
ABSTRACT
Most men show sexual arousal to one, preferred sex, whereas most women respond to both sexes, regardless of their sexual orientation. A different research program indicates that men have lower second-to-fourth finger length ratios (2D:4D) than women, possibly because men are exposed to higher levels of androgens during prenatal development. We hypothesized that sex differences in sexual arousal patterns are influenced by prenatal androgen exposure and would thus be explained by sex differences in 2D:4D. We measured the sexual response patterns of 139 men and 179 women via genital arousal and pupil dilation to erotic videos, in addition to their 2D:4D. Compared to women, men showed stronger responses to one sex over the other, although this pattern was clearer in genital arousal than pupil dilation. Men also had lower 2D:4D than women. However, there was no evidence that sex differences in sexual arousal related to sex differences in 2D:4D. Thus, whichever factor explains sex differences in sexual arousal patterns may not be reflected in 2D:4D.
Research measuring sexual responses to explicit stimuli has uncovered a robust difference between the sexes. Most men show a strong sexual response to stimuli featuring their preferred sex, and little to no response to stimuli featuring their non-preferred sex. In contrast, women tend to respond equally to stimuli featuring either their preferred or non-preferred sex regardless of their sexual orientation (Bailey et al., Citation2016). This sex difference in sexual response is highly robust, and has been detected using measures of genital arousal (Rieger et al., Citation2015; Suschinsky et al., Citation2009), pupil dilation (Attard-Johnson et al., Citation2016; Rieger et al., Citation2015; Rieger & Savin-Williams, Citation2012), viewing time (Ebsworth & Lalumière, Citation2012; Israel & Strassberg, Citation2009), and neural responses (Safron et al., Citation2019; Sylva et al., Citation2013) to sexual stimuli. For this sex difference, a specific pattern of sexual arousal to one sex over the other has sometimes been described as “male-typical,” and a non-specific pattern of arousal to both sexes as “female-typical” (Chivers et al., Citation2007; Rieger et al., Citation2016). There are exceptions to this general sex difference. In men, sexual responses to one, preferred gender is more pronounced in heterosexual and homosexual men than bisexual men (Jabbour et al., Citation2020). In women, nonspecific responses to both sexes are more common in heterosexual women than bisexual or homosexual women (Rieger et al., Citation2016). In general, however, nonspecific sexual arousal to both genders characterizes women more than men (Bailey, Citation2009).
Several (not necessarily mutually exclusive) theories seek to explain this nonspecific pattern of sexual arousal in women (see review by Chivers, Citation2017), including that women’s sexuality may depend more on contextual and social factors than men’s (Baumeister, Citation2000; Diamond, Citation2003), or that their sexual responses may change according to their levels of fertility, which does not occur in men (Gangestad & Simpson, Citation2000). The most prominent proposal is the “preparation hypothesis” (Suschinsky & Lalumière, Citation2011). This hypothesis is based on the observation that forced copulation is common in many species, including humans (Galdikas, Citation1985; Palmer, Citation1989). Because forced copulation can lead to genital injury (Slaughter et al., Citation1997), women may have evolved to respond to any sexual situation with sexual arousal, as it leads to protective lubrication that minimizes the risks of genital trauma. The need for such a protective mechanism may have been so strong that women have evolved to respond to any sexual situation, perhaps especially if it contains cues to possible vaginal penetration, even if unwanted or unpleasant (Bossio et al., Citation2014). Consistent with this hypothesis, heterosexual women, on average, show similar responses to all sexual stimuli, including those featuring violent, nonconsensual sex, whereas heterosexual men show the strongest genital response to stimuli depicting consensual, nonviolent sexual intercourse (Suschinsky & Lalumière, Citation2011). However, more recent findings challenged this hypothesis. Although women show an increase in vaginal pulse amplitude to both sexes, they only lubricate to their preferred sex (Sawatsky et al., Citation2018). Yet, it is possible that increased vaginal pulse amplitude is a necessary pre-cursor for lubrication, but it only leads to lubrication when sexual activity is seemingly imminent, due to strong subjective arousal or the presence of a sexual threat. Thus, the preparation hypothesis remains the favored explanation for the nonspecific sexual arousal patterns in women (see review by Lalumière et al., Citation2020).
Unlike the aforementioned work, the present study did not focus on an ultimate explanation for the sex difference in sexual response. Instead, we examined potential developmental differences between men and women, to determine whether this could explain the difference in their sexual responses. In mammals, exposure to prenatal androgens – specifically testosterone – is thought to account for the majority of sex differences in brain and behavior (Breedlove, Citation2017; Swift-Gallant et al., Citation2020). For instance, individuals with an XY karyotype and complete androgen insensitivity syndrome (CAIS), which results in insensitivity to androgens throughout the lifespan, typically report sexual orientations toward men with the same frequency as genetic women (Wisniewski et al., Citation2000), and do not differ from genetic females in a wide range of psychosexual measures, such as gender identity and gender role behavior in both childhood and adulthood (Hines et al., Citation2003). Additionally, individuals with XY karyotype and CAIS show neural responses to both male and female sexual stimuli, and therefore have female-typical (nonspecific) sexual responses (Hamann et al., Citation2014). Thus, if levels of early androgen exposure influence the development of sex differences, in general (Breedlove, Citation2010; Motta-Mena & Puts, Citation2017; Puts & Motta-Mena, Citation2018), then they could possibly also affect sex differences in the specificity of their sexual arousal.
Since the direct measurement of androgen exposure in utero is difficult to achieve in humans, the majority of research relies on biomarkers – lasting indications on the body that are thought to reflect degree of androgen exposure during fetal development. Of these, the easiest to measure is the ratio of the length of the second digit to that of the fourth digit (2D:4D). Reviews and meta-analyses indicate that men have, on average, lower (more masculine) ratios than women, with the proposal that men’s exposure to higher levels of androgens during fetal development drives this sex difference in digit ratio (Grimbos et al., Citation2010; Hönekopp et al., Citation2007; Swift-Gallant et al., Citation2020; Xu & Zheng, Citation2015).
It is worth noting that 2D:4D is a controversial measure, with ongoing debates about causation (McCormick & Carré, Citation2020; Swift-Gallant et al., Citation2020) and validity of any associated findings due to noise in the data (Bailey et al., Citation2016). However, much of this controversy pertains to the relationship between 2D:4D and sexual orientation, rather than sex differences in 2D:4D. Whereas a previous meta-analysis suggested publication bias with respect to the link of 2D:4D with sexual orientation – resulting in a potential overestimation of the strength of the relationship between the two – the same meta-analysis found no evidence of publication bias with regards to a sex difference in 2D:4D (Grimbos et al., Citation2010). Additionally, in individuals with XY karyotype and CAIS, finger length ratios are feminized, similar to those of unaffected individuals with XX karyotype (Berenbaum et al., Citation2009). Thus, in the complete absence of androgens, it appears that individuals with XY karyotype develop female-typical 2D:4D along with female-typical psychosexual traits (Hines et al., Citation2003). In addition, individuals with an XX karyotype and Congenital Adrenal Hyperplasia, which results in excessive androgen exposure throughout the lifespan, have more male-typical finger length ratios than unaffected females (Brown et al., Citation2002). That being said, there has been debate in the literature about whether any differences in 2D:4D, including sex differences, are confounded by overall digit length: As humans are a sexually dimorphic species with males being larger than females (Kurki, Citation2011) and having longer finger digits than females, on average (Kratochvíl & Flegr, Citation2009), the impact of finger length on 2D:4D could be substantial. As such, in the present research we made an effort to control for finger length, in an attempt to ensure that any detected differences in 2D:4D are not confounded by finger length.
In sum, the literature suggests a sex difference in sexual arousal patterns, with men generally showing strong responses to one sex over the other, whereas women show, compared to men, more equal responses to both sexes, regardless of their own sexual orientation. This sex difference in sexual arousal patterns may be driven by sex differences in prenatal androgen exposure, which may themselves be reflected by 2D:4D.
We therefore tested the following hypotheses:
Men’s sexual arousal patterns will be more sex-specific, on average, whereas women’s sexual responses will be less sex-specific.
On average, men will have a lower 2D:4D than women.
Sex differences in the specificity of sexual arousal will be associated with a sex difference in 2D:4D.
Method
Participants
Based on estimates from pooled previous data produced by our research group, we predicted that the effect size (Cohen’s d) of the sex difference in employed variables of genital arousal to be d= 1.18. For the corresponding sex difference in pupil dilation we expected d= 1.00 (Rieger & Savin-Williams, Citation2012). For the sex difference in 2D:4D, we based our estimates on those published in a meta-analysis (converted from Hedge’s g to Cohen’s d), and anticipated that the effect size would be approximately d = .55 for the right hand, and d= .44 for the left hand. Power analyses in G*Power 3.1.9.7 (Faul et al., Citation2007) estimated, for the smallest of these effects (d= .44) a minimum of 220 participants to achieve significant results with a power of .90. With regards to the association analysis, estimating the necessary sample size proved difficult, as no other study has conducted an association analysis in the same manner as the present study. We therefore erred on the side of caution with participant numbers: Our power analysis for the main effect was based on the more conservative power value of .90 rather than the commonly-used .80, resulting in a sample size requirement of 220 instead of 166 for the smallest expected main effect (d = .44). Additionally, we continued recruiting past this figure as participants were visiting our laboratory for other studies, resulting in a final sample size of 318 – substantially larger than that recommended by the power analysis.
We recruited participants in the United Kingdom via pride festivals in Colchester, Chelmsford, London, and Norwich, via online magazines, and university fairs and mailing lists. Participants reported their sexual orientation using a 7-point scale (Kinsey et al., Citation1953). The 139 recruited men self-identified as “exclusively straight” (n = 33), “mostly straight” (n = 10), “bisexual leaning straight” (n = 4), “bisexual” (n = 10), “bisexual leaning gay” (n = 12), “mostly gay” (n = 11), or “exclusively gay” (n = 59). The 179 women self-identified as “exclusively straight” (n = 37), “mostly straight” (n = 25), “bisexual leaning straight” (n = 9), “bisexual” (n = 15), “bisexual leaning lesbian” (n = 11), “mostly lesbian” (n = 41), or “exclusively lesbian” (n = 41). The mean (SD) age was 24.45 (8.85) for men and 24.18 (7.21) for women. For men, 83% were White, followed by 3% Chinese, 3% Indian, and other ethnicities. For women, 79% were White, 5% Chinese, 4% Black, and other ethnicities.
Only participants for whom valid 2D:4D data were available were entered into the current sample. As such, 2D:4D data were available for all 318 participants, apart from one female participant whose data were lost for the right hand. Some participants opted out of the genital arousal component, and independent of this, we experienced pupil data loss because of problems with the apparatus. Consequently, genital arousal data were available for 305 of the 318 participants, and pupil data for 273 of the 318 participants. Thus, the number of participants varies across analyses, and the specific number of participants included in each stage of analysis is listed in the caption of the corresponding table.
Measures and Materials
Self-reported Sexual Orientation
Participants reported both their sexual orientation and sexual attraction to men and women on 7-point scales (Kinsey et al., Citation1953). These scales were highly correlated in both men p< .0001, r= .98, 95% CI [.98, .99] and women, p< .0001, r= .97 [.96, .98], and therefore averaged within participants. For this average, a score of 0 represented exclusive heterosexuality, and 6 represented exclusive homosexuality. This composite score was used for all analyses.
2D:4D
Digit measurements were taken from either high-resolution photographs or scans of participants’ hands. For the photographs, participants placed their hands on a flat surface in a supinated (palms facing up) position, with their fingers slightly spread apart, and images were taken from approximately 30 cm above this surface. For the scans, participants placed their hands flat in a pronated (palms facing down) position on the surface of the scanner. In both cases, the palmar surfaces of the hands were visible in the resultant images. Different methods of capturing images (photograph or scanner) did not moderate the relationship between sex and 2D:4D.
Using these images, digit ratios were measured by two independent raters who were blind to the participants’ sex and sexual orientation. Measurements were performed with the vector graphics package Inkscape 0.92, as computer-assisted techniques produce the most reliable measurements (Allaway et al., Citation2009). Each rater drew a line as wide as the finger along the proximal skin crease at the base of the finger, between the metacarpal and proximal phalanx. A second line was drawn downwards from the tip of the finger, where it automatically snapped to the center of the base line. Raters then zoomed in on the tip of the finger for fine adjustments, to ensure that this line matched the tip as closely as possible. Measurements for each digit were averaged between raters, as inter-rater reliability (Cronbach’s alpha) exceeded .99 for each digit. For each hand, 2D:4D was calculated by dividing the averaged length of the index finger by the averaged length of the ring finger.
It is possible that sex differences in 2D:4D may be confounded by overall digit length (Kratochvíl & Flegr, Citation2009). To examine this in the present data, we also kept the raw length variables for each digit for use as covariates.
Stimuli
The sexual stimuli consisted of 3-minute videos, three featuring a female model and three featuring a male model, each of them masturbating in a bedroom. Females were penetrating themselves with toys or their fingers. These stimuli were selected in a previous study in which 200 videos were rated on their sexual appeal by men and women of different sexual orientations (Rieger et al., Citation2015), and the top three female and male videos were used in the present study. Neutral stimuli to assess baseline genital responses were 2-minute clips taken from a nature documentary. Their engaging but nonsexual content facilitated participants’ return to an unaroused level. However, these nature videos were not used for pupil dilation baseline, as their engaging content might elicit dilation for reasons other than sexual arousal. Thus, two 1-minute animations of clouds were used to obtain a pupillary baseline. All videos were edited using MPEG Streamclip and Final Cut Pro to be of similar luminance.
Genital Arousal
For both sexes, genital response was captured using a BIOPAC MP150 data acquisition unit and the AcqKnowledge software. For men, genital arousal was measured as change in penile circumference with a penile strain gauge. Prior to each participant, the gauge was calibrated on a cone at 80 mm and 110 mm. The signal was sampled at 200 Hz, low-pass filtered to 10 Hz and digitized with 16- bit resolution. For women, genital arousal was measured as changes in peak-to-trough vaginal pulse amplitude (VPA) with a vaginal photoplethysmograph. The signal was sampled at 200 Hz, and high-pass filtered at 0.5 Hz with 16-bit resolution. The VPA exhibits both convergent and discriminant validity for the measurement of female sexual response (Suschinsky et al., Citation2009).
Pupil Dilation
This was measured with a SR Research EyeLink 1000 eye tracking unit. A 35 mm lens focused on the participant’s right eye, positioned approximately 60 cm from the participant’s head, and sampling at a rate of 500 Hz. The infrared light emitted by the eye tracker is reflected by the pupil, and the number of pixels reflected were recorded. Because raw pupil area data included “0’s” for missing values, for instance, from blinks or head movements, these values were removed prior to analyses.
Procedure
Participant Session
The University of Essex’s Ethics Committee approved this study (GR1702). All participants were over the age of 18 and provided written informed consent. After giving consent, participants completed a survey on their demographics and sexual orientation, and had their hands photographed or scanned. Participants were then seated in a sealed booth, with dim lighting conditions. Eyes were calibrated by participants fixating on dots outlining the screen. Participants were instructed on how to use the genital probe, and then were left to apply it in privacy. The experimenter was contactable via an intercom throughout the experiment. The signal from both the genital device and the eye tracker were checked before the experiment commenced. Participants were instructed to watch the screen throughout the experiment, regardless of whether they enjoyed the content. Participants first viewed an animation of clouds, followed by alternating sexual and nature videos. These were displayed in a random order, but a sexual video was always followed by a nature video. Following the sixth nature video, a final animation of clouds was displayed. Participants were compensated for their time. The entire procedure took approximately 90 minutes.
Data Treatment
Following previous procedures (Watts et al., Citation2018), genital data and pupil data were averaged across the duration of each stimulus and for each participant. These averages were then standardized within participants, producing a z-score for each participant and stimulus. For genital data, standardized responses to the 5 seconds preceding each sexual stimulus (following the display of a neutral stimulus, and after the participant had returned to baseline) were subtracted from the standardized response to the sexual stimulus. For pupil data, standardized responses to neutral stimuli (the animated clouds) were subtracted from standardized responses to all sexual stimuli. We then computed, for each participant, average responses across all sexual stimuli of a given type (female or male), which reflected their responses to each sex as compared to baseline.
These standardized response scores were used to calculate the two experimental variables. The first was the absolute difference between each participant’s responses to males and females, calculated by deducting one mean from the other, such that zero indicates equal responses to males and females, and deviation from zero means a stronger response to one sex over the other. We expected men to have a large difference in their sexual arousal to one sex and the other. For example, a heterosexual man should show strong arousal to females, and little to males. Conversely, a homosexual man should show strong arousal to males and little to females. In each case, there would be a notable absolute difference in their responses to males or females (but see our below comment on bisexual men). In contrast to heterosexual and homosexual men, heterosexual and homosexual women (and bisexual women) were expected to show smaller absolute differences, because they would respond similarly to both males and females. Thus, regardless of their sexual orientations, we expected men’s absolute differences in their arousal to males or females to be larger than women’s (Rieger et al., Citation2015).
The second variable used in the present study was participants’ responses to their less-arousing sex, or their minimum arousal. To calculate this, we selected the mean response to whichever stimulus category (male or female) each participant had a lower response to. For men (relative to women), we expected their response to their less-arousing sex to be low, because they often respond strongly to one sex and weakly to the other, regardless of their sexual orientation. For women, more than men, we expected their response to their less-arousing sex to be higher, because women of all sexual orientations are more likely to respond to both sexes, including their less-arousing sex (Rieger et al., Citation2015).
There was the possibility of an interaction between sex and sexual orientation affecting sexual response, because bisexual men’s responses can be more bisexual than the responses of heterosexual or homosexual men, and the responses of homosexual women can be less bisexual than that of other women (Rieger et al., Citation2016; Slettevold et al., Citation2019). In fact, such patterns were found in the present data. However, the present research had no main focus on sexual orientation, but rather on sex differences. Therefore, for the sake of simplicity, we did not concentrate on complex analyses that differentiated bisexual from heterosexual and homosexual men and women, and we decided to investigate sex differences across all participants, regardless of their sexual orientations. Moreover, the inclusion or exclusion of bisexual and homosexual individuals did not change the direction of main findings with respect to sex differences in sexual arousal, although, overall, effect sizes (d’s) increased if only heterosexual men and women were compared, with the largest change being d= 1.96 to d= 2.69 for genital arousal to the less-arousing sex, and the smallest change being d= −.31 to d = −.75 for absolute difference in pupil dilation. However, the exclusion of homosexual and bisexual individuals left the sex difference in 2D:4D only marginally significant in the left hand (p= .06), although the right hand remained significant (p= .03), and the sex differences slightly increased, from .30 to .31, and from .30 to .36, respectively.
Results
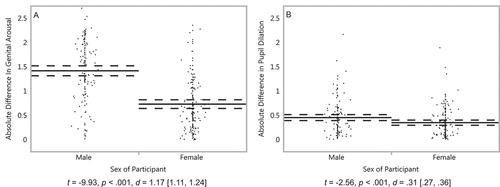
Hypothesis 1. We hypothesized that men’s sexual responses would be more sex-specific, on average, whereas women’s responses would be less sex-specific. We first examined the absolute difference in sexual arousal (measured via genital arousal and pupil dilation) to one sex or the other, expecting that it would be greater in men than in women. For genital arousal, an independent-samples t-test indicated that the absolute difference score was significantly greater in men (M= 1.41, SD = .63) than in women (M= .73, SD = .55), t(292) = −9.93, p< .0001, d= 1.17, 95% CI [1.11, 1.24] (). Similarly, for pupil dilation, the absolute difference score in men (M= .45, SD = .37) was significantly greater than the absolute difference score in women (M= .35, SD = .30), t(271) = −2.56, p= .01, d = .31 [.27, .36] ().
Figure 1. Genital arousal and pupil dilation of men and women to sexual stimuli. Absolute difference between stimuli featuring males and stimuli featuring females in (a) genital responses of 126 men and 168 women and (b) pupil dilation of 118 men and 155 women. On the Y axis, scores reflect the absolute difference between sexual arousal to males and females, standardized within participants. Solid lines represent group means, and dashed lines their 95% confidence intervals. Dots represent participants’ average scores. Captions are independent-samples t-tests, with effect sizes and their 95% confidence intervals.
We then repeated these analyses for sexual response to the less-arousing sex, expecting it to be lower in men than in women. For genital arousal, men’s responses to the less-arousing sex (M= .24, SD = .41) were significantly lower than women’s responses to the less-arousing sex (M = 1.28, SD = .61), t(292) = 16.60, p < .0001, d= −1.96 [−2.02, −1.90] ().
Figure 2. Genital arousal and pupil dilation of men and women to sexual stimuli. Minimum arousal to stimuli featuring males and stimuli featuring females – whichever is lowest – in (a) genital responses of 126 men and 168 women and (b) pupil dilation of 118 men and 155 women. On the Y axis, scores reflect minimum arousal values, standardized within participants. Solid lines represent group means, and dashed lines their 95% confidence intervals. Dots represent participants’ average scores. Captions are independent-samples t-tests, with effect sizes and their 95% confidence intervals.
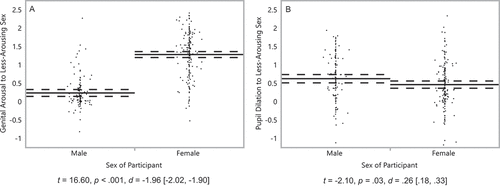
For pupil dilation, men’s responses to the less-arousing sex were not, as hypothesized, significantly lower, but rather greater (M= .62, SD = .64) than women’s (M= .46, SD = .62), and this sex difference was significant, t(271) = −2.10, p = .04, d= .26 [.18, .33] (). As men and women differed as expected in their absolute difference in pupil dilation (), but men unexpectedly showed stronger dilation than women to their less-arousing sex (), we reasoned that this must be driven by an unpredicted difference between men and women in their pupil dilation to their more-arousing sex, whichever sex this may be. That is, we did not hypothesize any difference between men and women in their responses to the more-arousing sex, but investigated this possibility as a potential explanation for the unexpected findings thus far. In fact, men’s pupil dilation to their more-arousing sex (M = 1.08, SD = .60) was significantly greater than women’s (M = .81, SD = .67) t(271) = −3.41, p = .0007, d= .42 [.34, .50]. We revisit this finding in the Discussion.
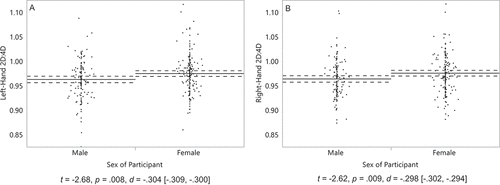
Hypothesis 2. We hypothesized that, on average, men will have a lower 2D:4D than women. In their left hand, men (M = .96, SD = .04) had significantly lower 2D:4D than women, (M = .98, SD = .04) t(316) = −2.68, p= .008, d= −.304 [−.309, −.300] (). Similarly, in their right hand, men (M = .96, SD = .04) had significantly lower 2D:4D than women, (M = .98, SD = .04) t(315) = −2.62, p= .009, d= −.298 [−.302, −.294] ().
Figure 3. Left-hand 2D:4D (a) of 139 men and 179 women, and right-hand 2D:4D (b) of 139 men and 178 women. On the Y axis, 2D:4D is the length of the index finger divided by the length of the ring finger. Solid lines represent group means, and dashed lines their 95% confidence intervals. Dots represent participants’ scores. Captions are independent-samples t-tests, with effect sizes and their 95% confidence intervals.
We further examined whether overall digit length was a confounding variable in the relationship between sex and 2D:4D. To do this, we computed a series of regression analyses predicting either left-hand or right-hand 2D:4D by participant sex and a single digit length variable from the same hand (as digit lengths were highly correlated with each other, r >.98, controlling for both digit lengths simultaneously produced collinearity issues). Sex differences in 2D:4D remained significant, and similar in magnitude, regardless of whether digit length as a potential confound variable was included in the regression analysis or not. For instance, the sex difference in left-hand 2D:4D was significant and identical in effect before controlling for left-hand fourth digit length, p= .008, β = .15 [.04, .26], and afterward, p= .008, β = .14 [.04, .26]. Thus, digit length did not appear to be a confounding factor for observed sex differences in digit ratios.
Hypothesis 3. We hypothesized that sex differences in the specificity of sexual arousal would be associated with a sex difference in 2D:4D. To investigate this, we computed a total of 12 regression analyses predicting absolute difference or response to the less-arousing sex for both genital arousal and pupil dilation. In each analysis, in Step 1, sex was the only predictor of sexual response. In Step 2, either left-hand 2D:4D () or right-hand 2D:4D () were included alongside sex as a predictor. If 2D:4D explained differences between men and women in their sexual response, then the inclusion of either 2D:4D variable should weaken the relationship between sex and sexual response.
Table 1. Multiple regression analyses for sex and left hand 2D:4D predicting absolute difference in genital arousal and response to the less-arousing sex (N = 294) and pupil dilation (N = 273)
Table 2. Multiple regression analyses for sex and right hand 2D:4D predicting absolute difference in genital arousal and response to the less-arousing sex (N = 293) and pupil dilation (N = 273)
Both show that the inclusion of either 2D:4D variable had almost no effect on the relationship between sex and any of the four measures of sexual response. In all Step 2 analyses, the relationship between sex and measure of sexual response remained as strong in effect, and as significant, as in Step 1. However, to conduct further association analysis of 2D:4D variables, we followed these regression analyses with association analyses on the basis of 10,000 bootstrapped samples (Preacher & Hayes, Citation2008). Neither measure of 2D:4D significantly reduced the relationship between sex and any of the four measures of sexual response, as the confidence intervals of the indirect effects included zero in all eight computed analyses. Betas of indirect effects ranged from −.002 to .030, and their CI’s ranged from −.041 to .073.
Discussion
The present research confirmed, in general, two sex differences that have been previously reported in separate research programs: men and women differ in the specificity of their sexual response patterns (Chivers et al., Citation2004; Rieger et al., Citation2015, Citation2016; Suschinsky et al., Citation2009), and men have lower 2D:4D than women in both hands (Grimbos et al., Citation2010). Yet, in the present data, there was no evidence that these two patterns were related.
If one assumes that 2D:4D is a valid measure of prenatal androgen exposure, then this would suggest that sex differences in sexual arousal patterns and digit ratios develop independently of each other. Indeed, some research suggests that there exist several “critical windows” during prenatal and postnatal development in mammals. During these windows, exposure to a stimulus – in this case testosterone – causes masculinization of a specific aspect of brain, morphology or behavior, but to varying degrees and at different time frames, and thus their level of relatedness with each other may be low (Goy et al., Citation1988; McCarthy et al., Citation2018). In humans, a sex difference in 2D:4D appears in fetuses as young as 9 weeks of gestation (Malas et al., Citation2006), and it is possible that the development of sexual arousal is unaffected by androgens during this period. Instead, sexual arousal may be masculinized by androgen exposure separately, during a different critical window of development, meaning that the two are statistically unrelated.
We further stress that the effectiveness of 2D:4D as a measure of prenatal androgenization in general has previously been called into question. However, a large part of this debate is specifically with regards to using 2D:4D as a predictor of sexual orientation, whereas the sex difference is generally considered to be highly robust and reliable (Bailey et al., Citation2016; Swift-Gallant et al., Citation2020). A meta-analysis found no evidence of publication bias in the literature on sex differences in 2D:4D (Grimbos et al., Citation2010). Moreover, our participant numbers substantially exceeded the sample size determined by the power analysis. We therefore believe that if there had been a relationship between sex differences in sexual arousal and in digit ratios, we should have detected it.
Additionally, we note that the sexual response patterns found in pupil dilation did not fully reflect those found in genital arousal, and that for one variable (pupil dilation to the less-arousing sex), the sex difference was in the opposite direction to that predicted (). While genital arousal is a well-evidenced measure of sexual arousal (Janssen, Citation2002; Suschinsky et al., Citation2009), pupil dilation is comparatively newer and has a smaller body of associated evidence. In some studies it has produced results which match with those found through genital arousal (Rieger et al., Citation2015), but in others it has produced results which are not fully identical to genital arousal patterns (Watts et al., Citation2018). Because pupil dilation indicates not only sexual arousal, but also emotion, cognition, or non-sexual interest in stimuli (Bradley et al., Citation2008; Goldinger & Papesh, Citation2012), it is likely that for some participants, pupillary responses were driven by factors other than sexual interest. Despite this limitation, the present unexpected finding that men showed greater pupil dilation to their more-arousing sex than women is still in line with the general observation that men, unlike women, respond more strongly to one sex than the other.
Given the controversy of 2D:4D as a biomarker of prenatal androgen exposure (McCormick & Carré, Citation2020), future research could explore the link between sex differences in sexual arousal and other purported biomarkers of prenatal androgen exposure. Possible options are otoacoustic emissions (McFadden & Pasanen, Citation1998) and anogenital distance (Pasterski et al., Citation2015). Another promising (but involved) method is the direct measurement of androgen levels in the amniotic fluid (amniocentesis), which is a prenatal predictor of gender-typed behavior in later life (Auyeung et al., Citation2009). In theory, participant cohorts who underwent amniocentesis prenatally may be recruited in the future for a study on sexual arousal patterns once they reach adulthood.
In sum, the present study found evidence that men have more specific patterns of sexual response than women, and that men have more male-typical 2D:4D. However, we found no evidence that these two patterns were linked to one another. It may be the case that whichever factor contributes to sex differences in sexual arousal patterns is not reflected in 2D:4D.
Acknowledgment
This research was supported by the Open Access Fund of the Economic and Social Research Council, United Kingdom.
Additional information
Funding
References
- Allaway, H. C., Bloski, T. G., Pierson, R. A., & Lujan, M. E. (2009). Digit ratios (2D:4D) determined by computer-assisted analysis are more reliable than those using physical measurements, photocopies, and printed scans. American Journal of Human Biology, 21(3), 365–370. https://doi.org/https://doi.org/10.1002/ajhb.20892
- Attard-Johnson, J., Bindemann, M., & Ó Ciardha, C. (2016). Pupillary response as an age-specific measure of sexual interest. Archives of Sexual Behavior, 45(4), 855–870. https://doi.org/https://doi.org/10.1007/s10508-015-0681-3
- Auyeung, B., Baron-Cohen, S., Ashwin, E., Knickmeyer, R., Taylor, K., Hackett, G., & Hines, M. (2009). Fetal testosterone predicts sexually differentiated childhood behavior in girls and in boys. Psychological Science, 20(2), 144–148. https://doi.org/https://doi.org/10.1111/j.1467-9280.2009.02279.x
- Bailey, J. M. (2009). What is sexual orientation and do women have one? In D. A. Hope (Ed.), Contemporary perspectives on lesbian, gay, and bisexual identities (Vol. 54, pp. 43–63). Springer. https://doi.org/https://doi.org/10.1007/978-0-387-09556-1_3
- Bailey, J. M., Vasey, P. L., Diamond, L. M., Breedlove, S. M., Vilain, E., & Epprecht, M. (2016). Sexual orientation, controversy, and science. Psychological Science in the Public Interest, 17(2), 45–101. https://doi.org/https://doi.org/10.1177/1529100616637616
- Baumeister, R. F. (2000). Gender differences in erotic plasticity: The female sex drive as socially flexible and responsive. Psychological Bulletin, 126(3), 347–374. https://doi.org/https://doi.org/10.1037/0033-2909.126.3.347
- Berenbaum, S. A., Bryk, K. K., Nowak, N., Quigley, C. A., & Moffat, S. (2009). Fingers as a marker of prenatal androgen exposure. Endocrinology, 150(11), 5119–5124. https://doi.org/https://doi.org/10.1210/en.2009-0774
- Bossio, J., Suschinsky, K., Puts, D., & Chivers, M. (2014). Does menstrual cycle phase influence the gender specificity of heterosexual women’s genital and subjective sexual arousal? Archives of Sexual Behavior, 43(5), 941–952. https://doi.org/https://doi.org/10.1007/s10508-013-0233-7
- Bradley, M. M., Miccoli, L., Escrig, M. A., & Lang, P. J. (2008). The pupil as a measure of emotional arousal and autonomic activation. Psychophysiology, 45(4), 602–607. https://doi.org/https://doi.org/10.1111/j.1469-8986.2008.00654.x
- Breedlove, S. M. (2010). Minireview: Organizational hypothesis: Instances of the fingerpost. Endocrinology, 151(9), 4116–4122. https://doi.org/https://doi.org/10.1210/en.2010-0041
- Breedlove, S. M. (2017). Prenatal influences on human sexual orientation: Expectations versus data. Archives of Sexual Behavior, 46(6), 1583–1592. https://doi.org/https://doi.org/10.1007/s10508-016-0904-2
- Brown, W. M., Hines, M., Fane, B. A., & Breedlove, S. M. (2002). Masculinized finger length patterns in human males and females with Congenital Adrenal Hyperplasia. Hormones and Behavior, 42(4), 380–386. https://doi.org/https://doi.org/10.1006/hbeh.2002.1830
- Chivers, M. L. (2017). The specificity of women’s sexual response and its relationship with sexual orientations: A review and ten hypotheses. Archives of Sexual Behavior, 46(5), 1161–1179. https://doi.org/https://doi.org/10.1007/s10508-016-0897-x
- Chivers, M. L., Rieger, G., Latty, E., & Bailey, J. M. (2004). A sex difference in the specificity of sexual arousal. Psychological Science, 15(11), 736–744. https://doi.org/https://doi.org/10.1111/j.0956-7976.2004.00750.x
- Chivers, M. L., Seto, M. C., & Blanchard, R. (2007). Gender and sexual orientation differences in sexual response to sexual activities versus gender of actors in sexual films. Journal of Personality and Social Psychology, 93(6), 1108–1121. https://doi.org/https://doi.org/10.1037/0022-3514.93.6.1108
- Diamond, L. M. (2003). What does sexual orientation orient? A biobehavioral model distinguishing romantic love and sexual desire. Psychological Review, 110(1), 173–192. https://doi.org/https://doi.org/10.1037/0033-295X.110.1.173
- Ebsworth, M., & Lalumière, M. L. (2012). Viewing time as a measure of bisexual sexual interest. Archives of Sexual Behavior, 41(1), 161–172. https://doi.org/https://doi.org/10.1007/s10508-012-9923-9
- Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–191. https://doi.org/https://doi.org/10.3758/bf03193146
- Galdikas, B. M. F. (1985). Subadult male orangutan sociality and reproductive behavior at Tanjung Puting [https://doi.org/10.1002/ajp.1350080202]. American Journal of Primatology, 8(2), 87–99. https://doi.org/https://doi.org/10.1002/ajp.1350080202
- Gangestad, S. W., & Simpson, J. A. (2000). The evolution of human mating: Trade-offs and strategic pluralism. Behavioral and Brain Sciences, 23(4), 573–587. https://doi.org/https://doi.org/10.1017/S0140525X0000337X
- Goldinger, S. D., & Papesh, M. H. (2012). Pupil dilation reflects the creation and retrieval of memories. Current Directions in Psychological Science, 21(2), 90–95. https://doi.org/https://doi.org/10.1177/0963721412436811
- Goy, R. W., Bercovitch, F. B., & McBrair, M. C. (1988). Behavioral masculinization is independent of genital masculinization in prenatally androgenized female rhesus macaques. Hormones and Behavior, 22(4), 552–571. https://doi.org/https://doi.org/10.1016/0018-506X(88)90058-X
- Grimbos, T., Dawood, K., Burriss, R. P., Zucker, K. J., & Puts, D. A. (2010). Sexual orientation and the second to fourth finger length ratio: A meta-analysis in men and women. Behavioral Neuroscience, 124(2), 278–287. https://doi.org/https://doi.org/10.1037/a0018764
- Hamann, S., Stevens, J., Vick, J. H., Bryk, K., Quigley, C. A., Berenbaum, S. A., & Wallen, K. (2014). Brain responses to sexual images in 46,XY women with complete androgen insensitivity syndrome are female-typical. Hormones and Behavior, 66(5), 724–730. https://doi.org/https://doi.org/10.1016/j.yhbeh.2014.09.013
- Hines, M., Ahmed, S. F., & Hughes, I. A. (2003). Psychological outcomes and gender-related development in complete androgen insensitivity syndrome. Archives of Sexual Behavior, 32(2), 93–101. https://doi.org/https://doi.org/10.1023/A:1022492106974
- Hönekopp, J., Bartholdt, L., Beier, L., & Liebert, A. (2007). Second to fourth digit length ratio (2D: 4D) and adult sex hormone levels: New data and a meta-analytic review. Psychoneuroendocrinology, 32(4), 313–321. https://doi.org/https://doi.org/10.1016/j.psyneuen.2007.01.007
- Israel, E., & Strassberg, D. (2009). Viewing time as an objective measure of sexual interest in heterosexual men and women. Archives of Sexual Behavior, 38(4), 551–558. https://doi.org/https://doi.org/10.1007/s10508-007-9246-4
- Jabbour, J., Holmes, L., Sylva, D., Hsu, K. J., Semon, T. L., Rosenthal, A. M., Safron, A., Slettevold, E., Watts-Overall, T. M., Savin-Williams, R. C., Sylla, J., Rieger, G., & Bailey, J. M. (2020). Robust evidence for bisexual orientation among men. Proceedings of the National Academy of Sciences, 117(31), 18369–18377. https://doi.org/https://doi.org/10.1073/pnas.2003631117
- Janssen, E. (2002). Psychophysiological measurement of sexual arousal. In M. W. Wiederman & B. E. Whitley Jr (Eds.), Handbook for conducting research on human sexuality (pp. 139–171). L. Erlbaum.
- Kinsey, A. C., Pomeroy, W. B., Martin, C. E., & Gebhard, P. H. (1953). Sexual behavior in the human female. Saunders. https://doi.org/https://doi.org/10.1001/jama.1954.02940460077030
- Kratochvíl, L., & Flegr, J. (2009). Differences in the 2nd to 4th digit length ratio in humans reflect shifts along the common allometric line. Biology Letters, 5(5), 643–646. https://doi.org/https://doi.org/10.1098/rsbl.2009.0346
- Kurki, H. K. (2011). Pelvic dimorphism in relation to body size and body size dimorphism in humans. Journal of Human Evolution, 61(6), 631–643. https://doi.org/https://doi.org/10.1016/j.jhevol.2011.07.006
- Lalumière, M. L., Sawatsky, M. L., Dawson, S. J., & Suschinsky, K. D. (2020). The empirical status of the preparation hypothesis: Explicating women’s genital responses to sexual stimuli in the laboratory. Archives of Sexual Behavior, 49(1), 1–20. https://doi.org/https://doi.org/10.1007/s10508-019-01599-5
- Malas, M. A., Dogan, S., Evcil, E. H., & Desdicioglu, K. (2006). Fetal development of the hand, digits and digit ratio (2D:4D). Early Human Development, 82(7), 469–475. https://doi.org/https://doi.org/10.1016/j.earlhumdev.2005.12.002
- McCarthy, M. M., Herold, K., & Stockman, S. L. (2018). Fast, furious and enduring: Sensitive versus critical periods in sexual differentiation of the brain. Physiology & Behavior, 187, 13–19. https://doi.org/https://doi.org/10.1016/j.physbeh.2017.10.030
- McCormick, C. M., & Carré, J. M. (2020). Facing off with the phalangeal phenomenon and editorial policies: A commentary on Swift-Gallant, Johnson, Di Rita and Breedlove (2020). Hormones and Behavior, 120, 104710. https://doi.org/https://doi.org/10.1016/j.yhbeh.2020.104710
- McFadden, D., & Pasanen, E. G. (1998). Comparison of the auditory systems of heterosexuals and homosexuals: Click-evoked otoacoustic emissions. Proceedings of the National Academy of Sciences, 95(5), 2709–2713. https://doi.org/https://doi.org/10.1073/pnas.95.5.2709
- Motta-Mena, N. V., & Puts, D. A. (2017). Endocrinology of human female sexuality, mating, and reproductive behavior. Hormones and Behavior, 91, 19–35. https://doi.org/https://doi.org/10.1016/j.yhbeh.2016.11.012
- Palmer, C. (1989). Is rape a cultural universal? A re-examination of the ethnographic data. Ethnology, 28(1), 1–16. https://doi.org/https://doi.org/10.2307/3773639
- Pasterski, V., Acerini, C. L., Dunger, D. B., Ong, K. K., Hughes, I. A., Thankamony, A., & Hines, M. (2015). Postnatal penile growth concurrent with mini-puberty predicts later sex-typed play behavior: Evidence for neurobehavioral effects of the postnatal androgen surge in typically developing boys. Hormones and Behavior, 69, 98–105. https://doi.org/https://doi.org/10.1016/j.yhbeh.2015.01.002
- Preacher, K. J., & Hayes, A. F. (2008). Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behavior Research Methods, 40(3), 879–891. https://doi.org/https://doi.org/10.3758/BRM.40.3.879
- Puts, D., & Motta-Mena, N. V. (2018). Is human brain masculinization estrogen receptor-mediated? Reply to Luoto and Rantala. Hormones and Behavior, 97, 3–4. https://doi.org/https://doi.org/10.1016/j.yhbeh.2017.07.018
- Rieger, G., Cash, B. M., Merrill, S. M., Jones-Rounds, J., Dharmavaram, S. M., & Savin-Williams, R. C. (2015). Sexual arousal: The correspondence of eyes and genitals. Biological Psychology, 104, 56–64. https://doi.org/https://doi.org/10.1016/j.biopsycho.2014.11.009
- Rieger, G., & Savin-Williams, R. C. (2012). The eyes have it: Sex and sexual orientation differences in pupil dilation patterns. PLoS One, 7(8), e40256. https://doi.org/https://doi.org/10.1371/journal.pone.0040256
- Rieger, G., Savin-Williams, R. C., Chivers, M. L., & Bailey, J. M. (2016). Sexual arousal and masculinity-femininity of women. Journal of Personality and Social Psychology, 111(2), 265–283. https://doi.org/https://doi.org/10.1037/pspp0000077
- Safron, A., Sylva, D., Klimaj, V., Rosenthal, A. M., & Bailey, J. M. (2019). Neural responses to sexual stimuli in heterosexual and homosexual men and women: Men’s responses are more specific. Archives of Sexual Behavior, 49(2), 433–445. https://doi.org/https://doi.org/10.1007/s10508-019-01521-z
- Sawatsky, M. L., Dawson, S. J., & Lalumière, M. L. (2018). Genital lubrication: A cue-specific sexual response? Biological Psychology, 134, 103–113. https://doi.org/https://doi.org/10.1016/j.biopsycho.2018.02.003
- Slaughter, L., Brown, C. R. V., Crowley, S., & Peck, R. (1997). Patterns of genital injury in female sexual assault victims. American Journal of Obstetrics and Gynecology, 176(3), 609–616. https://doi.org/https://doi.org/10.1016/S0002-9378(97)70556-8
- Slettevold, E., Holmes, L., Gruia, D. C., Nyssen, C. P., Watts-Overall, T. M., & Rieger, G. (2019). Bisexual men with bisexual and monosexual genital arousal patterns. Biological Psychology, 148, 107763. https://doi.org/https://doi.org/10.1016/j.biopsycho.2019.107763
- Suschinsky, K. D., & Lalumière, M. L. (2011). Prepared for anything? An investigation of female genital arousal in response to rape cues. Psychological Science, 22(2), 159–165. https://doi.org/https://doi.org/10.1177/0956797610394660
- Suschinsky, K. D., Lalumière, M. L., & Chivers, M. L. (2009). Sex differences in patterns of genital sexual arousal: Measurement artifacts or true phenomena? Archives of Sexual Behavior, 38(4), 559–573. https://doi.org/https://doi.org/10.1007/s10508-008-9339-8
- Swift-Gallant, A., Johnson, B. A., Di Rita, V., & Breedlove, S. M. (2020). Through a glass, darkly: Human digit ratios reflect prenatal androgens, imperfectly. Hormones and Behavior, 120, 104686. https://doi.org/https://doi.org/10.1016/j.yhbeh.2020.104686
- Sylva, D., Safron, A., Rosenthal, A. M., Reber, P. J., Parrish, T. B., & Bailey, J. M. (2013). Neural correlates of sexual arousal in heterosexual and homosexual women and men. Hormones and Behavior, 64(4), 673–684. https://doi.org/https://doi.org/10.1016/j.yhbeh.2013.08.003
- Watts, T. M., Holmes, L., Raines, J., Orbell, S., & Rieger, G. (2018). Sexual arousal patterns of identical twins with discordant sexual orientations. Scientific Reports, 8(1), 1–7. https://doi.org/https://doi.org/10.1038/s41598-018-33188-2
- Wisniewski, A. B., Migeon, C. J., Meyer-Bahlburg, H. F., Gearhart, J. P., Berkovitz, G. D., Brown, T. R., & Money, J. (2000). Complete androgen insensitivity syndrome: Long-term medical, surgical, and psychosexual outcome. Journal of Clinical Endocrinology & Metabolism, 85(8), 2664–2669. https://doi.org/https://doi.org/10.1210/jcem.85.8.6742
- Xu, Y., & Zheng, Y. (2015). The digit ratio (2D: 4D) in China: A meta‐analysis. American Journal of Human Biology, 27(3), 304–309. https://doi.org/https://doi.org/10.1002/ajhb.22639
