 enlarged, strongly curved. Female antennae with an indistinct 7-segmented club. Pronotal foveae separated by a medial bridge. Dorsal scutellar surface usually smooth, sometimes more or less strongly sculptured in the posterior half. Scutellar plate large. Anteroventral cavity of metapleura absent, posterior incision of metapleura absent. Wings relatively broad.
Distribution
Widespread throughout Europe. Records listed here follow Fauna Europaea and Nordlander (Citation1978), with additions by the authors. EUROPE: Bosnia, England, Finland, Germany, Italy, Poland, Spain, Sweden.
Biology
Specimens of Rhoptromeris nigriventris have been collected in field and forest areas. The species is probably a parasitoid of grass-inhabiting Chloropidae (Nordlander Citation1978).
Rhoptromeris rufiventris (Giraud, 1860)
()
Diagnosis
Body brown, anteroventral portion of metasoma with a reddish tinge, this colour more pronounced on females. The species is most similar to R. macaronesiensis sp. nov., from which it can be distinguished by the absence of the posterior incision of the metapleura. R. rufiventris can further be separated from R. nigriventris by the pronotal fovea fused medially into a continuous furrow (separated by a medial bridge in R nigriventris), narrow scutellar fovea and very short scutellum.
Several studied specimens exhibited a mix of diagnostic characters of R. nigriventris and R. rufiventris, and more extensive molecular studies could be useful to better elucidate the relationship between both species.
Redescription
Female: Compound eyes small. Pronotal fovea fused medially forming a transverse furrow. Antennae approximately 0.7 times body length, club 7-segmented. Dorsal scutellar surface smooth, scutellar plate narrow. Scutellum short, sloping steeply posteriorly after the scutellar plate. Anteroventral cavity of metapleura absent, posterior incision of metapleura absent. Wings narrow.
Male unknown.
Distribution
Widespread throughout Europe. Records listed here follow Fauna Europaea and Nordlander (Citation1978), with additions by the present authors. EUROPE: Austria, Spain and Sweden.
Biology
Rhoptromeris rufiventris has been collected by sweeping on lawns, and at least one specimen was captured in a pitfall trap placed in an oat field (Nordlander Citation1978). Probably a parasitoid of grass-inhabiting Chloropidae (Nordlander Citation1978).
Rhoptromeris strobigena Nordlander & Grijpma, Citation1991
(,)
Diagnosis
Most similar to R. koponeni, with which it shares the pronotal fovea fused into a continuous furrow, large compound eyes and broad wings. It can be distinguished by the antennae, shorter on females and with shorter flagellomeres on males, as well as by the more strongly humped mesoscutum and the shorter marginal cell with vein Rs longer than 2r (veins subequal in R. koponeni).
Redescription
Compound eyes relatively large. Male antennae elongate, F1 and F2 lighter than remaining flagellomeres. Female antennae short, pre club flagellomeres light brown to brown, club darker, distinct, 6-segmented. Pronotal fovea fused medially forming a transverse groove. Dorsal scutellar surface costulate, scutellar plate broad. Anteroventral cavity of metapleura present, posterior incision of metapleura absent. Wings relatively broad, marginal with vein Rs longer than 2r.
Distribution
Widespread throughout Europe. Records listed here follow Fauna Europaea and Nordlander and Grijpma (Citation1991), with additions by the authors. EUROPE: Finland, France, Italy, Poland, Spain, Sweden, The Netherlands.
Biology
Rhoptromeris strobigena has been reared from puparia of Gaurax and Hapleginella spp. (Diptera: Chloropidae) collected from cones of Abies procera, Larix decidua and Larix kaempferi (Nordlander & Grijpma, Citation1991). The biology and courtship behaviour of the species are discussed by Nordlander and Grijpma (Citation1991).
Rhoptromeris villosa (Hartig, 1840)
(,)
Diagnosis
A common and remarkably variable species in terms of colouration, size, sculpture of scutellum and size of scutellar plate. It is possible that large-scale molecular studies will eventually reveal that the species as currently circumscribed comprises a complex of closely related species. The short female antennae with F3 smaller than F4 and the male antennae with almost straight F2 separate R. villosa from all other Rhoptromeris species. The species can be further distinguished from R. heptoma by the slender femora, and from R. dichromata by the smooth scutellar surface.
Redescription
Compound eyes small. Male antennae brown with the first two flagellomeres light brown or yellow, F1 nearly straight. Female antennae short, F1 subequal to F2, both longer than F4, F3 smaller than F4. Pre-club flagellomeres yellow or light brown, club distinct, dark, 6-segmented. Pronotal foveae separated by a medial bridge, dorsal surface of scutellum smooth, scutellar plate narrow. Anteroventral cavity of metapleura present, posterior incision of metapleura absent. Wings narrow, females sometimes brachypterous.
Distribution
Widespread throughout Europe. Records listed here follow Fauna Europaea and Nordlander (Citation1978), with additions by the authors. EUROPE: Bulgaria, Croatia, Denmark, England, Finland, Germany, Greece, Italy, Norway, Poland, Spain, Sweden. MACARONESIA: Madeira Islands.
Biology
Rhoptromeris villosa has been collected mostly on grass, particularly in sandy areas. Probably a parasitoid of grass-inhabiting Chloropidae (Nordlander Citation1978).
Discussion
Most Rhoptromeris species are considered to be poorly adapted for flight due to the narrow wings and small eyes (Nordlander and Grijpma Citation1991), a hypothesis supported by the fact that specimens are most often collected in grassy areas or among low-growing vegetation, where they presumably find their hosts (Nordlander Citation1978). The one known exception is R. strobigena, which attacks flies in conifer cones and has relatively broad wings, large compound eyes and a humped mesoscutum (Nordlander and Grijpma Citation1991), presumably due to the insertion of large wing muscles. The new species R. koponeni sp. nov. shares with R. strobigena the rather large wings and compound eyes, and for this reason we hypothesise it may also be a relatively good flier which attacks tree-inhabiting hosts.
Among the new taxa described in the present study, R. dichromata sp. nov. and R. macaronesiensis sp. nov. follow the general Rhoptromeris pattern of small eyes and narrow wings, indicating they probably search for hosts near the ground. Rhoptromeris leptocornis sp. nov., however, exhibits relatively large eyes and ocelli, but rather narrow wings. Possible explanations for this combination of characters include nocturnal habits, preference for hosts which can only be located through developed visual capacity, or a shift to a less flight-demanding lifestyle in a lineage of originally good fliers. Further ecological and phylogenetic studies would be necessary to better elucidate the matter, however.
Two of the new species, R. leptocornis sp. nov. and R. macaronesiensis sp. nov., also display a character so far not observed in other Palearctic Rhoptromeris, but which is present in several African species: the posterior metapleural incision. Various Eucoilinae have the median part of the posterior edge of the metapleura depressed, often to the point of interrupting a posterior carina when this feature is present. It is only in a few not so closely related groups that the depression has changed into a distinctly circular incision such as observed in these two species and also in many other Rhoptromeris, Ganaspis Förster, 1869 and all Areaspis Lin, 1988. In Areaspis the incision is even more developed, often quite large and delimited by sharp edges. It is unclear if this feature has any adaptive function and whether some kind of evolutionary pressure may have driven its parallel evolution in different Eucoilinae lineages.
Among the west Palearctic material analysed for this study, several specimens were found which could not be confidently assigned to any of the described species. These corresponded mostly to singletons or morphotypes represented by very few individuals, and we considered it better for the taxonomy of the group to regard them as extremes of morphological variation until more material becomes available. On the subject of Rhoptromeris diversity, it should be noted that the Afrotropical region still appears to be the centre of greatest radiation of the Trichoplastini. Not only are Angustacorpa, Nanocthulhu and Stentorceps endemic to the region, but Rhoptromeris and Trichoplasta also exhibit a remarkable variety of forms, many of which remain undescribed. It would be interesting to see future studies developing phylogenetic analyses with large samples of African and European taxa, as this could greatly improve our understanding of how the different species groups of both continents evolved.
Acknowledgements
We would like to thank the following people for their support and hospitality during museum visits and for providing specimens for the study: Lars Ove Hansen (ZMUN), Fritz Gusenleitner, Martin Schwarz (OLML), Roy Danielsson, Christoffer Fägerström, Christer Hansson (MZLU), Melania Stan (MGAB), José Luis Nieves Aldrey (MNCN), Oleg Kovalev (ZIN), Göran Nordlander (SLU), Antoni Ribes Escolà†, Martti Koponen and Thorkild Munk†. Milos Bartol, from the Department of Earth Sciences, Uppsala University, kindly assisted with the SEM imaging. We are also indebted to the SMTP staff, who sorted countless malaise trap samples from which a large number of specimens were obtained. We thank Matt Buffington and Simon van Noort for reviewing this manuscript and helping us improve it by providing valuable comments and suggestions. Finally, we would like to thank Göran Nordlander, whose work set the foundations that made this study possible.
Disclosure statement
No potential conflict of interest was reported by the authors.
Related Research Data
References
- Buffington MB, Nylander JAA, Heraty JM. 2007. The phylogeny and evolution of Figitidae (Hymenoptera: Cynipoidea). Cladistics. 23:1–29.
- Fontal-Cazalla FM, Buffington ML, Nordlander G, Liljeblad J, Ros-Farré P, Nieves-Aldrey JL, Pujade Villar J, Ronquist F. 2002. Phylogeny of the Eucoilinae (Hymenoptera: Cynipoidea: Figitidae). Cladistics. 18:154–199.
- Forshage M, Nordlander G. 2008. Identification key to European genera of Eucoilinae (Hymenoptera, Cynipoidea, Figitidae). Insect Syst Evol. 39:341–359.
- Forshage M, Nordlander G, Buffington M. 2013. Eucoilinae of North America: a revised catalog of genera and described species. Proc Entomol Soc Wash. 115:225–255.
- Forshage M, Nordlander G, Ronquist F. 2008. Quasimodoana, a new Holarctic genus of eucoiline wasps (Hymenoptera, Cynipoidea, Figitidae) with a phylogenetic analysis of related genera. Syst Entomol. 33:301–318.
- Kovalev OV. 1989. [A new tribe and a new genus of eucoilid wasps (Hymenoptera, Cynipoidea, Eucoilidae)]. Trudy Zoologitseskogo Instituta an SSSR Leningrad. 188:109–113. Russian.
- Nordlander G. 1978. Revision of the genus Rhoptromeris Förster, 1869 with reference to north-western European species. Studies on Eucoilidae (Hym.: Cynipoidea) II. Entomol Scand. 9:47–62.
- Nordlander G, Grijpma P. 1991. Systematics and biology of Rhoptromeris strobigena sp. n., a parasitoid of chloropids inhabiting conifer cones (Hymenoptera: Cynipoidea: Eucoilidae). Entomol Scand. 22:209–218.
- Quinlan J. 1986. A key to the Afrotropical genera of Eucoilidae (Hymenoptera), with a revision of certain genera. Bull Br Mus Nat Hist Entomol. 52:243–366.