Abstract
The effect of liveweight at the start of the breeding period and liveweight gain of hoggets during the breeding period and in pregnancy on breeding performance, pregnancy loss and liveweight and the performance of their progeny were examined. Two groups of hoggets with an average liveweight of 36 kg (denoted L) and 42 kg (denoted H) at the start of the breeding period were allocated to either medium or ad lib levels of herbage during pregnancy, resulting in four treatment groups (L-med, L-ad lib, H-med and H-ad lib). Liveweight changes of the medium and ad lib treatment hoggets were 153 and 208 g/day, respectively. Starting liveweight had no effect (P>0.05) on the overall breeding performance, pregnancy rate, foetuses per hogget joined, number of hoggets lambing or pregnancy loss. Lambs born to the H treatment hoggets were heavier (P<0.05) at weaning than those born to L hoggets (21.69 and 20.11 kg, respectively). More (P<0.05) medium treatment hoggets were not bred during the 22 day breeding period than ad lib treatment hoggets (14 and 4%, respectively). The results indicate that heavy hoggets at the commencement of the breeding period increase the progeny liveweight at the time of weaning and increase hogget weaning weight, but with no effect on reproductive performance. On the other hand, ensuring hoggets are increasing in weight by more than 200 g/day during breeding and pregnancy results in fewer hoggets not bred and hoggets heavier at the time of weaning. Farmers should therefore ensure that adequate herbage is available so that hoggets can achieve these targets.
Introduction
Under New Zealand (NZ) pastoral systems, the reproductive performance of hoggets (age 7–9 months) can be varied and disappointing (Kenyon et al. Citation2004a). In 2006, only 31% of hoggets wintered were bred with an average lambing percentage of 57%, while adult ewe lambing percentage averaged 121% (MWNZ Citation2007). These results indicate that there is scope to lift the lambing percentage of hoggets considerably. Increased liveweight at the time of breeding is positively associated with the proportion of two-tooths and mature ewes displaying oestrus (Coop Citation1962; Adalsteinsson Citation1979; Knight Citation1980; Kenyon et al. Citation2004c) and increases lambing percentage (Kenyon et al. Citation2004b). In hoggets, it has similarly been reported that increasing liveweight at the time of breeding increases the proportion of hoggets cycling (Meyer & French Citation1979; Craig Citation1982), displaying oestrus (Allison et al. Citation1975; Kilgour Citation1977, Moore et al. Citation1978; Moore & Smeaton Citation1980), conceiving (Kenyon et al. Citation2005,Citation2006) and hogget lambing percentage (Kenyon et al. Citation2004b).
Hoggets that have reared a lamb tend to be lighter at the time of breeding as a two-tooth than hoggets not bred (Baker et al. Citation1981; McMillan & McDonald Citation1983; Kenyon et al. Citation2008a) and this can reduce two-tooth reproductive performance (Johnston et al. Citation1982; Kenyon et al. Citation2008a). Relatively high levels of nutrition during pregnancy and in lactation of hoggets are a potential means of achieving adequate two-tooth breeding liveweight.
Under housed conditions in the United Kingdom (UK) and utilising complete concentrate diets, Wallace et al. (Citation1996,Citation1997a,Citationb) and Wallace (Citation2000) have consistently demonstrated that young (5–7 months of age) and well grown (43.7–47.4 kg) hoggets fed to achieve a high liveweight gain (234–301 g/day) during pregnancy displayed increased pregnancy loss and reduced lamb birth weight and survival rates. Under NZ pastoral conditions, pregnancy loss associated with relatively high liveweight gains (210–230 g/day) in pregnancy have only been reported once (Mulvaney et al. Citation2008), with a number of abortions (35%) occurring from day 50 to day 130. However there was no effect on lamb birth weight or survival. In contrast, further studies (Morris et al. Citation2005; Kenyon et al. Citation2008b) of ad libitum pasture-fed hoggets achieving liveweight gains of 210–230 g/day failed to repeat either the UK reports of a detrimental effect of high levels of feeding during pregnancy in hoggets or the results of Mulvaney et al. (Citation2008). It is possible that the abortions reported by Mulvaney et al. (Citation2008) were associated with Neospora caninum infection (West et al. Citation2006; Howe et al. Citation2008), although this is unknown. A clear difference between the UK housed studies and the NZ pastoral studies is hogget liveweight at the start of breeding: 44–47 kg in the UK studies and 36–38 kg in the NZ studies (Wallace et al. Citation1996, Citation1997a,Citationb; Wallace Citation2000; Morris et al. Citation2005; Kenyon et al. Citation2008b; Mulvaney et al. Citation2008).
The aim of the present experiment was to investigate the effect of liveweight at the time of breeding and liveweight gain during pregnancy on the reproductive performance of hoggets, their subsequent liveweight at weaning, and the liveweight and survival to weaning of their progeny.
Materials and methods
Experimental design
The experiment was conducted from May to December in 2007 at Massey University's Keeble farm (latitude 41°10'S), 5 km south of Palmerston North, NZ. The experiment was a 2×2 factorial experiment, incorporating two liveweight groups (light (L) 36.2±0.3 kg, n = 101, range 33.0–39.0 kg and heavy (H) 42.6±0.3 kg, n=102, range 40.0–48.5 kg) and two liveweight gain groups during pregnancy (medium, n=105 and unrestricted liveweight gain (ad lib) n=98) commencing at ram joining (P1) until P139. The aim of the medium nutritional treatment was to achieve a liveweight gain of 100 g/day from P1 to P139. This was to be achieved by maintaining herbage pre- and post-grazing mass between 1000 and 1400 kg DM/ha, respectively. The ad lib treatment hoggets were offered ad libitum levels of pasture (pre-grazing mass >1800 and a post-grazing mass of 1200 kg DM/ha). The hoggets grazed a total area of 16 ha of mixed perennial ryegrass (Lolium perenne) and white clover (Trifolium repens) swards. The size of the grazing area and the grazing interval for both nutritional treatments were determined by the previous weeks’ liveweight gain with the grazing interval adjusted to ensure liveweight targets were met. Herbage mass was monitored using a rising plate meter (Ashgrove Pastoral Products, Palmerston North, NZ) to allocate the interval spent on each grazing area.
Animals
In total, 203 Romney ewe hoggets (7–9 months of age) were used in the experiment. All hoggets had controlled internal drug release dispensers (CIDRs) (progesterone; Pharmacia & Upjohn, NZ) inserted for 12 days. At CIDR removal (P1), L and H liveweight groups were randomly allocated to either medium or ad lib liveweight gain treatments, resulting in four groups within the experiment (L-med, L-ad lib, H-med and H-ad lib). During the experimental period, L-med and H-med were managed as one group and L-ad lib and H-ad lib as another.
At P1, 10 Cheviot, crayon-harnessed, mature rams (ram to ewe hogget ratio of 1:10) were introduced into each liveweight gain treatment group (n=20 in total) for a 5 day period. Crayon marks on the rumps of ewe hoggets were used as an indication of breeding activity and were recorded daily from P1 to P5. Following the 5 day breeding period, the harness crayon colour was changed on the rams, and the rams were re-introduced for a further 17 day breeding period. New hogget crayon rump marks were recorded at the conclusion of this period. Hoggets displaying no rump marks were removed from the remainder of the experiment. At the end of the 22 day breeding period, two harnessed vasectomised rams (n=4 in total) were introduced to each nutritional group for 34 days (P23–P57). Hoggets returning to service between P23 and P57 remained in the experiment until pregnancy diagnosis. At P140, both liveweight gain treatments were combined for lambing and lactation at a stocking rate of 7 hoggets/ha and herbage mass during lactation did not fall below 1800 kg DM/ha.
Animal measurements
Hoggets were weighed un-fasted (within 1 h off pasture) at 7–10 day intervals throughout pregnancy (P1, P7, P13, P20, P26, P34, P41, P48, P55, P62, P74, P88, P102, P117, P139), 35 days after the first lamb was born (L35), L57 and L71 (weaning).
Based on hogget crayon rump marks, the hogget breeding pattern was defined as being bred during the first 5 day period only (P1–P5, synchronised period), bred from P6 to P22 only (P6–P22, post-synchronised period only), bred in both periods (P1–P5 and P6–P22, return to service) or being identified with no crayon rump mark during the 22 day breeding period (not bred at all). Hoggets returning to service post ram removal were recorded on P40 and P57 according to vasectomised rams crayon rump marks. All hoggets that showed evidence of being bred during the 22 day breeding period (P1–P22) had their pregnancy status and number of foetuses (0, 1 or 2) determined via real-time ultrasound on P63. Pregnancy diagnosis was also carried out on P89 and P118. Pregnancy loss was assumed to occur if the hogget was identified as pregnant at P63 but later diagnosed as not pregnant at either P89 or P118. All non-pregnant hoggets at each pregnancy diagnosis were removed from the remainder of the experiment.
Lamb measurements
Lambs were tagged, identified to dam, date of birth recorded, litter size and sex determined, and liveweight recorded within 12 h of birth (L0). In addition, crown rump length (CRL), thoracic girth, forelimb length (FL; distance from the shoulder to the tip of the hoof) and rearlimb length (RL; distance from the hip to the tip of the hoof) were measured. Lambs were re-weighed, un-fasted, at L35, L57 and L71 (weaning). The experiment was conducted with the approval of the Massey University Animal Ethics Committee.
Data analysis
Hogget liveweight and lamb liveweight at birth, L35, L57 and L71 (weaning) and dimensions were analysed using the fixed effects of hogget liveweight group and liveweight gain treatments and their interaction and the sex of the lamb in the model for lamb liveweight using the Generalised Linear Model procedure (SAS Citation2005). Non-significant (P>0.05) interactions were removed and the sum of squares added to the error terms and the model was re-run. The date of birth was used as a covariate for lamb liveweights.
Hogget breeding pattern, percentage of hoggets pregnant per hogget joined at P63, pregnancy loss from P63 to P89 and P89 to P118, number of hoggets lambed per hogget joined and lamb survival from birth to weaning were analysed using the GENMOD procedure for binomial categorical data (SAS Citation2005). Fixed effects of hogget liveweight group and liveweight gain treatments were run for each parameter. Non-significant (P>0.05) interactions were removed and the model was re-run. Foetuses per hogget joined, lambs born and weaned per hogget joined were analysed using the GENMOD procedure for categorical data (SAS Citation2005). The fixed effects of hogget liveweight group and liveweight gain treatment were applied to each parameter. Non-significant interactions (P>0.05) were removed and treatment effects re-analysed.
Results
Hogget liveweight during pregnancy
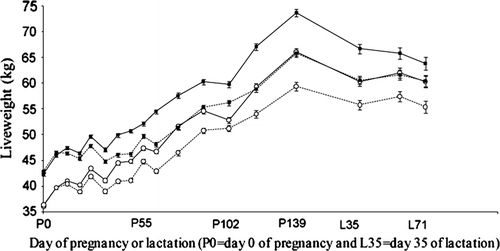
An interaction (P<0.05) between liveweight group and liveweight gain treatment was observed where H-ad lib treated hoggets were heavier (P<0.05) than the L-med, L-ad lib and H-med treatment hoggets from P13 to P148 (). The H-med treatment hoggets were heavier (P<0.05) than the L-ad lib and L-med hoggets from P13 to P74. At P74, P88, P117 and P139, L-ad lib and H-med hoggets were heavier (P<0.05) than L-med treated hoggets. There was no difference (P>0.05) between L-ad lib and H-med treated hoggets from P74 to P139. Liveweight change from P1 to P139 was 216, 151, 194 and 158 g/day for H-ad lib, H-med, L-ad lib and L-med, respectively.
Fig. 1 The effect of hogget liveweight group (L (○) or H (▪)) and nutritional treatment (medium (---) or ad lib (—)) on hogget liveweight during pregnancy (day 0 of pregnancy = P0) and lactation (day 35 of lactation = L35) (mean±SE).
Hogget liveweight during lactation
From L35 to weaning, H-ad lib treatment hoggets were heavier (P<0.05) than L-ad lib, L-med and H-med treatment hoggets, while L-ad lib and H-med hoggets were heavier (P<0.05) than L-med hoggets from L35 to weaning. There was no difference (P>0.05) between L-ad lib and H-med from L35 to weaning.
Hogget breeding performance
Liveweight group had no effect (P>0.05) on the proportion of hoggets bred in the synchronised period only (P1–P5), those returned to service (bred in both periods P1–P5 and P6–P22) or those not bred at all (). However, more (P<0.05) L hoggets were bred when compared with H hoggets in the post-synchronised period only (P6–P22).
Table 1 The effect of hogget liveweight group (L or H) and hogget nutritional treatment (medium or ad lib) during pregnancy on the proportion of hoggets bred (% of hoggets joined) in the first 5 day synchronised breeding period only (P1–P5), only bred during the 17 day period post-synchronised period (P6–P22), bred in both the synchronised breeding and the 17 day period post-synchronised period (P1–P5 and P6–P22, ‘return to service’) and ‘not bred at all’. Values are logit-transformed (mean±SE) and numbers in parentheses are back-transformed percentages. Within columns and treatment, means with letters in common or no superscript do not differ significantly (P>0.05).
Hogget liveweight gain treatment had no effect (P>0.05) on the proportion of hoggets bred during the synchronised period only (P1–P5). A greater proportion (P<0.05) of medium treatment hoggets were bred during the post-synchronised period only (P6–P22) and not bred at all when compared with the ad lib treatment hoggets. More (P<0.05) of the ad lib treatment hoggets returned to service.
Pregnancy rate, foetuses per hogget joined, number of hoggets lambing and lambs weaned per hogget joined were not affected (P>0.05) by liveweight group or liveweight gain treatment (). The percentage of hoggets bred that returned to service between P23 and P63 was not affected (P>0.05) by liveweight group (17 and 25% for L and H, respectively) or liveweight gain treatment (21 and 21% for medium and ad lib treatments, respectively). Pregnancy loss between P64 and P118 was not affected (P>0.05) by liveweight group (18 versus 10% of those pregnant at P64 that lost pregnancy, for L and H, respectively) or liveweight gain treatment (13 versus 13% of those pregnant at P64 that lost pregnancy, for medium versus ad lib, respectively).
Table 2 The effect of hogget liveweight group (L or H) and hogget nutrition (medium or ad lib) during pregnancy on proportion of hoggets pregnant per hogget joined (Percentage of hoggets pregnant), hoggets lambing per hogget joined (Percentage of hoggets lambing), number of foetuses identified per hogget joined (Foetuses), number of lambs born per hogget joined (Lambs born) and lambs weaned per hogget joined (Lambs weaned). Values for percentage of hoggets pregnant, percentage of hoggets lambing are logit-transformed (mean±SE) and numbers in parentheses are back-transformed percentages. Within columns and treatment, means with letters in common or no superscript do not differ significantly (P>0.05).
Lamb dimensions, liveweight and survival
Liveweight group and liveweight gain treatment had no effect (P>0.05) on lamb liveweight at L0, L35 or L57 (). CRL, girth, FL and RL did not differ (P>0.05) between liveweight group and liveweight gain treatment at L0 (data not shown). At L71, lambs born to L treatment hoggets were lighter (P<0.05) than lambs born to H treatment hoggets. Lamb survival was not influenced (P>0.05) by hogget liveweight group or liveweight gain treatment during pregnancy.
Table 3 The effect of hogget liveweight group (L or H) and hogget nutritional treatment (medium or ad lib) during pregnancy on liveweight at birth (L0) and at L35, L57 and L71 (mean±SE) and the proportion of lambs surviving until L71. Values for lamb survival are logit-transformed (mean±SE) and numbers in parentheses are back-transformed percentages. Means within columns with letters in common or no superscript do not differ significantly (P>0.05).
Discussion
This experiment investigated the effect of hogget liveweight and liveweight gain during the breeding period and pregnancy on breeding performance and liveweight of hoggets and the liveweight and survival of their progeny. The liveweight gain treatments resulted in differences in liveweight gains of approximately 50 g/day between treatments, irrespective of liveweight group. However, while total liveweight gains of the ad lib treatment group were similar to those reported by Morris et al. (Citation2005) (210 g/day), Kenyon et al. (Citation2008b) (225 g/day) and Mulvaney et al. (Citation2008) (210 g/day), they were lower than those achieved in the study of Wallace et al. (Citation1996 ,Citation1997a ,Citationb) (234–294 g/day) using a concentrate ration. It is possible that 200 g/day is near the maximum that pregnant hoggets can achieve under NZ pastoral conditions during the late autumn and winter period.
Hogget liveweight
Increasing liveweight at the start of the breeding period has been shown to increase breeding activity (Moore et al. Citation1978; Meyer & French Citation1979; Craig Citation1982; McMillan & Moore Citation1983), therefore potentially increasing the number of hoggets conceiving (Kenyon et al. Citation2005 ,Citation2006). However, in the present experiment, an average difference in hogget liveweight at the time of breeding of 6 kg did not affect the proportion of hoggets bred. McMillan & Moore (Citation1983) reported that a breeding liveweight of 33 kg was required for at least 90% to be reproductively active and Kenyon et al. (2004b) suggested hoggets should be at least 36 kg to ensure production is not limited. In the present experiment, the minimum individual liveweight was 33 kg and this may explain why there was no liveweight treatment effect on the proportion of total hoggets bred.
In the present experiment, liveweight at the start of breeding did not affect pregnancy loss from P63 to P118 and the H treatment hoggets were 42 kg, which was similar to the liveweight at embryo transfer reported in the studies of Wallace et al. (Citation1996,Citation1997b). This might indicate that the pregnancy losses in the studies of Wallace et al. (Citation1996,Citation1997b) were not due to liveweight at the beginning of the experiment.
In mature ewes, the liveweight of the ewe at the time of breeding is positively associated with the lamb birth weight of singles (Russel et al. Citation1981) and twins (Kenyon et al. Citation2004c). In contrast, in the present experiment, hogget liveweight at the time of breeding did not affect lamb birth weight. Kenyon et al. (Citation2006) reported lamb birth weight of single lambs to increase by only 5 g per kilogram increase in hogget breeding liveweight. The numbers used in the present experiment may not have allowed such a small difference to be detected (i.e. 30 g). In the present experiment, lambs born to the H treatment hoggets were 284 g heavier at weaning for each kilogram increase in liveweight at the time of breeding compared with the lambs born to the L treatment hoggets, which were higher than the 146 g/day reported by Kenyon et al. (Citation2006). This increased lamb liveweight could be due to superior milk production for a longer period of time and improved milk composition, although this is unknown and may require further investigation.
The H treatment hoggets were, on average, 6 kg heavier at the start of the experiment and maintained this advantage throughout the experiment. It is unknown if this difference would have remained to two-tooth breeding and potentially affect two-tooth breeding performance but, if it did, there is potential to improve ovulation rate by 4–5% (Smeaton et al. Citation1979) for each kilogram increase in liveweight at the start of the breeding period, and 2% (Morley et al. Citation1978) and 2.5–3% (Cumming Citation1977) for each kilogram increase in mature ewe liveweight at the time of breeding. To the authors’ knowledge, no previous studies have investigated the carry-over effect of heavier hoggets that reared a lamb on subsequent liveweight at two-tooth breeding and their ovulation rate. Further studies are required to determine the impacts of these potential carry-over effects.
Nutrition during pregnancy
In the present experiment, fewer ad lib treatment hoggets were not bred; this indicates that ad libitum levels of nutrition around breeding are beneficial to hogget breeding activity. Increasing the level of nutrition offered to hoggets has increased the proportion displaying oestrus (Moore & Smeaton Citation1980), which is indicative of increased breeding activity.
However, the higher return to service rates in the ad lib treatment hoggets resulted in a similar percentage of hoggets pregnant compared with the medium treatment. This is not the only NZ pastoral study to report this finding (Kenyon et al. Citation2008b). The hoggets in the present experiment and that of Kenyon et al. (Citation2008b) were progesterone synchronised and the results observed may not be directly applicable to non-synchronised natural breeding circumstances. Increasing the level of nutrition offered to mature ewes immediately post-breeding has previously led to a reduction in embryonic survival and was associated with low peripheral progesterone concentrations (Parr et al. Citation1987; McEvoy et al. Citation1995). However, the only study conducted in NZ pastoral conditions investigating progesterone concentration and nutritional treatments during early hogget pregnancy (Mulvaney Citation2006) reported similar progesterone concentrations between nutritional treatments, although greater return rates in the higher nutritional regimen were also observed. Absolute progesterone concentration may not be the only possibility for reduced embryo survival. The balance between progesterone and growth hormone may also be a factor due to an increase in growth hormone associated with increased nutrition (McFadden et al. Citation1990) and progesterone decreases under conditions of increased nutrition (Parr et al. Citation1987; McEvoy et al. Citation1995). Increasing concentrations of growth hormone (McFadden et al. Citation1990) and progesterone (Parr et al. Citation1987; McEvoy et al. Citation1995) are related to increased embryo survival.
The present study showed that pregnancy rate was not affected by liveweight gain during pregnancy, which supports the findings of two previous NZ studies (Morris et al. Citation2005; Kenyon et al. Citation2008b). This suggests that increased hogget liveweight gain under NZ pastoral grazing conditions does not negatively affect pregnancy rates, as shown by Wallace et al. (Citation1996) under indoor concentrate diet systems. Increased liveweight gain was consistently shown to increase pregnancy loss in the studies of Wallace et al. (Citation1996 ,Citation1997b), however only one NZ study (Mulvaney et al. Citation2008) has reported that greater liveweight gains resulted in increased pregnancy loss. The present study supports two other NZ studies (Morris et al. Citation2005; Kenyon et al. Citation2008b) that were undertaken on the same Massey University farm. During the same time period and on the same farm as that studied by Mulvaney et al. (Citation2008), it has been reported that infectious disease caused pregnancy loss in hoggets (West et al. Citation2006; Howe et al. Citation2008). Overall, the combined results indicate that increased liveweight gain during pregnancy under NZ pastoral grazing conditions does not affect the incidence of pregnancy loss.
The studies of Wallace et al. (Citation1996 ,Citation1997a ,Citationb) showed a reduction in lamb birth weight as a result of elevated liveweight gain. However, NZ studies, including the present study, have consistently shown increased liveweight did not affect lamb birth weight. Potential explanations for this could be the different diet types and the resulting differences in metabolic end products and differences in liveweight gains. Altered nutrient partitioning in favour of maternal liveweight gain could have limited the nutrient supply for conceptus growth and placental development as a result of the very high liveweight gains (296 versus 210–230 g/day) during pregnancy in the UK studies (Wallace Citation2000). In addition, concentrate diets have been found to produce a greater proportion of propionate (a gluconeogenesis precursor) than a pastoral diet (Burke et al. Citation2007). This may have affected nutrient supply to the conceptus and affected placental growth and development differently under pastoral grazing systems. However, this has not been investigated. Other potential differences include embryo transfer and age of the hogget at the start of the breeding period (Wallace et al. Citation1996).
Other NZ studies (Morris et al. Citation2005; Kenyon et al. Citation2008b; Mulvaney et al. Citation2008) and the present study have consistently shown that increased liveweight gain during pregnancy did not have a negative affect on lamb survival. In contrast, Wallace et al. (Citation1996) reported that high levels of nutrition reduced lamb survival. This could be due to similar lamb birth weight between nutritional treatments in the NZ studies. In comparison, the lamb birth weight difference was 1.6 kg (34%) in the study of Wallace et al. (Citation1996).
The ad lib treatment hoggets were 6 kg heavier than the medium treatment hoggets at the conclusion of the present study. There is potential for this difference to have a positive effect on two-tooth breeding performance, although this was not measured.
Conclusions
Heavier hoggets at the time of breeding had no effect on breeding performance, pregnancy loss or lamb survival; however, these hoggets remained heavier and produced heavier lambs at weaning. Increasing liveweight gain during pregnancy decreased the proportion not bred during the 22 day period, but did not affect the percentage of hoggets pregnant, pregnancy loss, lamb survival or lamb weaning weight, but the weaning weight of these hogget dams was heavier.
These results suggest that farmers should aim for hoggets that are at least 42 kg at the time of breeding to maximise their weight and that of their offspring at weaning, and should aim for liveweight gains of 200 g/day to the liveweight of the hogget at weaning. In contrast, within the scope of the liveweight groups and nutritional regimens used in the present study, the data indicate that it is difficult to manipulate ewe hogget reproductive performance.
Acknowledgements
The authors would like to thank Mr Dean Burnham for his technical assistance, Massey University and Meat and Wool New Zealand for financial support for this experiment and C. Alma Baker Trust for personal financial assistance.
Related Research Data
References
- Adalsteinsson , S . 1979 . The independent effects of live weight and body condition on fecundity and productivity of Icelandic ewes . Animal Production , 28 : 13 – 23 .
- Allison , AJ , Kelly , RW and Lewis , JS . 1975 . Preliminary studies on the efficiency of mating ewe hoggets . Proceedings of the New Zealand Society of Animal Production , 35 : 83 – 90 .
- Baker , RL , Clarke , JN and Diprose , GD . 1981 . Effect of mating Romney ewe hoggets—preliminary results . Proceedings of the New Zealand Society of Animal Production , 41 : 198 – 203 .
- Burke , J , Murphy , JJ , O'Donovan , MA , O'Mara , FP , Kavanagh , S and Mulligan , FJ . 2007 . Comparative evaluation of alternative forages to grass silage in the diet of early lactation dairy cows . Journal of Dairy Science , 90 : 908 – 917 .
- Coop , IE . 1962 . Liveweight–productivity relationships in sheep. 1: Liveweight and reproduction . New Zealand Journal of Agricultural Research , 5 : 249 – 264 .
- Craig RL 1982 . Breeding from Romney ewe hoggets in the Waihora group breeding scheme . New Zealand Agricultural Science 16 : 101 104 .
- Cumming , IA . 1977 . Relationships in the sheep of ovulation rate with liveweight, breed, season and plane of nutrition . Australian Journal of Experimental Agriculture and Animal Husbandry , 17 : 234 – 241 .
- Howe , L , West , DM , Collet , MG , Tattersfield , G , Pattison , RS , Pomroy , WE , Kenyon , PR , Morris , ST and Williamson , NB . 2008 . The role of Neospora caninum in three cases of unexplained ewe abortions in the southern North Island of New Zealand . Small Ruminant Research , 75 : 115 – 122 .
- Johnston , MT , Kelly , RW and Webb , RR . 1982 . The reproductive performance of Waihora Romney cross hoggets and two-tooth ewe and commercial Romneys . Proceedings of the New Zealand Society of Animal Production , 42 : 47 – 50 .
- Kenyon , PR , Morris , ST , Perkins , NR and West , DM . 2004a . Hogget mating in New Zealand—a survey . Proceedings of the New Zealand Society of Animal Production , 64 : 217 – 222 .
- Kenyon , PR , Pinchcheck , GL , Perkins , NR , Morris , ST and West , DM . 2004b . Identifying factors which maximise the lambing performance of hoggets: a cross-sectional experiment . New Zealand Veterinary Journal , 52 : 371 – 377 .
- Kenyon , PR , Morel , PCH and West , DM . 2004c . The effect of individual liveweight and condition score of ewes at mating on reproductive and scanning performance . New Zealand Veterinary Journal , 52 : 230 – 235 .
- Kenyon , PR , Morel , PCH , Morris , ST and West , DM . 2005 . The effect of individual live weight and use of teaser rams prior to mating on the reproductive performance of ewe hoggets . New Zealand Veterinary Journal , 53 : 340 – 343 .
- Kenyon , PR , Morel , PCH , West , DM and Morris , ST . 2006 . Effect of liveweight and teasing of ewe hoggets prior to breeding on lambing pattern and weight of singleton lambs . New Zealand Journal of Agricultural Research , 49 : 341 – 347 .
- Kenyon , PR , Proctor , L , Morel , PCH , Morris , ST and West , DM . 2008a . The effect of breeding ewe lambs on subsequent two-year-old ewe performance . Livestock Science , 115 : 206 – 210 .
- Kenyon , PR , Morris , ST , Burnham , DL and West , DM . 2008b . Effect of nutrition during pregnancy on hogget pregnancy outcome and birth weight and liveweight of lambs . New Zealand Journal of Agricultural Research , 51 : 77 – 83 .
- Kilgour R 1977 . Hogget mating under good conditions . Proceedings of the Ruakura Farmers Conference 29 : 97 .
- Knight , TW . 1980 . Effects of diet and liveweight on ovulation rates in Romney ewe . Proceedings of the New Zealand Society of Animal Production , 40 : 38 – 42 .
- McEvoy , TG , Robinson , JJ , Aitken , RP , Findlay , PA , Palmer , RM and Robertson , IS . 1995 . Dietary-induced suppression of preovulatory progesterone concentrations in superovulated ewes impairs the subsequent in-vivo and in-vitro development of their ova . Animal Reproduction Science , 39 : 89 – 107 .
- McFadden TB , Daniel TE , Akers RM 1990 . Effects of plane of nutrition, growth hormone and unsaturated fat on growth hormone, insulin and prolactin receptors in prepubertal lambs . Journal of Animal Science 68 : 3171 3179 .
- McMillan , WH and McDonald , MF . 1983 . Reproduction in ewe lambs and its effect on 2-year old performance . New Zealand Journal of Agricultural Research , 26 : 437 – 442 .
- McMillan , WH and Moore , RW . 1983 . Capitalising on hogget oestrus . Proceedings of the New Zealand Society of Sheep and Beef Cattle Veterinarians , 13 : 47 – 52 .
- Meyer , HH and French , RL . 1979 . Hogget liveweight–oestrous relationship among sheep breeds . Proceedings of the New Zealand Society of Animal Production , 39 : 56 – 62 .
- Moore , RW and Smeaton , DC . 1980 . Effects of different growth paths from 4 to 11 months of age on Romney hogget oestrus and subsequent reproduction . Proceedings of the New Zealand Society of Animal Production , 40 : 27 – 33 .
- Moore , RW , Knight , TW and Whyman , D . 1978 . Influence of hogget oestrus on subsequent ewe fertility . Proceedings of the New Zealand Society of Animal Production , 38 : 90 – 96 .
- Morley , FHM , White , DH , Kenney , PA and Davis , IF . 1978 . Predicting ovulation rate from live weight in ewes . Agricultural Systems , 3 : 27 – 45 .
- Morris , ST , Kenyon , PR and West , DM . 2005 . Effect of hogget nutrition in pregnancy on lamb birth weight and survival to weaning . New Zealand Journal of Agricultural Research , 48 : 165 – 175 .
- Mulvaney FJ 2006 . The effect of nutrition during pregnancy on hogget reproduction . Thesis , Massey University , Palmerston North .
- Mulvaney , FJ , Morris , ST , Kenyon , PR and West , DM . 2008 . Hogget nutrition during pregnancy affects pregnancy outcome . Australian Journal of Experimental Agriculture , 48 : 1085 – 1089 .
- MWNZ 2007 Agricultural Production Census. Meat and Wool New Zealand Statistics Wellington , Meat and Wool New Zealand .
- Parr , RA , Davis , IF , Fairclough , RJ and Miles , MA . 1987 . Overfeeding during early-pregnancy reduces peripheral progesterone concentration and pregnancy rate in sheep . Journal of Reproduction and Fertility , 80 : 317 – 320 .
- Russel , AJF , Foot , JZ , White , IR and Davies , GJ . 1981 . The effect of weight at mating and of nutrition during mid-pregnancy on the birth-weight of lambs from primiparous ewes . Journal of Agricultural Science (Cambridge) , 97 : 723 – 729 .
- SAS 2005 . SAS Institute Inc , Cary, NC .
- Smeaton DC Knight TW , Sumner RMW 1979 . Effect of early weaning on ewe liveweight and ovulation rate at mating . Proceedings of the Ruakura Farmers Conference, Agricultural Research Division . Annual report 1979/80 . 171 171 .
- Wallace , JM . 2000 . Nutrient partitioning during pregnancy: adverse gestational outcome in overnourished adolescent dams . Proceedings of the Nutrition Society , 59 : 107 – 117 .
- Wallace , JM , Aitken , RP and Cheyne , MA . 1996 . Nutrient partitioning and foetal growth in rapidly growing adolescent ewes . Journal of Reproduction and Fertility , 107 : 183 – 190 .
- Wallace , JM , Aitken , RP , Cheyne , MA and Humblot , P . 1997a . Pregnancy-specific protein B and progesterone concentrations in relation to nutritional regimen, placental mass and pregnancy outcome in growing adolescent ewes carrying singleton foetuses . Journal of Reproduction and Fertility , 109 : 53 – 58 .
- Wallace , JM , Da Silva , P , Aitken , RP and Cruickshank , MA . 1997b . Maternal endocrine status in relation to pregnancy outcome in rapidly growing adolescent sheep . Journal of Endocrinology , 155 : 359 – 368 .
- West , DM , Pomroy , WE , Collet , MG , Hill , FI , Ridler , PR , Morris , ST and Pattision , RS . 2006 . A possible role for Neospora caninum in ovine abortion in New Zealand . Small Ruminant Research , 62 : 135 – 138 .