Abstract
Concerns about climate change and water quality make it necessary to have a better understanding of the cycling of carbon (C) and nitrogen (N) within landscapes. In New Zealand, pastoral farming on hill country is a major land use, and there is little information available at a landscape level on the cycling of C and N within these systems, particularly the impacts of land use intensification. Published information on the individual components of C and N cycles in hill country pastoral systems was used to construct simple C and N models for two notional hill country paddocks with contrasting amounts of annual net herbage accumulation (NHA). These simple models of C and N cycling were used to explore the potential impacts of intensification on soil C and N pools. The C and N models constructed illustrate that both the C and N cycles in hill country pastoral farming are characterised by large fluxes in and out of the system and relatively small annual accumulations or depletions of their respective soil pools. The mechanisms by which these inward and outward fluxes are generated differ greatly between C and N. Most of the C cycling through the soil/plant/animal system in a year is sourced from the atmosphere through photosynthesis in that same year and the annual quantity of C ingested by grazing animals is only approximately 25% of that fixed annually. In contrast, the annual quantity of N ingested by grazing animals is 300–400% of that added to the system annually by fixation, and most of this ingested N (>80%) is returned to the soil in animal excreta. This contrasts with <25% of ingested C returned in animal excreta. Grazing animals, with their propensity to exhibit camping behaviour, are therefore much more influential in the spatial redistribution of N than C in hill country pastures. The simple C and N models presented in this paper indicate that while land use intensification results in increased inputs of both C and N to the soil, intensification is also likely to result in increased losses of C and N from the soil pool. Which of these processes dominates in a given situation determines whether intensification will result in an increase or decrease of soil C and N. Based on a simple C model, we postulate that if intensification increases NHA by one unit there is likely to be an increase in soil C unless at the same time increased grazing pressure results in at least a three-unit increase in dry matter eaten/harvested.
Introduction
Two environmental issues of major concern are increasing concentrations of greenhouse gases in the atmosphere and increased concentrations of nitrogen (N) in receiving waters. These are affected by global carbon (C) and N cycles, and soil C and N are major components of these cycles. Hill country farmers are intensifying operations on at least parts of their land, with consequences for the C and N cycles (MacLeod & Moller Citation2006).
Hill country pastoral landscapes occupy a large area of New Zealand, but compared with flat land, relatively little is known of nutrient cycling in these landscapes. The work that has been done has focused mainly on phosphorus (P) and, to a lesser extent, sulphur (S). Although there has been considerable work done on some processes (i.e. rates of net herbage accumulation (NHA) and N fixation, nitrification and denitrification) that are important components of the C and N cycles in hill country, very little experimental work has focused on the whole-system cycling of N and C within hill country landscapes.
This paper briefly reviews existing information of the processes important in the cycling of C and N in hill country and uses this existing knowledge to build simple C and N models for two notional hill country paddocks with contrasting amounts of annual NHA. Critical areas of weakness in existing knowledge are identified and these simple models of C and N cycling are used to explore likely effects of intensification of production on soil C and N in these two notional paddocks. We acknowledge that more detailed and comprehensive models of C and N cycling in other farming systems exist and we do not intend to present an exhaustive review. Rather, the intent of this paper is to present simple models of C and N cycling in grazed hill country to encourage and stimulate thought and discussion by those interested in and working in this unique environment.
System boundaries, definitions and scenarios
In this study, the term ‘hill country’ refers to land that is used for pastoral agriculture and in which the predominant slope is >15°. Within the hill country landscape there are areas of land with slopes <15°, but they usually occupy only a small percentage (<30%) of most hill country paddocks. There are approximately 4.6 million ha of hill country in New Zealand (de Klein et al. Citation2009).
The C and N models for hill country presented here use existing information to create quantitative C and N cycles for two notional 1 ha hill country paddocks with contrasting amounts of annual NHA, representing a summer moist and a summer dry hill country environment. The two notional paddocks have been constructed to maximise the use of published information when creating these nutrient cycles. The two paddocks are nominally divided into flat/camp (0–12°), easy (13–25°) and steep (>25%) slope categories representing 9%, 52% and 39%, respectively, of the total paddock area.
The decision to develop the models specifically for 1 ha paddocks rather than for a more generic hill country landscape was to avoid confusion in the presentation of data. When aggregating these losses or gains on individual slope categories into a landscape model, corrections must be made for the proportion of the landscape occupied by each slope category. In presenting these data, there is potential for confusion between ‘losses or gains per hectare’ within individual slope categories and ‘losses or gains per hectare’ at the landscape scale. We have therefore chosen to refer to our landscape scale model as being a 1 ha paddock to differentiate the resulting losses and gains clearly from those on individual slope categories.
The first step was to establish a ‘base scenario’. The NHA data, herbage N concentrations and legume contents assigned to the ‘summer moist’ paddock () in the base scenario are based on data presented by Ledgard et al. (Citation1987) for a hill country site at Whatwhata in the Waikato. The corresponding data for the ‘summer dry’ paddock are taken from Blennerhassett (Citation2002) for a hill country site near Waipawa in Central Hawke's Bay. The differences in measured annual NHA between these two sites result mainly from differences in annual rainfall (Zhang et al. Citation2005).
Table 1 Base scenario input parameters.
In the base scenarios, for both paddocks, pasture utilisation was assumed to be 60%, 70% and 50% on flat, easy and steep sites, respectively. These values are lower than those measured by Gillingham (Citation1978), but are consistent with observations of Ledgard et al. (Citation1987). The pattern of excretal deposition in the base scenario was based on the work of Gillingham (Citation1978) with 67%, 27% and 4% of all excreta deposited on flat, easy and steep areas, respectively.
Once the base scenario had been established, a number of alternative scenarios were investigated to determine the effect of changing the assigned patterns of pasture utilisation and excretal deposition on the C and N models. These alternative scenarios were devised to investigate the likely effect of changing pasture utilisation and pattern of excretal deposition. In the intensively grazed scenario, utilisation was set at 79%, 83% and 76% for flat, easy and steep slopes, respectively. For the extensively grazed scenario, pasture utilisation was set at 50% for all three slope categories.
The base scenario assumed a high degree of nutrient transfer with only 4% of total excreta being deposited on the steep slopes. A ‘moderate-transfer’ scenario was investigated in which 33% of the total excreta was returned to each of the three slope categories. Because the flat/camp areas only occupy 9% of the total paddock area, there was still significant transfer on a ‘per hectare’ basis to these areas in this scenario. A ‘zero transfer’ scenario was also investigated in which excreta were returned to slopes in direct proportion to their areas (9%, 52% and 33% to flat, easy and steep slopes, respectively).
Finally, a more ‘intensively managed’ scenario was investigated. In this scenario, assumed annual NHA on easy and steep slopes for both sites was increased by 1000 kg DM ha−1 yr−1 through the addition of N fertiliser. At the same time it was assumed that pasture utilisation would be improved from that in the base scenario to correspond to the high-utilisation scenario described above (i.e. 79%, 83% and 76% for flat, easy and steep slopes, respectively). As a consequence of this higher utilisation, it was assumed that the pattern of excretal deposition would be as described in the moderate-transfer scenario described above (i.e. 33% to each slope class). We assumed no change in the N content of the grazed herbage with the addition of fertiliser N (see ).
C cycling in grazed hill country with emphasis on the soil C pool
Background
Pasture plants link atmospheric and soil C pools through the process of photosynthesis (C fixation), rhizodeposition, translocation and respiration. The C concentration of the soil at any time is determined by the balance between C inputs through litter, dung, rhizodeposition and root senescence, and the rate of decomposition of these materials that releases C back to the atmosphere as carbon dioxide (CO2) through respiration at the soil surface (R s). Soil C balance must also take account of losses of C from the soil as dissolved organic C (DOC) through leaching and/or runoff (Ghani et al. Citation2007; Sanderman et al. Citation2008) and particulate organic C (POC) through erosion and/or runoff (Lambert et al. Citation1985; Page et al. Citation2004). Although of great agronomic importance, a grazing animal contributes only a small proportion of photosynthate to the soil C pool in a grazing system (Soussana et al. Citation2004).
A simple C model: model development
A simple C model for grazed hill country, which included features of terrestrial C cycles as presented by Soussana et al. (Citation2004), Sitaula et al. (Citation2004) and Sanderman & Amundson (Citation2008), was constructed. The key processes in the C cycle used to construct the simple C model are briefly described below, with emphasis on information drawn from studies on hill country in New Zealand. At the end of each section the approach taken to derive values for that particular process in the model for the two case study paddocks is outlined. Where information for grazed hill country or even New Zealand grazed pastures was lacking, information was taken from temperate grazing situations outside of New Zealand. All values are presented on an annual basis.
Inputs of C to the terrestrial pool
Photosynthesis
The process of photosynthesis is the key driver of C cycling in a grazing system. The total amount of C fixed or assimilated through photosynthesis (TCA) and the net amount of C accumulated in above ground herbage (NCA) in any grazing system is dependent on climate, slope, soil fertility and grazing management (Zhang et al. Citation2005). In a grazed pasture TCA has been measured/estimated to be approximately four times NCA (Saggar et al. Citation1999; Soussana et al. Citation2004).
For our simple C model, TCA was calculated as being NCA/0.26. In order to calculate NCA we assumed the organic matter content of NHA to be 90% (i.e. we assumed an average ash content of herbage of 10% (Frame Citation1992)), and that C comprises approximately 40% of herbage organic matter (Parsons & Chapman Citation2000).
Example calculation for summer moist flat site: if NHA=14,310 kg DM ha−1, then NCA=14,310×0.90×0.40=5152 kg C ha−1 and TCA=5152/0.26=19,814 kg C ha−1.
Rhizodeposition and root turnover
The flux of C from living roots into the soil (rhizodeposition) results in the close coupling of plant roots and the soil microbial activities of free living and symbiotic micro-organisms. Rhizodeposition comprises exudates (sugars, amino acids and organic acids), secretions (including enzymes and mucilages) and dead cells sloughed from root caps and senescing roots. It also includes the C flux that is transported out of roots through the symbiotic mycorrhizal fungi that are associated with over 80% of plants. In grasslands, the root-infecting mycelia of these fungi receive approximately 10% or more of net plant photosynthate and transport this out of roots through their external mycelial networks that proliferate extensively through the soil, absorbing nutrients (Leake et al. Citation2006).
In our simple C model we assumed that 37% of the TCA would be allocated to rhizodeposition and root turnover (Saggar et al. Citation1997; Soussana et al. Citation2004).
Example calculation for summer moist flat site: if TCA=19,814 kg C ha−1, then rhizodeposition and root turnover=19,814 kg C ha−1×0.37=7331 kg C ha−1.
Translocations of fixed C
Return of C in plant litter
In a grazing system, NCA is either harvested by the grazing animal or is returned to the soil surface as litter. As grazing systems undergo intensification, a greater proportion of NCA is ingested and a smaller proportion of NCA is returned to the soil as litter. Soil fauna play a particularly important role in the incorporation of litter C into the soil C pool (Parfitt et al. Citation2005; Mackay Citation2008). Estimates of the amount of NCA returned to the soil C pool as litter are difficult as a proportion of the C in litter is released back into the atmosphere as it undergoes senescence and before it can be incorporated into the soil C pool. There are several schools of thought that variably estimate the amount of litter returned to the soil pool; these range from litter being a minor contributor to the soil C pool to a near complete incorporation of the C in litter into the soil C pool (Parsons et al. Citation2009). Photodegradation of plant litter, a process by which solar radiation directly breaks down the compounds of organic matter in litter and thus increasing CO2 fluxes, has been shown to be of importance in warm, dry environments and on recently mined peat soils in New Zealand (Rutledge et al. Citation2010). There is no published information on the impact of photodegradation of plant litter on returns of litter C to the soil for flat land or hill country pastures in New Zealand (Rutledge et al. Citation2010).
For our simple C model, we assume complete incorporation of the unharvested fraction of NCA into the soil C pool (Soussana et al. Citation2004).
Example calculation for summer moist flat site: if NCA=5152 kg C ha−1 and utilisation is set at 60% (), then the amount of C returned to the soil C pool as litter is NCA×unutilised proportion of NCA=5152 kg C ha−1×0.40=2061 kg C ha−1.
Ingestion of C by grazing animals and excretion in dung
Of the NCA that is harvested by grazing ruminants, the non-digestible C is excreted, mainly in the form of dung.
In our simple C model, the organic matter digestibility of herbage is assumed to be 80% (Machado et al. Citation2005), and thus approximately 20% of C intake (i.e. the indigestible fraction) is returned to the soil surface as dung. We also assume complete incorporation of the C in dung into the soil C pool.
Example calculation for summer moist flat site: if NCA=5152 kg C ha−1 and utilisation is set at 60% (), then total C ingested (TCI)=5152 kg C ha−1×0.60=3091 kg C ha−1, and the amount of C in dung returned to the pasture=3091 kg C ha−1×0.20 (i.e. organic matter digestibility set at 80%)=618 kg C ha−1.
Redistribution of C in animal excreta
As noted earlier, C in dung is not returned evenly in hill country paddocks. In the base scenario of the present study, we assumed that 67% of the total excreta in the paddock was returned to the flat areas, 27% to the easy slopes and 4% to the steep areas (). As the flat areas were assumed to only occupy 9% of the paddock area it is apparent that there is a heavy concentration of excreta return to the flat areas and correspondingly lower returns on the easy and, particularly, the steep slopes. To calculate the quantities of dung C returned to each of the slope categories it was firstly necessary to calculate the total quantity of dung C excreted by grazing animals in the paddock. In the previous section the amount of dung C (618 kg C ha−1) excreted by animals from herbage ingested from the flat area of the paddock was calculated. As the flat area occupies 9% of the 1 ha paddock, this equated to 618×0.09=55.6 kg C in dung originating from herbage ingested from the flat area. Similar calculations give the quantities of dung C generated from herbage on the easy and steep areas as 420.7 kg C and 133.0 kg C, respectively. Therefore, the total quantity of dung C excreted in the paddock=55.6+420.7+133.0=609.3 kg C. Of this, it was assumed that 67% was deposited on the flat area, amounting to 609.3×0.67=408.2 kg C. This is equivalent to 408.2/0.09=4536 kg C ha−1. Similar calculations were made for the other slope categories.
Outputs of C from the terrestrial C pool
Plant and animal respiration and enteric fermentation
Approximately 37% of TCA is respired directly back to the atmospheric pool by the plant (Saggar et al. Citation1997; Soussana et al. Citation2004). The grazing animal additionally represents a major source of loss of fixed C from the terrestrial C pool back to the atmospheric pool. Nearly three quarters of the total ingested C is respired back to the atmosphere as CO2 by the grazing animal, with a further 6% of ingested C entering the atmospheric pool as methane (Soussana et al. Citation2004; Lassey Citation2007). Very little C is accumulated in the animal or in its products (i.e. milk and wool).
In our simple C model, we assume that 37% of TCA is respired by the plant, 72% of ingested C is respired by the animal, 6% of ingested C is lost to the atmosphere as methane and 2% of C ingested is removed in animal products (Soussana et al. Citation2004).
Example calculation for summer moist flat site: amount of C respired by plants=TCA×0.37=19,814 kg C ha−1×0.37=7331 kg C ha−1; amount of C respired by animals=TCI×0.72=3091 kg C ha−1×0.72=2226 kg C ha−1; amount of C emitted as methane=TCI×0.06=3091×0.06=185 kg C ha−1; amount of C removed in animal product=TCI×0.02=62 kg C ha−1.
Respiration at the soil surface
After photosynthesis, soil CO2 efflux or respiration at the soil surface (R s) constitutes the largest exchange of CO2 between terrestrial ecosystems and the atmosphere (Bahn et al. Citation2008; Brown et al. Citation2009). Respiration at the soil surface comprises CO2 respired from both plant roots and the microbial decomposition of soil organic compounds and detritus. Root respiration, which is generally the predominant source of soil CO2 efflux, has no influence on soil C balance as it is not associated with organic inputs or outputs from the soil. The remaining soil respiration is due to the activity of micro-organisms and soil fauna. Bacteria and fungi utilise C substrates released from plants (through root exudation, root turnover and litter excreta return) and from mineralisation of soil organic matter. This primary consumption is then processed through higher trophic levels in the food web, with respiratory losses associated with each step.
The exponential increase in R s with soil temperature is well documented (Bahn et al. Citation2008), but R s is also regulated by root zone volumetric water content, with R s being highest when soils are around field capacity and lowest under conditions of both high and low volumetric water content. There have been very few measurements of R s in New Zealand grazing systems, and only one study that measured R s consistently over a growing season (Brown et al. Citation2009). In that study, total ecosystem respiration was measured in a cattle-grazed pasture in Canterbury, New Zealand, where R s accounted for 84% and above-ground respiration (plant) accounted for 16% of total ecosystem respiration (Brown et al. Citation2009). Using field measurements and modelling, the estimated R s for a total growing season was 19,400 kg C ha−1 (Brown et al. Citation2009). This value is high compared with values of R s for grassland reported in the literature (5636–7004 kg C ha−1 yr−1 (Jones et al. Citation2006); 5150–6110 kg C ha−1 yr−1 (Klumpp et al. Citation2007)), although within the range of 4940–20,350 kg C ha−1 yr−1 reported in an extensive review of R s in grazed pastures and meadows in Europe (Bahn et al. Citation2008).
In constructing our C model, we assumed that the base scenario has been in place for a long period of time and that the soil C on each slope class has therefore reached an equilibrium value as suggested by Haynes & Williams (Citation1993) and in the more recent work of (Parfitt et al. Citation2010). The values for R s for our simple C model for the base scenario were calculated by difference, assuming no change in soil C. The calculated values for R s in our simple model for the base scenario were retained for the other scenarios, even though it is acknowledged that the various scenarios modelled are likely to affect R s (Cao et al. Citation2004). At present there is not sufficient information to alter R s with any confidence for the different scenarios modelled for our case study paddocks.
With the assumption of no change in soil C, R s values for our simple model were calculated to be 13,860 and 6142 kg C ha−1 yr−1 for the flat site, 10,232 and 5693 kg C ha−1 yr−1 for the easy sloped area, and 6556 and 1858 kg C ha−1 yr−1 for the steep sloped area for the summer moist and summer dry sites, respectively.
Soil methane
In agricultural soils, on average, the amount of C emitted as methane by the soil is expected to be approximately equal to that consumed by the soil over a year (Tate et al. Citation2007; Saggar et al. Citation2008) and thus our simple C model does not take into consideration the flux of methane at the soil surface.
Additional outputs from the soil C pool
Whilst loss of C from the soil pool through leaching, runoff and erosion are not considered as direct losses to the terrestrial C pool as such, they are important sources of loss to the soil C pool.
Leaching and runoff (DOC)
Pastoral soils in New Zealand contain large amounts of dissolved organic C (DOC) compared with cropped soils and soils under native vegetation and forestry (Ghani et al. Citation2007). Little is understood of the dynamics of DOC in soils under grazing systems (Van Kessel et al. Citation2009), although large seasonal fluctuations have been measured (Ghani et al. Citation2007). Large losses of DOC were measured from seepage zones in one small plot study in summer moist hill country from areas that had received no or high rates of phosphate fertiliser in the previous 25 years (i.e. 121 and 228 kg DOC ha−1 yr−1) (Parfitt et al. Citation2009). However, losses of DOC through leaching and runoff from pastoral land in New Zealand have been suggested to be much less, at 17 kg DOC ha−1 yr−1 (Scott et al. Citation2006). Studies of DOC in a grassland soil in coastal California indicated that fluxes in the top 20 cm of soil were rapidly attenuated with depth and very little DOC was lost below 1 m (Sanderman & Amundson Citation2008).
In the absence of more comprehensive information on DOC losses through leaching and runoff, in our simple C model, we have taken the more generic value of 17 kg C ha−1 yr−1 for DOC lost from the soil via leaching and runoff as suggested by Scott et al. (Citation2006).
Erosion and runoff (POC)
Although erosion results in a net soil C loss from the landscape, soil C recovery on eroded sites and soil C storage in depositional sites can significantly offset the gross C losses that occur (Jacinthe et al. Citation2001; Dymond Citation2010). In a study in a highly erodible steepland catchment in New Zealand, gross soil C loss was estimated to be approximately 940 kg C ha−1 yr−1, with C recovery on landslip scars (after oxidation) at approximately 610 kg C ha−1 yr−1, and calculated mobilisation rates of 500 kg C ha−1 yr−1 due to landsliding and 440 kg C ha−1 yr−1 due to sheetwash erosion (Page et al. Citation2004). Annual losses of particulate organic carbon (POC) and DOC measured in runoff water in a small grazed hill catchment were approximately 80 kg C ha−1 yr−1 (Lambert et al. Citation2000). Estimates of 53 kg ha−1 yr−1 of POC exported from pastoral land to the ocean in New Zealand have been suggested (Scott et al. Citation2006). Loss of POC from grazed hill country through erosion and runoff are likely to be very site dependent, varying according to topography, climate, soil type and management imposed (Scott et al. Citation2006). For our simple C model, we set the value for POC lost from the soil in erosion at 53 kg POC ha−1 yr−1, as estimated by Scott et al. (Citation2006).
A simple C model: model outputs
Carbon budgets were constructed separately for flat, easy and steep sloped areas of hill country () and are based on the base scenario input parameters in . In , arrows indicate C inputs, outputs and transformations, and C pools are illustrated in boxes. The numbers alongside the arrows represent the amount of C gained, lost or transformed annually through that process. The amounts of C in the various pools and fluxes are expressed per hectare.
Figure 1 Simple C models illustrating C cycles for the summer moist site (upper value for each pool/process) and the summer dry site (lower value for each pool/process) as described for a base scenario (): A, flat sites; B, easy sloped areas; C, steep sloped areas. All values are kg C ha−1 yr−1.
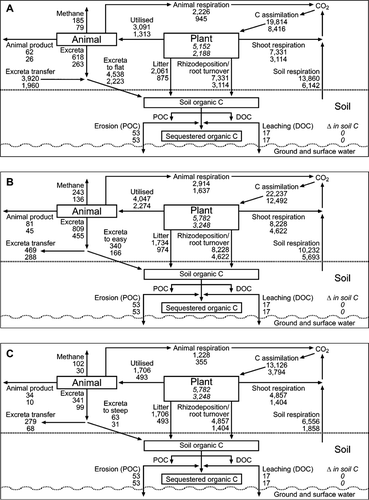
A simple C model () was then constructed for the two 1 ha case study paddocks described in the introduction (). The per hectare values in were adjusted for the proportion of the paddock occupied by each slope category and summed to give an overall C budget for the notional 1 ha paddock ( and )
Figure 2 A simple C model illustrating the C cycle for a notional 1 ha hill country paddock for the summer moist site (upper value for each pool/process) and the summer dry site (lower value of each pool/process) as described for a base scenario ().
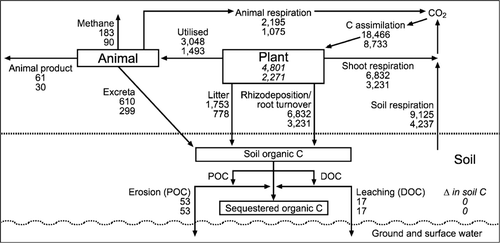
Table 2 Modelled soil C balances for both individual slope categories within notional 1 ha paddocks, consisting of 9% flat sites, 52% easy slopes and 39% steep slopes as described for a base scenario () (all values in kg C yr−1).
For both the summer moist and the summer dry sites, the total inputs of C to the soil, when expressed on a per hectare basis (), were predicted to be highest on the flat/camp areas and to decrease as the slope increased. The main source of C input on all slope categories was rhizodeposition and root turnover, and the high inputs per hectare from this source on the flat/camp and the easy sloped areas are a consequence of the higher NHAs (i.e. C assimilations) on these slope categories. On the flat/camp areas, excreta C was the second largest input to the soil C pool and was over twice the contribution from litter C on these areas at both the summer moist and the summer dry sites. In contrast, on both the easy and steep slope categories, the inputs to the soil C pool from excreta C were predicted to be much smaller than the contribution from litter C. This reflects the importance of animal transfer in the cycling of C in hill country pastures. The predicted inputs of C to the soil C pool were larger on all slope categories at the summer moist site than on the corresponding slope categories at the summer dry site, reflecting the higher values for NHA (and thus TCA and NCA) at the summer moist site.
If it is assumed that the soils on each of the slope categories at the two sites have reached equilibrium and that the soil C is therefore constant, R s must equal the total inputs of C to the soil, less the small losses through leaching and erosion. When expressed on a per hectare basis (), this results in calculated R s values ranging from 13,860 kg C ha−1 yr−1 on the flat/camp area at the summer moist site to 1858 kg C ha−1 yr−1 on the steep slope at the summer dry site.
Once the fluxes of C for each slope class were adjusted for the proportion of the notional 1 ha paddock they occupy ( and ), the net input of C (before considering R s) to the 1 ha paddock was predicted to be 9125 kg C ha−1 yr−1 at the summer moist site and 4237 kg C ha−1 yr−1 at the summer dry site. If soil C is assumed to be at equilibrium, these net inputs of C also equate to the values for R s. In our notional 1 ha paddock, the largest inputs of C (and losses from soil respiration) are from the area in the easy slope category, reflecting the high NHA and the large area of the paddock they are assumed to occupy.
When the assumed herbage utilisation levels were increased above the base levels for all slope categories (i.e. under more intensive grazing management) and excreta transfer was kept at base levels (), the predicted amount of C in litter returned to the soil pool decreased by nearly 45% at both sites. Although excreta C returned to the soil increased by 25% as a result of increased herbage utilisation by the grazing animal, it was only on the flat/camp areas at both sites that this was sufficient to compensate for the decrease in C entering the soil pool through litter. On the easy and steep slopes at both sites, the total amounts of C entering the soil C pool were predicted to decrease and thus there would be an overall loss of soil C within the paddock compared with the base scenario. This decrease in soil C was predicted to be greater at the summer moist site than at the summer dry site (660 versus 286 kg C ha−1 yr−1) (), assuming R s values remained the same as in the base scenario.
Table 3 Summary of soil C inputs minus outputs before (upper value for each scenario) and after (lower value for each scenario (in bold)) allowance for soil respiration for the different scenarios modelled.
When herbage utilisation levels were decreased below base levels for all slope categories (i.e. under a relatively extensive grazing management), the amount of litter returned to the soil pool increased by 40% while the amount of C returned in excreta decreased by 25% and thus there was an overall predicted increase in C inputs to the soil C pool compared with the base scenario for the 1 ha notional paddock at both sites (). This would likely result in an increase in soil C levels until a new equilibrium position was established. Once again, the predicted change in the soil C pool was greater at the summer moist site than the summer dry site. This is because a given percentage change in utilisation has a greater absolute effect at the summer moist site with its higher NHA.
According to our simple C model for grazed hill country, altering the extent of nutrient transfer affects the C inputs at the slope category level but not the overall C input in the 1 ha notional paddock ().
Intensification of the hill country grazing system by adding N fertiliser and improving pasture utilisation was predicted to have multiple impacts on the C cycle. The extra 1 t ha−1 NHA on the easy and steep slopes from the assumed application of N fertiliser translated to an increase in annual NCA for the 1 ha paddock of 7.5% for the summer moist site and 14.3% for the summer dry site. This resulted in increased C inputs to the easy and steep slopes through root growth and rhizodeposition. The increased NHA and the assumed greater pasture utilisation also resulted in greater C inputs to the soil via animal excreta. At the same time, however, the increased pasture utilisation reduced the amount of litter C returned to the soil C pool to approximately 50% of that of the base scenario.
At the summer moist site this decrease in litter C return was larger than the increased C inputs from root growth, rhizodeposition and animal excreta and the overall effect of intensification was predicted to be a small decrease (254 kg C ha−1 yr−1) in soil C for the 1 ha paddock—assuming no change in R s. It is interesting to note that, for the summer dry site, the intensification scenario had the opposite effect on the soil C balance, with intensification predicted to result in an increase in soil C of 214 kg C ha−1 yr−1 for the 1 ha paddock compared with the base scenario. This opposite effect of intensification on the soil C balance at the summer dry site could be explained by the relatively smaller drop in the amount of litter C returned to the soil in the intensification versus the base scenario than was calculated for the summer moist site. This is because in our C model, pasture utilisation (and therefore also litter fall) is expressed as a percentage of NHA. Changing the pasture utilisation by a set percentage will therefore have a larger impact on the actual amounts of herbage returned to the soil as litter on sites with a large NHA than sites with a lower NHA. At the summer dry site, therefore, the increase in the amount of C added to the soil pool by root and rhizodeposition due to the greater NCA on easy and steep slopes, and in excreta C due both to the greater NCA and to the greater utilisation levels, was more than sufficient to compensate for the reduced litter return.
In our simple C model, nutrient transfer effects on soil C balance are important at the slope category level, but not at the paddock scale, as a gain in soil C in one slope category translates to a loss in soil C in another. Excreta C makes up only a very small fraction of C input to the soil C pool; thus, from a C point of view, changes in patterns of excreta deposition have small effects on paddock-scale soil C balance.
Losses of soil C due to runoff, erosion and leaching are very poorly quantified and few studies have addressed these forms of soil C loss in hill country in general, or at each slope category in particular. However, in the simple C model presented here, the quantities of C leaving the soil C pool in runoff, erosion and leaching are small compared with that being respired from the soil on an annual basis.
N cycling in grazed hill country with emphasis on the soil N pool
Background
The total N contents of New Zealand topsoils generally fall within the range 0.15–1.00%. Virtually all (>95%) of this total N is contained within the soil organic matter and its distribution within the soil usually corresponds with the distribution of organic carbon. Total N concentrations tend to be highest in the topsoil and decrease rapidly with depth.
Inorganic forms of N in soils include nitrate (NO3) and ammonium (NH4). Although these two inorganic N forms make up only a small fraction of the total N in soils at a given point in time, they play a key role in N cycling in the soil/plant animal system.
In the following sections, the key processes in the N cycle are reviewed briefly, with emphasis on information drawn from studies on hill country in New Zealand. At the end of each section, the methods used to derive values for that particular transformation in the N cycle for the two case study paddocks are outlined.
A simple N model: model development
Nitrogen inputs
Symbiotic N fixation. Although symbiotic N fixation through clover is the main method of supplying N to the grass sward of New Zealand pastures, hill country pastures usually have low clover contents (Ledgard et al. Citation1987; Roach et al. Citation1996; Gillingham et al. Citation2003) and these result in low N inputs through fixation. At the dry summer dry hill country site, Blennerhassett (Citation2002) reported higher clover contents on easy sites than on steep sites that commonly had little or no clover in the sward. By contrast, Ledgard et al. (Citation1987) reported that at the summer moist site, which had higher annual rainfall, there was little difference between slope categories in total legume production but that there were marked seasonal and species difference between slope categories. Measurements of N fixation in hill country pastures have ranged from 13–85 kg N ha−1 yr−1 (Grant & Lambert Citation1979; Ledgard et al. Citation1987) to 107–392 kg N ha−1 yr−1 for more intensively farmed lowland pastures (Hoglund et al. Citation1979).
Nitrogen fixation by legumes is proportional to clover dry matter (DM) accumulation. In hill country, the proportionality constants vary between campsites and sloping sites with Ledgard et al. (Citation1987) suggesting values of 0.040 kg N fixed per kilogramme of clover DM grown on sloping sites and 0.030 kg N fixed per kilogramme of clover DM grown on campsites. The lower proportionality constant values in campsites were associated with high soil inorganic N levels resulting from the transfer of dung and urine by grazing sheep to these sites.
In this study, N fixation was calculated from legume production () using the proportionality constant values proposed by Ledgard et al. (Citation1987).
Example calculation for summer moist flat site: if NHA=14,310 kg DM ha−1, legume content=13% and the proportionality constant=0.03 kg N kg legume DM-1, then N fixation=14,310×0.13×0.03=55.8 kg N ha−1.
Non symbiotic N fixation
Grant & Lambert (Citation1979) suggested that in a poorly developed hill country pasture, N fixation by non-symbiotic organisms and atmospheric deposition can be, together, as important as clover N fixation. They estimated that approximately 21 kg N ha−1 yr−1 was fixed by non-symbiotic N fixation and atmospheric deposition using the intercept of a regression between N fixation and legume DM production. Lambert et al. (Citation1982) used a value of 3 kg N ha−1 yr−1 for deposition by rain in an N balance developed for the Ballantrae summer wet hill country site.
In this study, non-symbiotic N fixation was assumed to be 13 kg N ha−1 yr−1, and a contribution of 3 kg N ha−1 yr−1 was assumed for N deposition by rain (Lambert et al. Citation1982).
N fertiliser
The use of N fertiliser in hill country farming has increased considerably in recent years (MacLeod & Moller Citation2006). A range of N fertiliser responses of 1–33 kg DM/kg N have been reported in hill pastures (Luscombe Citation1980; Ball & Field Citation1982; Lambert et al. Citation1986; Clark & Lambert Citation1989; Gillingham et al. Citation1998; Blennerhassett Citation2002; Gillingham et al. Citation2007, Citation2008a). Factors affecting the magnitude of response include location (Ball & Field Citation1982; Gillingham et al. Citation2008a, Citation2008b), fertiliser application rate (Ball et al. Citation1982; Gillingham et al. Citation2007, Citation2008a, Citation2008b), season, slope (Ledgard et al. Citation1983; Gillingham et al. Citation1998; Blennerhassett Citation2002; Gillingham et al. Citation2003), aspect (Blennerhassett Citation2002; Gillingham et al. Citation2003) and pasture species composition (Ledgard et al. Citation1982; Rumball Citation1991).
In this study, no N fertiliser is applied in the base scenario, but in the more intensive scenario it is assumed that a total of 100 kg N ha−1 is applied annually with an average response of 10 kg DM/kg N.
Nitrogen translocations
Plant uptake
NHA on New Zealand hill country is often limited by the availability of soil N (Blennerhassett Citation2002; Gillingham et al. Citation2007, Citation2008a). Blennerhassett (Citation2002) noted that, after climate and its interaction with topography, N availability was the largest determinant of NHA. He observed that in the absence of added N, NHA was less than half its potential. Total N concentrations in hill pastures are usually in the range 2.5–4% (Ledgard et al. Citation1987; Bowatte Citation2003).
In this study, plant N uptake was calculated from the data in on NHA and herbage N concentrations and then subtracting the contribution of N fixation to the herbage N pool.
Example calculation for summer moist flat site: if NHA=14,310 kg DM ha−1, herbage N concentration=0.0378 kg N/kg DM and N fixation=55.8 kg N ha−1, then plant N uptake=14,310×0.0378–55.8=485.1 kg N ha−1.
N return in litter
As noted earlier, the quantity of litter is very dependent on the pasture utilisation. Bowatte et al. (Citation2006) estimated 11–40 kg N ha−1 yr−1 would be returned to the soil in litter. Sakadevan et al. (Citation1993a) estimated 40 and 84 kg N ha−1 added in litter over 7 months (winter–summer) at low-fertility and high-fertility sites at Ballantrae, respectively. In this study, N returned in litter was calculated as the difference between N taken up by plants and that ingested by animals.
Example calculation for summer moist flat site: if NHA=14,310 kg DM ha−1, herbage N concentration=0.0378 kg N/kg DM and pasture utilisation=60%, then N returned in litter=14,310×0.0378×0.4=216.4 kg N ha−1.
Nitrogen ingestion by grazing animals, retention in animal products and excretion in dung and urine
Nitrogen removed in animal products in hill country studies has been reported to be 4–30 kg N ha−1 yr−1 (Lambert et al. Citation1982; Ledgard Citation2001; Bowatte et al. Citation2006). Generally, this is 3–15% of N ingested (Whitehead Citation1995), with the remainder returned in excreta to the paddock. Because of the uneven distribution of excreta in hill country pastures, 35–48 kg N ha−1 could be transferred annually from sloped hill sites to flat stock camping areas (Ledgard et al. Citation1987; Ledgard Citation2001; Bowatte et al. Citation2006). Animal transfer to camp areas does not necessarily represent a total loss from the system, as the N can still potentially be taken up by plants. If, however, the annual input of N to camp areas is larger than the maximum amount of N that could be taken up by pasture, given the existing environmental conditions, then some of this transfer to camp areas does represent an effective loss of N from the system.
In this study, N ingested by grazing animals was calculated from the N taken up by plants and the pasture utilisation assumed for the site, and it was assumed that 10% of ingested N was retained in animal products. This is near the middle of the range reported by Whitehead (Citation1995). The remainder of the ingested N was excreted in dung and urine. Of the N excreted, 71% was assumed to be in urine and the remainder in dung. This partitioning of excreta N was near the middle of the range reported by Lambert et al. (Citation1982).
Example calculation for summer moist flat site: if NHA=14,310 kg DM ha−1, herbage N concentration=0.0378 kg N/kg DM and pasture utilisation=60%, then N ingested by grazing animals=14,310×0.0378×0.6=324.6 kg N ha−1. Ingested N from the summer moist flat site that is retained in animal products=324.6×0.1=32.5 kg N ha−1, excreted N in urine=324.6×0.9×0.71=207.4 kg N ha−1 and excreted N in dung=324.6×0.9×0.29=84.8 kg N ha−1.
Return of N in animal excreta
As noted earlier, N in dung and urine is not returned evenly in hill country paddocks, but is concentrated in the flat campsite areas. To calculate the quantities of N returned to each slope category, a similar approach to that described earlier for the C cycle was used. Using this approach, the total amounts of N excreted from herbage grazed on the flat easy, easy and steep sites amounted to 26.3 kg N, 183.9 kg N and 54.9 kg N for the flat, easy and steep slopes, respectively. Therefore, the total quantity of N excreted in the paddock=26.3+183.6+54.9=265.1 kg N. Of this, it was assumed that 67% was deposited on the flat area, amounting to 265.1×0.67=177.6 kg N. This is equivalent to 408.2/0.09=1973 kg N ha−1. Of this, 71% (1401 kg N ha−1) was as urine N and 29% (572 kg N ha−1) was as dung N. Similar calculations were made for the other slope categories.
N transformations
Mineralisation and immobilisation
Mineralisation is the general term for the conversion of organic N to inorganic N as either NH4 or NO3. Immobilisation is the conversion of inorganic N to organic N. Within hill country farming systems, this immobilisation can occur through plant uptake, incorporation by soil micro-organisms and by chemical interactions with complex carbon compounds. In this review, N immobilised in the above-ground plant pool is treated separately from N immobilised below-ground. In practice, it is very difficult to measure absolute rates of mineralisation and below-ground immobilisation in soils and therefore net mineralisation (or immobilisation) is reported instead.
The rate of incorporation of N into the microbial biomass and other soil organic fractions will depend on factors influencing the mineralisation/immobilisation turnover. These factors include the C/N ratio of organic residues and environmental conditions. Ball et al. (Citation1982) reported that many topsoils in the hill country of the lower North Island contain very large quantities of nitrogen, rendered largely unavailable by the relatively wide C/N ratio (12–19 depending on the micro-climate (Ledgard et al. Citation1982, Citation1987; Lambert et al. Citation2000)) of the soil organic matter present. Sakadevan et al. (Citation1993a) estimated that 119 and 250 kg N ha−1 was mineralised from low-fertility and high-fertility hill sites over 7 months (winter–summer) in summer moist hill country at Ballantrae, respectively.
In this study, net mineralisation of N was calculated by difference, assuming that from year to year the inorganic pool of N will remain constant and that all other inputs and outputs from the inorganic N pool can be either measured or estimated.
Example calculation for summer moist flat site: if N returned in urine=1401.2 kg N ha−1, N added in atmospheric deposition=3 kg N ha−1, N taken up by plants=485.1 kg N ha−1, N lost by leaching=420.4 kg N ha−1, N lost by ammonia volatilisation=462.4 kg N ha−1 and N lost through emission of nitrous oxide (N2O)=19.7 kg N ha−1, then net mineralisation=1401.2+3.0–485.1–420.4–462.4–19.7=− 16.6 kg N ha−1.
Nitrification
Nitrification is a particularly important process in grazed grassland. It is the conversion of relatively immobile NH4-N (derived from animal excreta, fertiliser and the mineralisation of soil organic N) to NO3-N, which is vulnerable to leaching and denitrification. Bowatte (Citation2003) observed that soils collected from hill country sites had lower nitrification rates (0.3–11.3 µg NO3-N g soil−1 day−1) than soils collected from intensively managed lowland sites (16.6–18.3 µg NO3-N g soil−1 day−1), with the nitrification rate in soils collected from hill country being highly variable. Soil collected from flat stock campsites had greater nitrification rates (0.6–11.3 µg NO3-N g soil−1 day−1) than soils from steep slopes (0.3–2.1 µg NO3-N g soil−1 day−1).
In this study, both NH4-N and NO3-N are included in the soil inorganic N pool; nitrification is thus not explicitly recognised in this model, but it has indirect effects on the rates of N leaching and denitrification.
Nitrogen outputs
Ammonia volatilisation
Volatilisation of ammonia (NH3) is an important pathway of N loss from the urine voided by grazing animals (Ball et al. Citation1979; Whitehead Citation1995). Measured volatilisation losses of urine N under New Zealand conditions vary from 15–18% (Ball et al. Citation1979), through 12–25% (Sherlock & Goh Citation1984), to 17–36% (Carran et al. Citation1982). However, few (if any) of these measurements were made in hill country, and Lambert et al. (1982) suggested that the hill country environment was unlikely to be conducive to volatilisation. Accordingly, they assumed volatilisation losses of only 5% of excretal N when constructing an N balance for hill country. Bowatte (Citation2003), using a passive sampling technique, detected apparently large emissions of ammonia from urine patches at two hill country sites. Using a calibration curve that was developed on flat land dairy farms, Bowatte (Citation2003) suggested that volatilisation losses could be in the range of 21–51% of applied urine N, but stressed that calibration curves would need to be developed under hill country conditions before reliable quantitative estimates of N volatilisation could be made.
Based on the work of Bowatte (Citation2003), it was assumed in this study that 33% of urine N was volatilised. It is acknowledged that this leads to very high predicted volatilisation losses and more work quantifying such N losses is identified at the end of this review as a high priority for future work.
Example calculation for summer moist flat site: if N returned in urine=1401.2 kg N ha−1, then N lost through volatilisation=1401.2×0.33=462.4 kg N ha−1.
Nitrous oxide emission
Theoretically, the potential for N2O losses from grazed pastures is high due to the high levels of readily available C in surface soils and the high concentrations of NO3-N present in soil under urine patches (de Klein & Eckard Citation2008). To date, there have been very few measurements of N2O emissions from grazed pastures in hill country. Carran et al. (Citation1995) suggested that very low rates of N2O emissions (0.5 kg N2O-N ha−1 yr−1 or less) could be expected from low-fertility hill lands. More recently, Hoogendoorn et al. (Citation2008) estimated losses ranging from 0.1 to 0.5 kg N2O-N ha−1 over a 6 week period in spring time from relatively intensively grazed hill country at two contrasting locations in New Zealand. These estimates were based on measured losses from urine patches and a calculation of the likely number of urine patches from actual stocking rates. These estimates increased to 0.2–5.2 kg N2O-N ha−1 over a 6 week period when fertiliser N was applied at a rate of 300 kg N ha−1 yr−1 (Hoogendoorn et al. Citation2008).
In this study, N2O emissions were assumed proportional to inputs of N in urine, dung and fertiliser, but the proportionality constant varied according to land slope. Emissions of N2O were assumed to be 1%, 0.5% and 0.1% of urine and fertiliser N deposited on campsites, easy slopes and steep slopes, respectively. The lower emission rates assigned to easy and steep slopes reflect the lower nitrification rates commonly observed on these areas.
Example calculation for summer moist flat site: if N returned in urine=1401.2 kg N ha−1, N returned in dung=572.3 kg N ha−1 and N added in fertiliser=0 kg N ha−1, then N lost through emission of N2O=(1401.2+572.3)×0.01=19.7 kg N ha−1.
Leaching
In the past, leaching has not been considered a major N loss mechanism in hill country pastures (Sakadevan et al. Citation1993b; Lambert et al. Citation2003). Bowatte (Citation2003) measured very low nitrification activities in soils collected from steep slopes in hill country, and suggested that this would mean there was little potential for loss of N by leaching from steep slopes. However, Bowatte (Citation2003) also found that soils collected from campsites had higher nitrification activities and this led him to suggest that leaching could be a significant mechanism of N loss from stock campsites that receive large quantities of N in animal excreta.
This has been supported by more recent studies. Parfitt et al. (Citation2009) reported annual leaching losses of 38–274 kg N ha−1 from a high-fertility hill site that had N fertiliser applied at an annual rate of 300 kg N ha−1 yr−1. Hoogendoorn et al. (in preparation) measured leaching losses over a 3 year period of 84–196 kg N ha−1 yr−1 from flat and easy sloping areas of intensively grazed hill country that had received N fertiliser at rates of 0–300 kg N ha−1 yr−1.
In this study, 30% of N in urine and 7% of N in fertiliser applied to campsites was assumed to be leached. The higher assumed loss from urine reflected the higher rate of N application in urine patches. It was assumed that no leaching occurs on easy and steep slopes, because of the low rates of nitrification.
Example calculation for summer moist flat site: if N returned in urine=1401.2 kg N ha−1 and N added in fertiliser=0 kg N ha−1, then N lost through leaching=1401.2×0.3=420.4 kg N ha−1.
A simple N model: model output
Simple N balances () were developed for each of the slope categories in the notional 1 ha paddocks described earlier (). In , the boxes represent N pools and the arrows represent N inputs, outputs and transformations. The numbers alongside the arrows represent the amount of N (kg ha−1) gained, lost or transformed annually through that process, and the numbers in the boxes represent the amount of N (kg ha−1) entering that pool annually. The N cycles for each slope class were adjusted according to the proportion of the paddock occupied by that slope category, and were then combined to give an overall N budget for the notional 1 ha paddock as a whole () and the soil in that paddock ().
Figure 3 Simple N models illustrating N cycles for the summer moist site (upper values for each pool/process) and the summer dry site (lower value for each pool/process) as described for the base scenario (): A, flat sites; B, easy sloped areas; C, steep sloped areas. All values are kg N ha−1 yr−1.
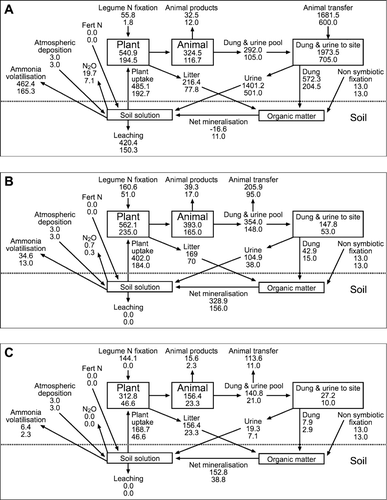
Figure 4 A simple N model illustrating the N cycle for a notional 1 ha hill country paddock for the summer moist site (upper value for each pool/process) and the summer dry site (lower value for each pool/process) as described for a base scenario ().
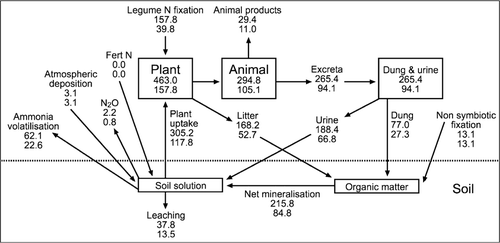
Table 4 Modelled soil N balances for individual slope categories within notional 1 ha paddocks consisting of 9% flat sites, 52% easy slopes and 39% steep slopes.
At both sites, the model predicted that soil N was accumulating on the flat sites and was slowly being depleted on easy slopes (). The main driver was removal of N in herbage by grazing animals, combined with little N return in animal excreta to sloped sites. Ledgard et al. (Citation1987) also predicted that accumulation of N on steep slopes would be very slow, or could be negative under high pasture utilisation. Although the transfer of nutrients by grazing animals is well known (Gillingham & During Citation1973; Gillingham Citation1978; Ledgard et al. Citation1987; Ledgard Citation2001), Ledgard (Citation2001) pointed out that these significant losses of N due to transfer of excreta are often ignored in published estimates and models of N flows and balances in grazed hill country systems.
On a whole-paddock basis (), the model predicts that N was accumulating in the summer moist hill country paddock, but in the summer dry paddock there was a small net loss of N. In other words, N losses due to retention in animal products, leaching and volatilisation were higher than N inputs by legume N fixation, non-symbiotic fixation and atmospheric deposition. This difference is mainly due to lower inputs through N fixation at the summer dry site.
Ledgard (Citation2001) reported a similar N balance for hill country soils. Comparison of the N balances reported in this study and that of Ledgard (Citation2001) confirms that the degree of N surplus or deficit on a particular slope category will depend on the levels of N input, pasture growth rate, pasture utilisation and excretal transfer that are assumed. The output from both models indicates that there are likely to be large areas of hill country where virtually all of the N added to the system by N fixation and other processes may be lost, and that it is likely that some areas may experience an annual N deficit. Clearly, such a deficit could not be maintained for an extended period, as soil reserves would become depleted and DM production would decrease until losses again matched inputs.
When pasture utilisation is assumed to be higher than in the base scenario, the predicted depletion of soil N on easy and steep slopes becomes more marked and the build-up of soil N on the campsites increases (). This is because of reduced N return in plant litter on easy and steep slopes and increased N transfer by grazing animals to campsites. The overall N surplus predicted to occur at the summer moist site in the base scenario becomes smaller as the pasture utilisation is increased. At the summer dry site, the predicted N deficit in the base scenario becomes even greater as pasture utilisation is increased. Reducing pasture utilisation below that assumed in the base scenario has the opposite effect.
Table 5 Soil N loss/gain for a range of scenarios in two contrasting hill country pastures.
As the distribution of excreta across the paddock is made more even than was assumed in the base scenario, there is greater N return to easy and steep slopes and soil N is predicted to increase in these areas (). There is a corresponding decrease in the predicted build-up of soil N on the campsites. Interestingly, although redistributing excretal N more evenly across slope classes does not change the total amount of excretal N returned, it is predicted to increase the overall build-up of N within the paddock (). This is a consequence of an assumption within the model that excretal N deposited on flat campsites is more susceptible to loss—primarily through leaching and denitrification—than excretal N deposited on steep slopes. This assumption, in turn, is based on experimental observations by Bowatte (Citation2003) that nitrification rates on campsites in hill country are very much greater than on steep slopes. This effect is not observed in the C cycle (), where changing the assumed pattern of excretal deposition affects C build-up or depletion on individual slopes, but is predicted to have no effect on the C balance at a whole-paddock level. This might not be the case if changes in the pattern of excretal deposition affected R s on the various slopes, but at present there is no experimental information available to assess this.
The impact of intensifying the production system on the N cycle is complex. Intensification in this study was assumed to involve the addition of N fertiliser at the rate of 100 kg N ha−1 to increase NHA by 1000 kg DM ha−1 on the easy and steep slopes. Intensification was also assumed to increase pasture utilisation, particularly on easy and steep slopes, and this was then assumed to increase the proportion of animal excreta returned to these easy and steep slopes.
Thus, while the addition of N fertiliser and increased pasture utilisation might normally be expected to increase N losses, a more even distribution of animal excreta across the landscape tends to reduce these losses. This is particularly the case in hill country where nitrification rates on easy and steep slopes seem to be much lower than on campsites. The outcome of these two conflicting trends depends very much on the assumptions made about the effects of intensification. Of particular importance may be the effect of N fertiliser addition on the legume percentage in the sward and the resulting N fixation.
In this study, management intensification was predicted () to reduce the build-up of organic N on campsites and increase the build-up of organic N on easy and steep slopes, and in the paddock as a whole. Losses of N through volatilisation were predicted to increase (data not presented) because more urine N was produced and the model assumed that a fixed percentage (33%) of urine N was volatilised, irrespective of where in the paddock it was deposited. In contrast, leaching of N was predicted to decrease (data not presented). This was because less excretal N was deposited in campsites where the high nitrification rates lead to a high risk of leaching.
General discussion
Both the C and N cycles in hill country pastoral farming are characterised by large fluxes in and out of the farming system and, by comparison, relatively small annual accumulations or depletions of the various soil pools. However, the mechanisms by which these inward and outward fluxes are generated differ greatly between C and N.
Most of the C cycling through the soil/plant/animal system in a year is sourced from the atmosphere through photosynthesis in that same year. Little or no C cycles through the soil/plant/animal system more than once without being returned to the atmosphere, and the annual quantity of C ingested by grazing animals is only approximately 25% of that fixed annually (according to the assumptions made in the simple C model presented in this paper).
In contrast, although in extensive hill country systems most of the N input also comes from the atmosphere via N fixation, most of this N cycles through the soil/plant/animal system several times before being lost from the system. The annual quantity of N ingested by grazing animals is 300–400% of that added to the system annually by fixation, and most of this ingested N (>80%) is returned to the soil in animal excreta. This contrasts with <25% of ingested C returned in animal excreta. Grazing animals are therefore much more influential in the redistribution of N than C in hill country pastures.
The transfer of nutrients by grazing animals means that, in hill country pastures, there is little or no accumulation of N on sloping areas, regardless of the level of N input or NHA (and thus NCA). There is, however, accumulation of N in flat camp areas. This accumulation of soil N will eventually lead to N saturation of the soil and greater losses through leaching, denitrification and volatilisation from these areas. There is some evidence that these increased losses are already occurring (e.g. Parfitt et al. (Citation2009) and Hoogendoorn et al. (in preparation)).
From the simple C model for grazed hill country presented here, the three major drivers that determine whether the soil C balance is positive or negative are:
| 1. | the amount of NCA (and hence the amount of C going to root growth and rhizodeposition | ||||
| 2. | the amount of C in litter returned to the soil pool | ||||
| 3. | soil respiration rate R s. | ||||
These calculated R s values were much lower than the R s value of 19,400 kg C ha−1 yr−1 reported by Brown et al. (Citation2009), but were closer to values presented in a summary of studies for grassland soils in the UK and in Europe (Jones et al. Citation2006; Klumpp et al. Citation2007; Bahn et al. Citation2008).
At a given NCA and R s, utilisation of herbage grown is a key determinant of soil C balance. The NCA that is not incorporated as litter is consumed by the grazing animal. However, the grazing animal is a major source of loss of C from the terrestrial pool as a large proportion of consumed C is respired as CO2 and lost in the production of methane in the rumen. Thus, a relatively small change in the partitioning of NCA into ingested (utilised) versus litter C can have a large impact on the amount of C entering the soil C pool.
Intensification potentially has at least three effects on the soil C balance. Firstly, increasing NCA leads to an increase in net C inputs to the soil through rhizodeposition and root turnover. Secondly, increasing pasture utilisation results in a decrease in C inputs from litter. Thirdly, changes in N availability as a grazing system intensifies may affect R s (Wang & Fang Citation2009). Which of these processes dominates in a given situation determines whether intensification will result in an increase or a decrease in soil C. In constructing our C model, the values used for R s for the different slope classes were kept constant across the different scenarios modelled, although it is acknowledged that R s may change with different management practices in the longer term (Lal Citation2001; Cao et al. Citation2004; Wang & Fang Citation2009). With this assumption of unchanging R s values, the intensification scenario as defined in this paper was predicted to result in a decrease in C inputs at the summer moist site and an increase in C inputs at the summer dry site.
These contrasting predictions arise from the assumed increase in NCA and in pasture utilisation as a result of intensification. Pasture utilisation was assumed to increase as a result of intensification from 60%, 70% and 50% on flat/camp, easy and steep slopes respectively in the base scenario to 79%, 83% and 76% on the equivalent slopes in the intensive scenario. Because these increases in pasture utilisation are expressed in percentage terms, the impact on the quantity of litter C returned to the soil depends on the absolute amount of NCA and hence is larger at the summer moist site than at the summer dry site. In contrast, the increase in NCA as a result of N fertiliser application was identical at both sites, with each site increasing in NCA by 328 kg C ha−1 yr−1. The combined impact of these two factors resulted in a decrease in predicted soil C for the summer moist site and an increase in soil C at the summer dry site.
When attempting to extrapolate these predicted effects of intensification on C cycling to other hill country paddocks, it is important to recognise that the impact on C inputs to the soil of increasing NHA accumulation by a given amount is much greater than the impact of improving utilisation and thereby increasing the amount of DM eaten by the same amount. This is shown in . Increasing NHA by 2 t DM ha−1 yr−1 results in an increase in C inputs of approximately 1750 kg C ha−1 yr−1, whereas increasing animal intake by the same 2 t DM ha−1 yr−1 decreases C inputs by only 550 kg C ha−1 yr−1—less than a third as much. This may provide a ‘rule of thumb’ for predicting the likely effects of intensification on soil C in hill country. If intensification increases NHA (and NCA), there is likely to be an increase in C inputs to the soil unless at the same time there is at least a three times larger increase in DM eaten. This was predicted to be the case at the summer moist site in this study, which had very high levels of NHA; however, even here, the predicted decrease in soil C was small (254 kg C ha−1 yr−1). The possibility of an increase in soil C with management intensification aligns with the observations made by (Schipper et al. Citation2010) that, across a range of grazed hill country sites in New Zealand, soil C increased by an average of 520 kg C ha−1 yr−1 over a 27 year period.
Figure 5 Predicted annual changes in soil C from the base scenario, with changes in NHA and animal DM harvested according to our simple C model.
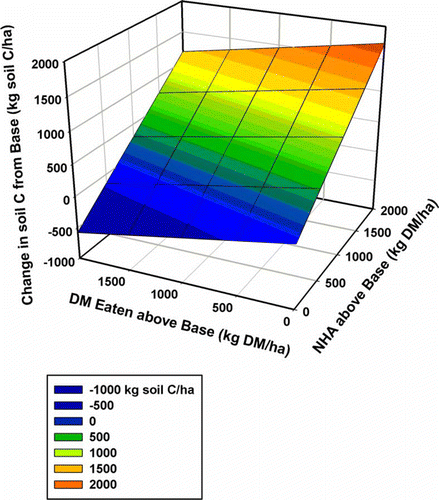
In contrast, Lambert et al. (Citation2000) reported a decrease of 200 kg C ha−1 yr−1 in soil C with intensification at a southern North Island summer moist hill country site. If soil C in hill country is decreasing over time, then our modelling work suggests that this is more likely to be from increases in R s than from decreases in C inputs from litter. To make further progress will require much more accurate estimates of R s. Of particular importance is the link between C and N. The sloping areas of hill country paddocks are chronically N deficient and this lack of N is one of the reasons thought to be responsible for lower R s rates on easy and steep slopes. The extent to which increased N inputs from fertiliser and excreta as a result of intensification will stimulate R s on these slopes is not known at present, but is likely to be very important.
It is interesting to estimate how much the values for R s set in our simple C model would need to decrease to achieve the annual gain in soil C measured by (Schipper et al. Citation2010), and likewise to calculate the decrease in R S needed to attain the decrease in soil C measured by Lambert et al. (Citation2000). For the base scenario, the weighted average value for R s (calculated by assuming soil C on each slope category had reached equilibrium) in the notional 1 ha paddock was 9125 kg C ha−1 yr−1 for the summer moist site and 4237 kg C ha−1 yr−1 for the summer dry site (). Using the work of Lambert et al. (Citation2000), where 200 kg C ha−1 yr−1 was measured as being lost from soils to 7.5 cm depth, and assuming that soil C to 10 cm accounts for approximately 30% of the total soil C to 100 cm (Scott et al. Citation2002), we can estimate that perhaps approximately 667 kg C ha−1 yr−1 was lost from 100 cm in the study of Lambert et al. (Citation2000). To achieve such a reduction in soil C, R s at the summer moist site would need to be 9792 kg C ha−1 yr−1 () and at the summer dry site R s would need to be 4904 kg C ha−1 yr−1 if soil C loss was 667 kg C ha−1 yr−1; these figures represent an increase of 7 and 16% respectively.
Table 6 Comparison of soil respiration rates used in the simple C model with those calculated by difference using soil C loss values reported by Lambert et al. (Citation2000) and soil C gain values reported by Schipper at al. (2010).
Considering the work of (Schipper et al. Citation2010) where a gain of 520 kg C ha−1 yr−1 from soils to 30 cm depth was reported, R s would need to be 8605 kg C ha−1 yr−1 at the summer moist site and 3717 kg C ha−1 yr−1 at the summer dry site. These are decreases of 6% and 12% respectively.
The findings of (Schipper et al. Citation2010) and Lambert et al. (Citation2000) serve to make the point that the changes in R s necessary to result in a predicted decrease in soil C are very small compared with the likely range in R s from 1856 to 13,855 kg C ha−1 yr−1 () across slope categories at these two sites—a difference of approximately 750%. Given this large spatial variability, detecting differences in R s as a result of intensification of hill country farming will require very careful measurements.
The impact of intensification on N cycling is also complex and will vary from farm to farm. Because hill country pastures are normally acutely N-limited, increasing NHA normally requires increasing N inputs either by adding N fertilisers or increasing N fixation through improving overall soil fertility and boosting legume production. The application of N fertiliser to increase NHA, combined with better pasture utilisation, would normally be expected to increase N accumulation in campsites and also losses of N through leaching and volatilisation. However, where the more intensive grazing systems involve rotational grazing with large mobs, there may be more even distribution of excreta (as suggested by O'Connor (Citation1966)) and this might significantly reduce the loss of N from flat campsites by leaching but may, over time, increase these forms of N loss on the easy slope areas. More work is required on the effect of increasing intensity of grazing on the distribution of excreta in hill country pastures.
The predicted impact of intensification on N leaching in this study is very dependent on assumptions about patterns of pasture utilisation and excretal deposition, and further assumptions about the fate of excretal N once it has been deposited on the soil. Our model of N cycling in hill country did not account for the leaching of DON from the root zone of pasture plants. Knowledge of the movement of both DOC and DON through the soil profile, and the relationship between DOC and DON in the soil under grazed pastures, is not well quantified (Ghani et al. Citation2003; Ghani et al. Citation2007), with information from grazed areas of hill country particularly lacking (Parfitt et al. Citation2009). The implications of management intensification on the storage and leaching of DOC and DON in hill country soils must be understood in order to adequately understand and model C and N pools and fluxes in this environment (Sanderman & Amundson Citation2008; Sanderman et al. Citation2008).
Another major gap in current knowledge concerns the volatilisation of ammonia from urine. The process of volatilisation is very dependent on factors such as temperature, soil water content and soil pH, and will therefore vary between sites and also between seasons. This is reflected in the wide range of measured volatilisation losses (12–36% of urine N) reported by New Zealand workers (Ball et al. Citation1979; Carran et al. Citation1982; Sherlock & Goh Citation1984). None of these studies were conducted on hill country, but measurements by Bowatte (Citation2003) suggested that volatilisation losses from urine in hill country could be substantial. Based on the work of Bowatte (Citation2003), volatilisation losses in this modelling study were set at 33% of urine N and this resulted in volatilisation being the greatest mechanism of N loss in the case study paddocks. More experimental work is required to establish the actual range of volatilisation losses in hill country. However, the model does indicate that the N cycle in hill country is more complex than on flat land and it should not automatically be assumed that intensification of grazing systems will always increase N losses through leaching, volatilisation or denitrification.
Conclusions and areas for future work
The most important gap in our knowledge of both the C and N cycles relates to the pattern of soil biological activity across the hill country landscape. Biological activity determines the rate of soil respiration in the C cycle and determines the rate of nitrification and denitrification in the N cycle. These are the key processes determining the extent of soil accumulation and losses of C and N. In qualitative terms, it is well understood that biological activity is high in flat campsites and reduces as slopes become steeper. However, considerably more work is required to develop a more quantitative understanding of these processes across different slope categories and aspects.
The work presented here suggests that leaching losses of N from high-fertility hill country pastures may be substantial, and recent experimental findings of Parfitt et al. (Citation2009) and Hoogendoorn et al. (in preparation) support that view. It is generally agreed that such significant N leaching is only likely to occur in the small areas of flat campsites, but the simple C and N models constructed in this paper suggest that this could still translate into significant N leaching losses over a paddock as a whole. Although both experimental and modelling studies suggest that there may be significant leaching of N below the root zone in campsite areas, the subsequent fate of that leached N is unknown. The patterns of above- and below-ground water movement in hill country are complex (e.g. Petch (Citation1984)) and it may well be that some of this leached N re-emerges at the surface in other parts of the paddock where it could be taken up again by plants. This requires further study.
The simple models presented here demonstrate that while land use intensification results in increased inputs of both C and N to the soil, intensification is also likely to result in increased losses of C and N from the soil pool. Which of these processes dominates in a given situation determines whether intensification will result in an increase or decrease of soil C and N. Based on our simple C model, we postulate that if intensification increases NHA by one unit there is likely to be an increase in soil C unless, at the same time, there is at least a three-unit increase in DM eaten/harvested.
Acknowledgements
This review was undertaken as part of the Foundation for Research Science and Technology funded Rural Futures programme (C10X0809 Rural Futures), Objective 2: Farm System Responses. The authors would like to acknowledge the contribution of three anonymous reviewers for their encouragement and helpful suggestions.
Related Research Data
References
- Bahn , M , Rodeghiero , M Anderson-Dunn , M . 2008 . Soil respiration in European grasslands in relation to climate and assimilate supply . Ecosystems , 11 : 1352 – 1367 .
- Ball , PR and Field , TRO . 1982 . “ Responses to nitrogen as affected by pasture characteristics, season and grazing management ” . In Nitrogen fertilisers in New Zealand agriculture , Edited by: Lynch , PB . 45 – 64 . Wellington : New Zealand Institute of Agricultural Sciences .
- Ball , PR , Luscombe , PC and Grant , DA . 1982 . “ Nitrogen on hill country ” . In Nitrogen fertilisers in New Zealand agriculture , Edited by: Lynch , PB . 133 – 147 . Wellington : New Zealand Institute of Agricultural Sciences .
- Ball , R , Keeney , DR , Theobald , PW and Nes , P . 1979 . Nitrogen balance in urine-affected areas of a New Zealand pasture . Agronomy Journal , 71 : 309 – 314 .
- Blennerhassett JD 2002 Massey University Palmerston North, , New Zealand Pasture growth constraints on dry steep east coast hill country Unpublished PhD thesis
- Bowatte , S , Tillman , R , Carran , A and Gillingham , A . 2006 . Can phosphorus fertilisers alone increase levels of soil nitrogen in New Zealand hill country pastures? . Nutrient Cycling in Agroecosystems , 75 : 57 – 66 .
- Bowatte WMSD Massey University Palmerston North, , New Zealand 2003 Urine nitrogen in hill country pasture soils Unpublished PhD thesis
- Brown , M , Whitehead , D , Hunt , JE , Clough , TJ , Arnold , GC , Baisden , WT and Sherlock , RR . 2009 . Regulation of soil surface respiration in a grazed pasture in New Zealand . Agricultural and Forest Meteorology , 149 : 205 – 213 .
- Cao , G , Tang , Y , Mo , W , Wang , Y , Li , Y and Zhao , X . 2004 . Grazing intensity alters soil respiration in an alpine meadow on the Tibetan plateau . Soil Biology and Biochemistry , 36 : 237 – 243 .
- Carran , RA , Ball , PR , Theobald , PW and Collins , MEG . 1982 . Soil nitrogen balances in urine affected areas under two moisture regimes in Southland . New Zealand Journal of Experimental Agriculture , 10 : 377 – 381 .
- Carran , RA , Theobald , PW and Evans , JP . 1995 . Emission of nitrous oxide from some grazed pasture soils in New Zealand . Australian Journal of Soil Research , 33 : 341 – 352 .
- Clark DA , Lambert MG 1989 . The use of fertiliser nitrogen for increased animal production from New Zealand hill pastures . Proceedings of the XVI International Grassland Congress . Pp. 1269 – 1270 .
- de Klein , CAM and Eckard , RJ . 2008 . Targeted technologies for nitrous oxide abatement from animal agriculture . Australian Journal of Experimental Agriculture , 48 : 14 – 20 .
- de Klein C , Hoogendoorn C , Manderson A , Saggar S , Giltrap D , Briggs C , Rowarth J 2009 . Refinement of the framework for upscaling nitrous oxide emissions from hill country . AgResearch Limited, Palmerston North, New Zealand, MAF-POL 0809-11061 . 77 p.
- Dymond , JR . 2010 . Soil erosion in New Zealand is a net sink of CO2 . Earth Surface Processes and Landforms , 35 : 1763 – 1772 .
- Frame , J . 1992 . “ Feeding value of grass ” . In Improved grassland management , Edited by: Frame , J . 146 – 160 . Ipswich : Farming Press Books .
- Ghani , A , Dexter , M and Perrott , KW . 2003 . Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilisation, grazing and cultivation . Soil Biology and Biochemistry , 35 : 1231 – 1243 .
- Ghani , A , Dexter , M , Carran , RA and Theobald , PW . 2007 . Dissolved organic nitrogen and carbon in pastoral soils: the New Zealand experience . European Journal of Soil Science , 58 : 832 – 843 .
- Gillingham AG Massey University Palmerston North, , New Zealand 1978 Phosphorus cycling in grazed, steep hill country Unpublished PhD thesis
- Gillingham , AG and During , C . 1973 . Pasture production and transfer of fertility within a long-established hill pasture . New Zealand Journal of Experimental Agriculture , 1 : 227 – 232 .
- Gillingham , AG , Gray , MH and Smith , DR . 1998 . Pasture responses to phosphorus and nitrogen fertilisers on dry hill country . Proceedings of the New Zealand Grassland Association , 60 : 135 – 140 .
- Gillingham , AG , Morton , JD and Gray , MH . 2003 . The role of differential fertiliser application in sustainable management of hill pastures . Proceedings of the New Zealand Grassland Association , 65 : 253 – 257 .
- Gillingham , AG , Morton , JD and Gray , MH . 2007 . Pasture responses to phosphorus and nitrogen fertilisers on East Coast hill country: Total production from easy slopes . New Zealand Journal of Agricultural Research , 50 : 307 – 320 .
- Gillingham , AG , Morton , JD and Gray , MH . 2008a . Pasture responses to phosphorus and nitrogen fertilisers on east coast hill country: 2. Clover and grass production from easy slopes . New Zealand Journal of Agricultural Research , 51 : 85 – 97 .
- Gillingham , AG , Morton , JD , Gray , MH and Roberts , AHC . 2008b . Pasture responses to phosphorus and nitrogen fertilisers on east coast hill country: 3. Production from steep slopes . New Zealand Journal of Agricultural Research , 51 : 425 – 437 .
- Grant , DA and Lambert , MG . 1979 . Nitrogen fixation in pasture 5. Unimproved North Island hill country Ballantrae New Zealand . New Zealand Journal of Experimental Agriculture , 7 : 19 – 22 .
- Haynes , RJ and Williams , PH . 1993 . Nutrient cycling and soil fertility in the grazed pasture ecosystem . Advances in Agronomy , 49 : 119 – 199 .
- Hoglund , JH , Crush , JR , Brock , JL , Ball , R and Carran , RA . 1979 . Nitrogen fixation in pasture New Zealand Journal of Experimental Agriculture , 7 : 45 – 51 .
- Hoogendoorn , CJ , De Klein , CAM , Rutherford , AJ , Letica , S and Devantier , BP . 2008 . The effect of increasing rates of nitrogen fertiliser and a nitrification inhibitor on nitrous oxide emissions from urine patches on sheep grazed hill country pasture . Australian Journal of Experimental Agriculture , 48 : 147 – 151 .
- Jacinthe , PA , Lal , R and Kimble , JM . 2001 . Organic carbon storage and dynamics in croplands and terrestrial deposits as influenced by subsurface tile drainage . Soil Science , 166 : 322 – 335 .
- Jones , SK , Rees , RM , Kosmas , D , Ball , BC and Skiba , UM . 2006 . Carbon sequestration in a temperate grassland; management and climatic controls . Soil Use and Management , 22 : 132 – 142 .
- Klumpp , K , Soussana , JF and Falcimagne , R . 2007 . Effects of past and current disturbance on carbon cycling in grassland mesocosms . Agriculture, Ecosystems and Environment , 121 : 59 – 73 .
- Lal , R . 2001 . World cropland soils as a source or sink for atmospheric carbon . Advances in Agronomy , 71 : 145 – 191 .
- Lambert MG Renton SW Grant DA 1982 Nitrogen balance studies in some North Island hill pastures . In: Nitrogen balances in New Zealand Ecosystems Gandar PW 35 – 39 Palmerston North, , New Zealand Plant Physiology Division, Department of Scientific and Industrial Research
- Lambert , MG , Devantier , BP , Nes , P and Penny , PE . 1985 . Losses of nitrogen, phosphorus, and sediment in runoff from hill country under different fertiliser and grazing management regimes . New Zealand Journal of Agricultural Research , 28 : 371 – 379 .
- Lambert , MG , Clark , DA , Grant , DA and Costall , DA . 1986 . Influence of fertiliser and grazing management on North Island moist hill country. 2. Pasture botanical composition . New Zealand Journal of Agricultural Research , 29 : 1 – 10 .
- Lambert , MG , Clark , DA , Mackay , AD and Costall , DA . 2000 . Effects of fertiliser application on nutrient status and organic matter content of hill soils . New Zealand Journal of Agricultural Research , 43 : 127 – 138 .
- Lambert , MG , MacKay , AD , Devantier , BP , McDougall , DB , Barker , DJ and Park-Ng , ZA . 2003 . Redefining the production potential of hill pastures using fertiliser nitrogen . Proceedings of the New Zealand Grassland Association , 65 : 35 – 40 .
- Lassey , KR . 2007 . Livestock methane emission: From the individual grazing animal through national inventories to the global methane cycle . Agricultural and Forest Meteorology , 142 : 120 – 132 .
- Leake , JR , Ostle , NJ , Rangel-Castro , JI and Johnson , D . 2006 . Carbon fluxes from plants through soil organisms determined by field 13CO2 pulse-labelling in an upland grassland . Applied Soil Ecology , 33 : 152 – 175 .
- Ledgard , SF . 2001 . Nitrogen cycling in low input legume-based agriculture, with emphasis on legume/grass pastures . Plant and Soil , 228 : 43 – 59 .
- Ledgard , SF , Sheath , GW and Gillingham , AG . 1982 . Influence of some soil and pasture components on the growth of hill country pastures. 1. Winter and spring production . New Zealand Journal of Experimental Agriculture , 10 : 239 – 244 .
- Ledgard , SF , Brier , GJ , Gillingham , AG and Sheath , GW . 1983 . Influence of some soil and pasture components on the growth of Hill Country Pastures 2. Response to nitrogen fertilizer . New Zealand Journal of Experimental Agriculture , 11 : 251 – 256 .
- Ledgard , SF , Brier , GJ and Litter , RA . 1987 . Legume production and nitrogen fixation in hill pasture communities . New Zealand Journal of Agricultural Research , 30 : 413 – 422 .
- Luscombe , PC . 1980 . Nitrogen fertilizer responses on hill country pastures . Proceedings of the New Zealand Grassland Association , 41 : 155 – 162 .
- Machado , CF , Morris , ST , Hodgson , J and Fathalla , M . 2005 . Seasonal changes of herbage quality within a New Zealand beef cattle finishing pasture . New Zealand Journal of Agricultural Research , 48 : 265 – 270 .
- Mackay , AD . 2008 . Impacts of intensification of pastoral agriculture on soils: Current and emerging challenges and implications for future land uses . New Zealand Veterinary Journal , 56 : 281 – 288 .
- MacLeod , CJ and Moller , H . 2006 . Intensification and diversification of New Zealand agriculture since 1960: An evaluation of current indicators of land use change . Agriculture, Ecosystems and Environment , 115 : 201 – 218 .
- O'Connor , KF . 1966 . The improvement and utilization of tussock grasslands: a scientist's point of view: Cycling nitrogen for production . Proceedings of the New Zealand Grassland Association , 28 : 59 – 77 .
- Page , M , Trustrum , N , Brackley , H and Baisden , T . 2004 . Erosion-related soil carbon fluxes in a pastoral steepland catchment, New Zealand . Agriculture, Ecosystems & Environment , 103 : 561 – 579 .
- Parfitt , RL , Yeates , GW , Ross , DJ , Mackay , AD and Budding , PJ . 2005 . Relationships between soil biota, nitrogen and phosphorus availability, and pasture growth under organic and conventional management . Applied Soil Ecology , 28 : 1 – 13 .
- Parfitt , RL , MacKay , AD , Ross , DJ and Budding , PJ . 2009 . Effects of soil fertility on leaching losses of N, P, and C in hill country . New Zealand Journal of Agricultural Research , 52 : 69 – 80 .
- Parfitt , RL , Yeates , GW , Ross , DJ , Schon , NL , Mackay , AD and Wardle , DA . 2010 . Effect of fertilizer, herbicide and grazing management of pastures on plant and soil communities . Applied Soil Ecology , 45 : 175 – 186 .
- Parsons , AJ and Chapman , DF . 2000 . “ The principles of pasture growth and utilization ” . In Grass: Its production and utilization , 3rd edn , Edited by: Hopkins , A . 31 – 89 . Oxford, , United Kingdom : Blackwell Science .
- Parsons AJ , Rowarth JS , Newton PCD 2009 . Managing pasture for animal and soil carbon . Proceedings of the New Zealand Grassland Association 71 : 77 – 84 .
- Petch RA University of Waikato Waikato, , New Zealand 1984 Studies of source area hydrology under indigenous forest and pastoral agriculture in the Hapuakohe Range, North island, New Zealand Unpublished PhD thesis
- Roach , CG , Nemaia , EKK , Ledgard , SF , Brier , GJ and McLay , CDA . 1996 . Effects of long term differences in fertiliser history on hill country: seasonal pasture production, legume growth and soil phosphorus status . Proceedings of the New Zealand Grassland Association , 57 : 105 – 109 .
- Rumball , PJ . 1991 . Performance of several subtropical grasses in Northland hill pastures . New Zealand Journal of Agricultural Research , 34 : 375 – 382 .
- Rutledge , S , Campbell , DI , Baldocchi , D and Schipper , LA . 2010 . Photodegradation leads to increased carbon dioxide losses from terrestrial organic matter . Global Change Biology , 16 : 3065 – 3074 .
- Saggar , S , Hedley , C and Mackay , AD . 1997 . Partitioning and translocation of photosynthetically fixed 14C in grazed hill pastures . Biology and Fertility of Soils , 25 : 152 – 158 .
- Saggar , S , Mackay , AD and Hedley , CB . 1999 . Hill slope effects on the vertical fluxes of photosynthetically fixed 14C in a grazed pasture . Australian Journal of Soil Research , 37 : 655 – 666 .
- Saggar , S , Tate , KR , Giltrap , DL and Singh , J . 2008 . Soil–atmosphere exchange of nitrous oxide and methane in New Zealand terrestrial ecosystems and their mitigation options: A review . Plant and Soil , 309 : 25 – 42 .
- Sakadevan , K , Hedley , MJ and Mackay , AD . 1993a . Mineralisation and fate of soil sulphur and nitrogen in hill pastures . New Zealand Journal of Agricultural Research , 36 : 271 – 281 .
- Sakadevan , K , Mackay , AD and Hedley , MJ . 1993b . Influence of sheep excreta on pasture uptake and leaching losses of sulfur, nitrogen and potassium from grazed pastures . Australian Journal of Soil Research , 31 : 151 – 162 .
- Sanderman , J and Amundson , R . 2008 . A comparative study of dissolved organic carbon transport and stabilization in California forest and grassland soils . Biogeochemistry , 89 : 309 – 327 .
- Sanderman , J , Baldock , JA and Amundson , R . 2008 . Dissolved organic carbon chemistry and dynamics in contrasting forest and grassland soils . Biogeochemistry , 89 : 181 – 198 .
- Schipper , LA , Parfitt , RL , Ross , C , Baisden , WT , Claydon , JJ and Fraser , S . 2010 . Gains and losses in C and N stocks of New Zealand pasture soils depend on land use . Agriculture, Ecosystems and Environment , 139 : 611 – 617 .
- Scott DT , Baisden WT , Davies-Colley R , Gomez B , Hicks DM , Page MJ , Preston NJ , Trustrum NA , Tate KR , Woods RA Localized erosion affects national carbon budget . Geophysical Research Letters 2006 33 L01402. 4 p.
- Scott , NA , Tate , KR , Giltrap , DJ , Tattersall , Smith C , Wilde , HR , Newsome , PJF and Davis , MR . 2002 . Monitoring land-use change effects on soil carbon in New Zealand: Quantifying baseline soil carbon stocks . Environmental Pollution , 116 : S167 – S186 .
- Sherlock , RR and Goh , KM . 1984 . Dynamics of ammonia volatilization from simulated urine patches and aqueous urea applied to pasture I. Field experiments . Fertilizer Research , 5 : 181 – 195 .
- Sitaula , BK , Bajracharya , RM , Singh , BR and Solberg , B . 2004 . Factors affecting organic carbon dynamics in soils of Nepal/Himalayan region: A review and analysis . Nutrient Cycling in Agroecosystems , 70 : 215 – 229 .
- Soussana , JF , Loiseau , P , Vuichard , N , Ceschia , E , Balesdent , J , Chevallier , T and Arrouays , D . 2004 . Carbon cycling and sequestration opportunities in temperate grasslands . Soil Use and Management , 20 : 219 – 230 .
- Tate , KR , Ross , DJ , Saggar , S , Hedley , CB , Dando , J , Singh , BK and Lambie , SM . 2007 . Methane uptake in soils from Pinus radiata plantations, a reverting shrubland and adjacent pastures: Effects of land-use change, and soil texture, water and mineral nitrogen . Soil Biology and Biochemistry , 39 : 1437 – 1449 .
- Van Kessel , C , Clough , T and Van Groenigen , JW . 2009 . Dissolved organic nitrogen: An overlooked pathway of nitrogen loss from agricultural systems? . Journal of Environmental Quality , 38 : 393 – 401 .
- Wang , W and Fang , J . 2009 . Soil respiration and human effects on global grasslands . Global and Planetary Change , 67 : 20 – 28 .
- Whitehead , DC . 1995 . Grassland nitrogen , Wallingford : CAB International .
- Zhang , B , Valentine , I and Kemp , P . 2005 . Modelling the productivity of naturalised pasture in the North Island, New Zealand: A decision tree approach . Ecological Modelling , 186 : 299 – 311 .