Abstract
This experiment assessed the effect of commonly used pasture species and their establishment technique on endoparasites on herbage in Canterbury, New Zealand. Pasture plots were artificially contaminated with lamb faeces with a known endoparasite egg count. Subsequent treatments included retaining the original pasture, replacing the pasture immediately with new pasture via cultivation or direct-drilling, replacing the pasture immediately with a brassica via cultivation or direct drilling, or replacing the pasture immediately with a barley grain crop via cultivation. Brassica and grain crops were later replaced with new pasture. Herbages were harvested and dry matter (DM) yield and numbers and species of endoparasites (L3) were determined. Pasture and brassica establishment technique had no effect on herbage production. The old pasture initially contained more L3 than all other treatments, but subsequently had a similar number to pasture renewed by direct drilling, both of which had considerably more than all other treatments. Establishing new pasture by direct drilling resulted in 12 760 L3/kg DM over 56 weeks, nearly three times more than occurred following cultivation. Brassica swards had fewer L3 than did new grass. Nematodirus L3 were disproportional to the number of eggs deposited; they represented 2% of the eggs deposited but 67% of the L3, and they were more persistent, representing 30% of the L3 present during March but 87% during October.
Introduction
There is a compelling need to reduce reliance on anthelmintics for gastro-intestinal parasite management in sheep systems. Endoparasites of sheep in New Zealand have traditionally been controlled using drugs but anthelmintic resistance in sheep endoparasites has become increasingly common (Rhodes et al. 2006). Consequently, farmers are now being encouraged to reduce anthelmintic use with the intention of reducing or slowing down the development of parasite resistance. However, this strategy is likely to result in increased parasite contamination of grazing areas.
Clearly, reducing the ingestion of parasite-infected herbage is an obvious means of reducing the impact of endoparasites on animal performance. Achieving this requires knowledge of both the level of parasite infection of the grazable herbage and how to reduce the level of parasite infection. According to Familton & McAnulty (Citation1996), more than 90% of the endoparasites in sheep systems in New Zealand are located within the pasture and soil environment. Factors known to have a major effect on populations within a sward include moisture, humidity and temperature (Anderson et al. Citation1970; Todd et al. Citation1976). The micro-climate within a sward can be affected by sward density (Moss & Bray Citation2006) and pasture species (Knapp Citation1964; Moss & Vlassoff Citation1993).
In a laboratory-generated micro-climate, Ostertagia ostertagi L3 survived from between 2 days to more than 52 weeks depending on the temperature to which they were subjected (Pandey Citation1972). In a pasture-based field experiment (with associated fluctuations in climatic conditions) conducted in Canterbury, New Zealand, Trichostrongylus spp. and Teladorsagia spp. were detected on herbage up to 26 weeks following an autumn contamination, with the peak occurring after 10 weeks (Moss & Bray Citation2006).
Prudent farm management would ensure that the most vulnerable stock would graze the least parasitised herbage. The objective of the present experiment was to assist this objective by determining the effect of some commonly used pasture species and their establishment technique on the endoparasite population within the sward over time.
Materials and methods
Site preparation
The experiment was conducted on flat land at Lincoln, near Christchurch, New Zealand. The area used was initially in 12 year old pasture that had been ungrazed for two years. Plots measuring 6×2.5 m with 3 m between them were established on this area. Fresh faeces obtained from parasitised lambs on 10 December 2006 were uniformly applied to a central 1×1 m area of each plot one day later. The application rate equated to 692 064 Nematodirus spp. and 1 640 736 other strongyle eggs per m2. Faeces were applied in individual mounds measuring approximately 40 mm in diameter and placed 110 mm apart.
Experimental design and management
The plot layout was a randomised block design with five replicates and the following six treatments:
| 1. | retained as old pasture (OP) | ||||
| 2. | new pasture via cultivation (NPC) | ||||
| 3. | new pasture via direct drilling (NPD) | ||||
| 4. | brassica via cultivation followed by new pasture (BC-NP) | ||||
| 5. | brassica via direct drilling followed by new pasture (BD-NP) | ||||
| 6. | barley grain crop via cultivation followed by new pasture (GC-NP) | ||||
Table 1 Mean temperatures and rainfall for the periods preceding herbage harvest dates, measured 1 km from the experimental site.
Herbage was harvested between 7.30 and 9.00 a.m. on eight occasions between early February 2007 and January 2008, when the mean herbage mass reached normal grazing height. No parasite assessments were made on the barley crop. All plots within replicates were harvested simultaneously with all replicates harvested over a 10-day period. Herbage was cut at 15 mm above ground level using a hand clipper. The area harvested measured 0.5 m2 in the centre of the area where faeces had been applied. Herbage was also cut from the remainder of the area to which faeces had been applied, and discarded. Nematode larvae were extracted from the herbage using a modified Baermann technique as described by Vlassoff (Citation1973). The herbage was subsequently weighed, sub-sampled for botanical composition determination and dried to determine dry matter (DM) yield.
Statistical analyses
Data were statistically analysed using analysis of variance, with five contrasts included in the analysis. The first contrast compared OP with the average of the five renewal treatments. The next three contrasts were the main effect and interaction contrasts for the 2×2 factorial treatment structure for treatments 2–5, where the factors were cultivation method (cultivation versus direct drilling) and timing of new pasture establishment (‘new pasture directly’ versus ‘new pasture following brassica’). Hence the second contrast was the ‘main effect’ comparison of cultivation versus direct drilling (treatments 2 and 4 versus 3 and 5). The third contrast was the ‘main effect’ comparison of ‘new pasture directly’ versus ‘new pasture following brassica’ (treatments 2 and 3 versus 4 and 5). The fourth contrast was the interaction of the two factors. The fifth contrast was a simple pairwise comparison of barley and brassica, both established by cultivation (treatment 4 versus 6).
To calculate the average number of larvae per kilogramme for a time period spanning several harvests, a weighted average was calculated for each plot by multiplying the number of larvae per kilogramme by the DM yield (in kg/ha) for each harvest, then summing the resulting estimated numbers of larvae per hectare over the relevant harvests, then dividing by the sum over the harvests of the relevant DM yields. This is conceptually equivalent to making a single heap of the material harvested from each plot, pooled over the harvests, and then estimating the number of larvae per kilogramme in the combined heap.
Results
All pastures consisted predominantly of grass, representing approximately 93% of the old grass sward, 75% of the new pasture, 62% of the post-brassica and 81% of the post-barley pastures (). The new and post-brassica pastures consisted of proportionally more clover than the other treatments.
Table 2 Botanical composition of grass- and brassica-based swards following different renewal techniques. OP, old pasture; NPC, new pasture via cultivation; NPD, new pasture via direct-drilling; BC-NP, brassica via cultivation followed by new pasture; BD-NP, brassica via direct drilling followed by new pasture; GC-NP, barley grain crop via cultivation followed by new pasture.
Replacing the OP immediately with either new pasture or brassica followed by new pasture, increased herbage DM production during all periods in which production was measured ().
Table 3 Effect of herbage type and pasture establishment technique on herbage production during different phases of pasture renewal. The first contrast compares old pasture with the average of the five other treatments (2–6). The second contrast is the main effect comparison of cultivation versus direct drilling (treatments 2 and 4 versus 3 and 5). The third contrast is the main effect comparison of new pasture directly versus new pasture following brassica (treatments 2 and 3 versus 4 and 5). In the last column, treatment 6 was omitted from the statistical analysis. Means within columns with superscript letters in common are not significantly different (P<0.05): ns, not significant; * P<0.05; ** P<0.01; ***P<0.001.
Brassica swards produced more DM than existing pasture or barley grain followed by new pasture during the three months in which they were grown. However, pasture established after the brassica produced less than was produced during the same time period by pasture established initially, resulting in less production over the entire experiment from the brassica treatments. Establishment technique had no effect on subsequent herbage production.
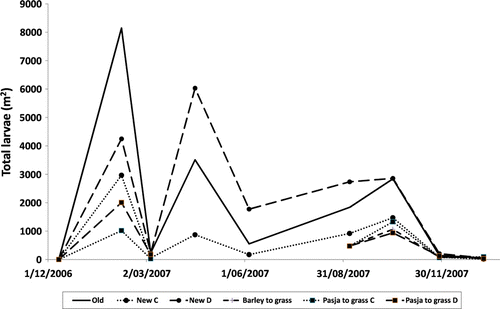
Old pasture yielded more endoparasite larvae/m2 than any other treatment during the 12 weeks following egg deposition (). However, OP and NPD produced similar numbers of larvae/m2 subsequently. This resulted in these two treatments having a similar number of larvae when assessed over the entire 56-week trial period, which was considerably more than the number produced by all other treatments.
Table 4 Effect of herbage type and establishment technique on the number of sward-borne endoparasite larvae (strongyle and Nematodirus). The contrasts reported are as in . In the second column, treatment 6 was omitted from the statistical analysis. In the last column, treatments 4, 5 and 6 were omitted from the statistical analysis.
Establishing new pasture by direct drilling resulted in more larvae/m2 than when cultivation was used, on all dates compared, although the difference was non-significant (P<0.05) during the initial 12 weeks. Overall, larval numbers equated to 18 144/m2 and 6560/m2 from direct drilled and cultivated new grass treatments respectively (). The brassica produced fewer larvae than did new grass, although this difference was only significant (P<0.05) when compared with new grass direct drilled. The post-brassica and post-grain pasture produced a similar number of larvae following their April sowing, which was fewer than occurred on all other treatments over this period.
The old pasture yielded more larvae/kg DM than did all other herbages during the first 12 weeks following egg deposition (). During the following 44 weeks, OP and NPD swards produced similar relatively high larval numbers. Brassica established following cultivation produced the lowest number of larvae during the first 12 weeks, while thereafter NPC, BC-NP, BD-NP and GC-NP yielded similar numbers. NPD contained more larvae/kg DM than NPC overall and on all measurement dates, although the difference during the first 12 weeks was not significant.
Table 5 Effect of herbage type and establishment technique on the number of sward borne endoparasite larvae (strongyle and Nematodirus). The contrasts reported are as in . In the last column, treatments 4, 5 and 6 were omitted from the statistical analysis.
There was considerable variation between sampling dates in the number of larvae extracted from herbage (). Numbers peaked during February, 8 weeks after eggs were deposited, but were extremely low at the next sampling 4 weeks later. Other, albeit, smaller peaks occurred during April (week 18) and September (week 38).
Figure 1 Effect of herbage species and establishment technique on the number of sward borne endoparasite larvae (strongyle and Mematodirus combined L3/m2).
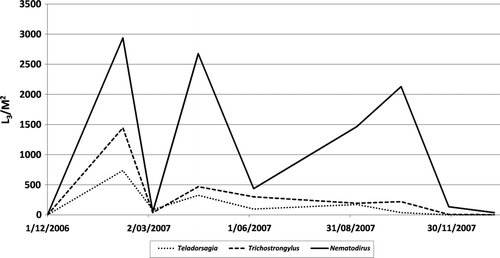
Nematodirus numbers were higher than strongyle on all measurement dates, particularly during autumn (week 18) and spring (week 44) (). While this species of nematode represented only 2% of the faecal worm eggs initially deposited they represented 67% of the larvae on the herbage when assessed over all treatments and dates. Nematodirus were also considerably more persistent. They represented 30% of the larvae present on herbage during March (week 12) but 87% during October (week 44) when assessed over all treatments. Nematodirus L3 when assessed over all treatments and dates, equated to 1.13% of the eggs deposited while the comparable figure for strongyle was 0.21%.
Figure 2 Comparing the number and seasonal variation of the different endoparasite L3 on old pasture, new pasture following cultivation and new pasture direct drilled.
Discussion
Endoparasite larval development and survival is affected by temperature, humidity, moisture (Rees Citation1950; Anderson et al. Citation1970) and soil shading (Furman Citation1944). In Furman's experiment, shaded larvae survived for up to 237 days whereas few survived to 60 days from unshaded soils. These climatic factors would have been affected by the treatments applied in the current experiment and consequently influenced the number of larvae extracted from herbage. Retaining the herbage canopy (as in the old pasture treatment in this experiment) resulted in the presence of more larvae on herbage than occurred on any other herbage type during the initial 12-week period following egg deposition.
Prior to the new pasture attaining ground cover, eggs and larvae on the direct drilled area would have been exposed to the summer weather with little protection compared with that available under the old pasture. Consequentially, there were initially fewer larvae where the original pasture was desiccated by herbicide. However, when assessed over the following 44 weeks, the OP and NPD contained similar numbers of larvae.
The low herbage mass from the old pasture accentuated the ratio of larvae to herbage DM on this pasture. Total pasture produced by the old pasture was lower than achieved by all other treatments, reflecting its high cocksfoot and low legume component.
Compared with pasture established after cultivation, those pastures that were direct drilled had more larvae on all measurement dates—equating to three time more when assessed over the entire experimental period. Cultivation in this experiment involved ploughing within a day of egg-containing faeces being deposited, which would have resulted in their burial at a depth of up to 185 mm. While this would have protected eggs and larvae from the ravages of the summer climate it would also have made it difficult for larvae to relocate to pasture.
Nematodirus were considerably more successful at surviving on the herbage in the conditions of this experiment. While they represented only 2% of the faecal worm eggs initially deposited on these plots, they represented 67% of the larvae on the herbage when assessed overall treatments and dates. Furthermore, Nematodirus persisted for a considerably longer period than did Teladorsagia or Trichostrongylus, with relatively high numbers (0.20 and 0.13% of eggs deposited) on herbage during autumn (week 18) and spring (week 44) respectively, compared with 0.03 and 0.01% for Teladorsagia and Trichostrongylus combined. Clearly, when predicting the likely level of contamination of a sward using the number of eggs deposited as a factor, due consideration needs to be given to the endoparasite species present and climatic conditions.
Larval levels on brassica were consistently lower than occurred on new grass, although differences were only significant when comparing BC-NP and NPD (P<0.05). This herbage type effect may reflect the greater density of the grass-based swards, a factor shown by Knapp (Citation1964), Moss and Vlassoff (Citation1993) and Moss and Bray (Citation2006) to affect larval populations on herbage. However, both the new pasture and pasture following brassica contained considerably more clover (average 23%) than did old pasture (1%) or pasture following barley (4%). Knapp (Citation1964) found clover was more conducive than grass to larval development and survival—an effect considered to reflect a more favourable micro-climate.
It would seem reasonable to assume that the extremely low larval counts in March reflect the exceptionally low rainfall (10 mm ()) and relatively high air temperatures (maximum 32°C (not shown)) recorded during the 26 days prior to this harvest. As mentioned earlier, sufficient moisture is critical to endoparasite development and survival (Anderson et al. Citation1970), hence the considerable variation in numbers between harvest dates in this experiment. Anyone attempting to monitor L3 loadings on pasture to determine endoparasite risk to susceptible grazing animals needs to be aware of the influence of prevailing climatic conditions when sampling herbage.
It should be noted that the number of L3 detected on the last sampling date of this experiment may have been influenced by the lack of rain (not shown) and air temperatures frequently reaching 30°C (not shown) during the 12 days preceding this harvest. Consequently, it cannot be assumed that endoparasites, especially Nematodirus, no longer existed beyond this date.
Acknowledgements
The authors acknowledge the Foundation for Research, Science and Technology (FRST) for funding the research and thank DJ Saville for the statistical design and analyses and A Vlassoff and RW McAnulty for their technical advice.
References
- Anderson , FL , Levine , ND and Boatman , PA . 1970 . Survival of third stage Trichostrongylus colubriformis larvae on pasture . Journal of Parasitology , 56 : 209 – 232 .
- Familton AS , McAnulty RW 1996 . Proceedings of 26th seminar of the Sheep and Beef Cattle Society of the New Zealand Veterinary Association 73 81 .
- Furman , DP . 1944 . Effects of environment upon the free-living stages of Ostertagia circumcincta (Stadelmann). Trichostrongylidae/laboratory experiments . American Journal of Veterinary Research , 5 : 79 – 86 .
- Knapp , SE . 1964 . Relationship of different species of forage to the survival and infectivity of Haemonchus contortus in lambs . Journal of Parasitology , 50 : 144 – 148 .
- Moss , RA and Bray , AR . 2006 . Effect of sward density and faecal deposit size on the development and persistence of third-stage trichostrongylid larvae of sheep . New Zealand Journal of Agricultural Research , 49 : 475 – 481 .
- Moss , RA and Vlassoff , A . 1993 . Effect of herbage species on gastro-intestinal roundworm populations and their distribution . New Zealand Journal of Agricultural Research , 36 : 371 – 375 .
- Pandey , PJ . 1972 . Effect of temperature on survival of the free living stage of Ostertagia ostertagi . Journal of Parasitology , 58 : 1042 – 1046 .
- Rees , FG . 1950 . Observations on the vertical migrations of the third stage larva of Haemonchus contortus (Rud.) on experimental plots of Lolium perenne S24 in relation to meteorological and micrometeorological factors . Parasitology , 40 : 127 – 143 .
- Rhodes , AP , Leathwick , DM , Pomroy , WE , West , R , Jackson , R , Lawrence , K and Waghorn , TS . 2006 . A profile of anthelmintic resistance and parasite control practises in New Zealand–results from a 2005 survey . Proceedings of the New Zealand Society of Animal Production , 66 : 14 – 19 .
- Todd , KS Jr , Levine , ND and Boatman , PA . 1976 . Effect of temperature on survival of free living stages of Haemonchus contortus . American Journal of Veterinary Research , 37 : 991 – 992 .
- Vlassoff , A . 1973 . Seasonal incidence of infective trichostrongyle larvae on pasture grazed by lambs . New Zealand Journal of Experimental Agriculture , 1 : 293 – 301 .