
ABSTRACT
A plot study was undertaken to investigate the interactive effect of three defoliation intervals (one-leaf, two-leaf and three-leaf stage in perennial ryegrass (PR) and one-leaf, two-leaf and four-leaf stage in tall fescue (TF)) and two defoliation height residuals (low, 5 cm and high, 8 cm) on production parameters of TF compared with PR plots. DM yields, botanical composition and tiller densities of both species were all generally greater at the longest defoliation intervals (TF-4 and PR-3). However, this consistently resulted in the poorest herbage quality in the TF-4 treatment. The herbage quality was higher in TF-2 compared with TF-4; however, the DM yield, tiller density and proportion of sown species in the sward were reduced. It is not possible to achieve an adequate compromise between DM yield, botanical composition, tiller survival and herbage quality in TF, throughout the year, when defoliation occurs at the two-leaf and the four-leaf stages.
Introduction
New Zealand’s pastoral systems rely on a mixture of perennial ryegrass (PR; Lolium perenne L.) and white clover (Trifolium repens L.) (Charlton and Stewart Citation1999). PR pasture growth and quality, and subsequent animal production can be limited during periods of moisture and temperature stress during summer and autumn (Litherland et al. Citation2002), which has increased interest in alternative permanent pasture species. Tall fescue (TF; Festuca arundinacea Schreb., syn., Schedonorus arundinaceus and Lolium arundinaceum) is a perennial grass with the potential to out-yield PR and supply quality forage during a dry summer and autumn (McCallum et al. Citation1992). The development of suitable TF cultivars has increased its use in pastoral agriculture (Easton and Pennell Citation1993) but for maximum growth TF must be managed differently from PR due to morphological differences between the two species (Fulkerson and Donaghy Citation2001; Donaghy et al. Citation2008; Hendriks et al. Citation2016). Grazing management based on leaf stage has been well defined for PR swards to optimise productivity and persistence (Fulkerson and Donaghy Citation2001; Turner et al. Citation2006). However, there is currently limited information available regarding the optimal leaf stage to defoliate TF swards under field conditions. Results from a glasshouse study by Donaghy et al. (Citation2008) identified the two-leaf stage of regrowth as the minimum defoliation interval for TF and the four-leaf stage as the maximum defoliation interval, in terms of optimum herbage quality and plant regrowth. The present study was conducted to validate the results of Donaghy et al. (Citation2008) under field conditions and multiple defoliations, and to compare the performance of TF swards with PR swards. A study was undertaken to investigate the interactive effect of defoliation at different leaf stages and to different defoliation heights on the dry matter (DM) yield, herbage quality, botanical composition and tiller density of TF compared with PR.
Methods
Site
The experiment (November 2012–November 2014) was undertaken on the Massey University Dairy 4 farm, 5 km south of Palmerston North (40.397431°S, 175.624011°E), on a Tokomaru silt loam. Soil pH and nutrient contents (75 mm depth) prior to the time of sowing were: pH 5.8, Olsen phosphorus (P) 30 mg/L, sulphur (S) 7 mg/kg, potassium (K) 133 mg/kg, calcium (Ca) 5780 mg/kg, magnesium (Mg) 80 mg/kg and sodium (Na) 32 mg/kg. The mean annual rainfall at the site over the three years (2012–2014) was 766 mm, 15% less than the 10-year average of 899 mm (collected from the NIWA/AgResearch Weather Station, Agent No. 21963, Palmerston North, 2014). The experimental site (19 m × 22 m) was sprayed with 2 L/ha Roundup® Transorb® (540 g/L a.i. of glyphosate) before power and Dutch harrowing. The experiment comprised 36 plots (6 m × 1.2 m), laid out as described below, and sown on 16 November 2012 with a vee-ring roller-drill followed by light chain harrows to cover the seed. Half of the plots, chosen randomly, were sown with TF (cv. Easton (MaxP endophyte); 25 kg/ha) and white clover (cv. Kopu II and Tribute; each sown at 2 kg/ha) and the other half with PR (cv. Bealey (NEA2 endophyte); 28 kg/ha) and white clover (cv. Kopu II and Tribute; each sown at 2 kg/ha). Cropmaster 15 (15.2% nitrogen (N); 10% P, 10% K and 7.7% S) was applied at 200 kg/ha on 15 November 2012.
Experimental design
The 36 plots were subject to different defoliation treatments to investigate the interactive effect of different defoliation intervals and defoliation heights under field conditions. The plots were defoliated with one of two contrasting residual defoliation height treatments – low (L; defoliation height of 5 cm) and high (H; defoliation height of 8 cm) and three defoliation intervals coinciding with the regrowth and full emergence of 1, 2 or 3 leaves/tiller for PR plots and 1, 2 or 4 leaves/tiller for TF plots. The treatment combinations (2 species × 2 defoliation heights × 3 defoliation intervals) were arranged in a randomised complete block design. All plots were defoliated to 8 cm prior to the commencement of the treatment defoliations on 4 October 2013. There was a total of six defoliations prior to the commencement of the experiment and these occurred on 23 January, 26 February, 18 April, 4 June, 4 August and 18 September 2013 before shading reduced tiller appearance and pasture quality due to canopy closure.
Herbage production and defoliation
Plots were defoliated using a rotary mower when the treatment reached the appropriate leaf development stage (1, 2, 3 or 4 leaves) and were mown according to the treatment defoliation height residual (L or H). The dates of all defoliation events during the experimental period are presented in . DM yield was measured by collecting herbage from one mower width (0.46 m), down the length of the plot (5 m) and total wet weight was recorded. A sub-sample of herbage approximately 150 g of wet weight was oven-dried for at least 48 h at 70°C and component dry weights recorded. Available DM yield for the plot at each defoliation event was calculated as the total wet weight multiplied by the DM%. Total available DM yield for the experimental period (4 October 2013–7 November 2014) was calculated as the sum of all available DM yields collected across all defoliation events. The defoliation events were each defined as occurring within a season: flowering period1 (4 October–31 December), summer/autumn (1 January 30 April), winter/spring (1 May 30 September) and flowering period2 (1 October 7 November). Total available DM yield for each season was calculated as the sum of all DM yields collected within each season (as defined above).
Table 1. Defoliation events of tall fescue (TF) and perennial ryegrass (PR) plots subjected to different defoliation intervals (one-leaf stage, -1; two-leaf stage, -2; three-leaf stage, -3; four-leaf stage, -4) during the experimental period.
Botanical composition
Herbage samples for botanical composition were collected at random locations within plots on 27 May 2014, when defoliation events for all treatments coincided (), and at the end of the experiment period (23 October 2014 for PR plots and 7 November 2014 for TF plots). Four samples were collected per plot by cutting 20 cm × 10 cm wide strips to ground level within each plot prior to defoliation. The samples were then separated into sown species (TF or PR), white clover and other [species including grasses and weed] and each sample was oven-dried at 70°C to a constant weight. The botanical composition was calculated on a dry weight basis.
Herbage nutritive value
Herbage samples were collected prior to commencement of the experiment, on 4 August 2013 and prior to defoliation on 15 November 2013, 27 May 2014 and at the end of the experiment on 7 November 2014, for herbage quality analysis. Samples collected on 4 August 2013 prior to experiment commencement were bulked according to species for all plots. At successive sampling dates, samples for each plot were analysed individually. A sample of approximately 150 g wet weight was ‘hand plucked’ from each plot. Samples were freeze-dried, ground to pass through a 1-mm sieve and analysed by near-infrared reflectance spectroscopy (Hills Laboratories, Hamilton) to determine N and dry organic matter digestibility (DOMD). Metabolisable energy (ME) was calculated as DOMD × 0.16, according to Roughan and Holland (Citation1977). Crude protein (CP) was calculated as N × 6.25.
Tiller density
Plots were sampled for tiller density at establishment (23 January 2013) and at the end of the experiment (23 October for PR plots and 7 November for TF plots). At establishment, tiller density was measured in all plots. Tiller density (tillers/m2) was calculated as plant density (plants/m2) multiplied by the average tiller number per plant. Plant density was measured by counting the total number of plants in a randomly selected 1 m segment of one of the internal rows in each plot and multiplying by the number of rows per m2. The average tiller number per plant was measured by counting plants and tillers for each species in a randomly selected 1 m segment of one of the rows (four samples per species) and calculating an average tiller number per plant across all plots according to species (TF or PR). At the end of the experiment, tiller density was calculated as the average number of tillers counted per plot, divided by the area sampled. Five randomly selected transects (17.4 × 15 cm) along identifiable rows were cut to ground level and the number of sown tillers was counted in each plot.
Statistical analysis
All statistical analyses were performed using Minitab™ version 10.51 (Minitab 10.51 Extra Statistical Software 1995, www.minitab.com). Available DM yield, cumulative DM yield, botanical composition, herbage quality and tiller density at the end of the experiment were analysed using a general linear model including the fixed effects of species, defoliation height, defoliation interval and all two-way and three-way interactions between species, defoliation height and defoliation interval. The three-leaf RG and four-leaf TF were considered equivalent for the analysis of variance. Tiller density at establishment was analysed using a general linear model including the fixed effect of species. Least square means were obtained and used for multiple mean comparisons using the Least Significant Difference test.
Results
Herbage DM yields
The TF plots had 57% greater cumulative DM yield on average than the PR plots (P < 0.001) () and a longer defoliation interval resulted in increased cumulative DM yields irrespective of species (P < 0.001) (). Defoliation interval had an effect on available DM yield across all periods and the TF plots under four-leaf defoliation interval (TF-4) resulted in 50% greater cumulative DM yield compared with the TF under one-leaf defoliation interval (TF-1), while the PR plots under three-leaf defoliation interval (PR-3) resulted in 41% greater cumulative DM yield compared with the PR under one-leaf defoliation interval (PR-1) (P < 0.05).
Table 2. Available dry matter (DM) yield and cumulative DM yield (kg DM/ha) of tall fescue (TF) and perennial ryegrass (PR).
Table 3. Available dry matter (DM) yield and cumulative DM yield (kg DM/ha) of tall fescue (TF) and perennial ryegrass (PR) subjected to defoliation at different defoliation intervals (one-leaf stage, -1; two-leaf stage, -2; three-leaf stage, -3; four-leaf stage, -4) during the experimental period.
Across all seasons, the TF-4 had the greatest available DM yield, except during the flowering period during year 1 where TF-2 had the highest yield compared to all other species and defoliation interval combinations (). There was a two-way interaction (P < 0.05) between species and defoliation interval for available DM yield during both flowering periods. There was no interactive effect of species and defoliation interval on cumulative DM yield.
Three-way interactions between species, defoliation interval and defoliation height are not reported as only one of four growth periods (summer/autumn) was significant (P < 0.01). Available DM yield was greater in the low and high defoliation TF-4 plots (P < 0.05) compared with all other treatment combinations. Additionally, low defoliation height resulted in greater DM yield compared with the high defoliation height in the TF-4 plots (5260 and 4161 kg DM/ha, respectively). The interaction had disappeared by winter/spring, resulting in no difference in available DM yield for the remainder of year 2.
Botanical composition
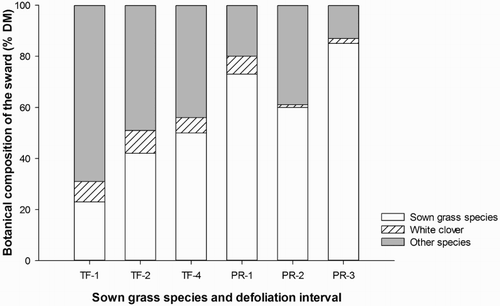
Botanical composition data measured in May 2014 (8 months after the experiment commenced) showed that overall the proportion of sown grass species in the PR plots was nearly double that of the TF plots (73% and 38%, respectively) (). There was a significant interaction between sown grass species and defoliation interval. In the TF plots, the proportion of sown grass species increased with increasing leaf stage (P < 0.05) and subsequently, the proportion of other species declined (P < 0.05). Conversely, this effect of defoliation interval on proportion of sown grass species in the plot was not mirrored in the PR plots, and the proportion of sown grass species was greater in PR-1 and PR-3 compared with PR plots under two-leaf defoliation interval (PR-2) (P < 0.05).
Figure 1. Effect of defoliation interval (one-leaf stage, -1; two-leaf stage, -2; three-leaf stage, -3; four-leaf stage, -4) and sown grass species (TF, tall fescue; PR, perennial ryegrass) on the botanical composition (%DM, dry matter) of the sward during year 2 (27 May 2014). The standard error bars are 8.3% for sown grass species, 1.2% for white clover and 7.5% for other species.
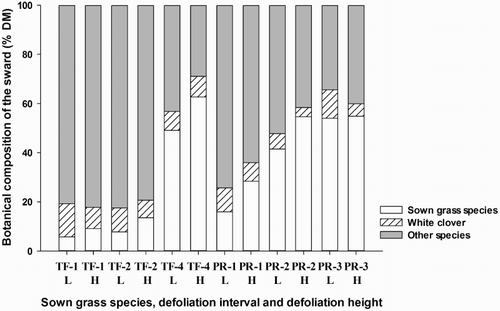
Botanical composition data measured at the end of the experimental period showed a significant (P < 0.001) interaction between species, defoliation interval and defoliation height for the proportions of sown grass species (). The proportion of sown grass species was greater in the PR plots than in the TF plots (P < 0.05) except in the PR-1 L and TF-1 L, and TF-4 L and PR-3 L plots which did not differ significantly. In the PR plots, the proportion of sown grass species increased concomitantly with leaf stage at defoliation and a further increase was observed at a high compared with low defoliation intensity at each leaf stage (P < 0.05). There was no further increase in the PR-3 L plots resulting in no difference between the PR-2 H, PR-3 L and PR-3 H plots. The proportion of sown grass species remained >15% in all PR plots and close to 50% of sown grass species was maintained in the PR-2 H, PR-3 L and PR-3 H plots. In the TF plots, the proportion of sown grass species in the TF-4 L and TF-4 H plots remained close to 50%. Although in the TF-4 plots, the proportion of sown grass species was greater (P < 0.05) under high compared with low defoliation intensity. These plots had higher proportions of sown grass species compared with TF-1 and TF-2 plots (P < 0.05) where there was no effect of defoliation interval and height between TF-1 and TF-2 plots and the proportion of sown species declined to <15%.
Figure 2. Effect of leaf stage (one-leaf stage, -1; two-leaf stage, -2; three-leaf stage, -3; four-leaf stage, -4), defoliation height; low (L, 5 cm) and high (H, 8 cm) and sown grass species (TF, tall fescue; PR, perennial ryegrass) on the botanical composition (%DM, dry matter) of the sward at the end of the experimental period (October/November 2014; year 2). The standard error bars are 5.8% for sown grass species, 0.7% for white clover and 5.7% for other species.
Herbage quality
The herbage quality values are presented in . The CP content was greater at the shortest defoliation interval (one-leaf stage) compared with the longest defoliation interval (three- or four-leaf stage) in TF and PR plots (P < 0.001) across all periods.
Table 4. Herbage quality values (metabolisable energy, ME; crude protein, CP%) of tall fescue (TF) and perennial ryegrass (PR) subjected to different defoliation intervals (one-leaf stage, -1; two-leaf stage, -2; three-leaf stage, -3; four-leaf stage, -4.
The ME content declined between the shortest defoliation interval (one-leaf stage) and the longest defoliation interval (three- or four-leaf stage) in TF and PR plots (P < 0.001) across all seasons except in PR during the flowering period in year 2, where there was no significant difference in ME content between PR-3 and PR-1. The CP and ME contents were generally not significantly different when comparing the two species with their corresponding leaf stage combinations.
Tiller density
The tiller densities of the two species prior to commencing the experiment were on average higher in PR plots than in TF plots (3071 ± 33 vs. 1193 ± 98 tillers/m2, respectively) (P < 0.001) (). By the end of the experiment, the average tiller density declined in the PR plots to 1706 ± 143 tillers/m2 and in the TF plots to 819 ± 66 tillers/m2 (P < 0.001). The tiller density at the end of the experiment was greater in the PR plots for all treatment combinations compared with the TF plots (P < 0.05) ().
Figure 3. Tiller density (tillers/m2) of tall fescue (TF) and perennial ryegrass (PR) plots under two defoliation heights (low, 5 cm, L; high, 8 cm, H) and subjected to defoliation at different defoliation intervals (one-leaf stage, -1; two-leaf stage, -2; three-leaf stage, -3; four-leaf stage, -4) at the end of the experiment (vertical columns) (October/November 2014). Horizontal dotted lines represent the tiller densities at the commencement of the experiment (January 2013). The standard error bars represent standard errors at P < 0.05.
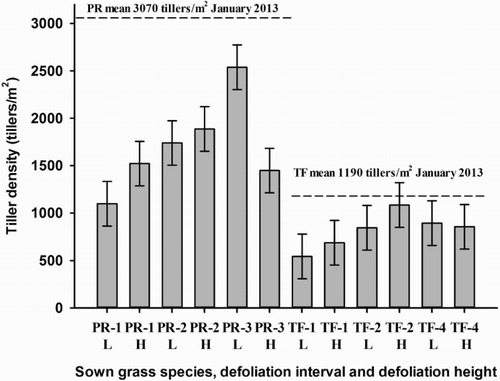
Defoliation height had an effect on tiller density, where at the one- and two-leaf stage low defoliation height decreased tiller density (). In PR-3 and TF-4, the tiller density was greater under low defoliation height compared with high defoliation height.
Discussion
Defoliation interval had a greater effect on all aspects of plant performance than defoliation height; this may be due to the implemented range of defoliation heights from low (5 cm) to high (8 cm) being too small to cause significant differences in the plant parameters measured. Studies reporting significant differences in DM yields, tiller densities and herbage quality in TF and PR typically investigated a range of defoliation heights ≤5 cm and ≥10 cm (Hart et al. Citation1971; Zarrough et al. Citation1983; Tavakoli Citation1993). In the present study, tiller density was further reduced at the one- and two-leaf stage defoliation intervals when defoliated to a low compared with a high defoliation height. This is likely a stress effect, where the lower defoliation height in combination with the shorter defoliation interval resulted in loss of tillers due to reduced water-soluble carbohydrate (WSC) reserves (Fulkerson Citation1994). Conversely, the reduced tiller density observed under high compared with low defoliation height at the longest defoliation intervals (PR-3 and TF-4) was a result of shading causing tiller death (Kays and Harper Citation1974).
The present study provides evidence that irrespective of species, defoliation interval affected DM yield. Both species achieved the highest cumulative yields under the longest defoliation intervals (TF-4 and PR-3). While various factors affect pasture regrowth rate, increased DM yield as the regrowth interval increases is a well-established concept (Brougham Citation1957). The TF plots had a greater cumulative yield compared with PR plots over the 13-month experimental period across all defoliation intervals and heights, which was generally a result of higher yields during the flowering periods and during summer/autumn. The finding that TF out-yielded PR in the present study, particularly during dry periods, is in agreement with results from previous studies (eg Allo and Southon Citation1967; Neal et al. Citation2009; Clark et al. Citation2010). The ability of TF to attain greater yields compared with PR during dry periods has been attributed to the greater tolerance of TF to dry conditions (Allo and Southon Citation1967), due to its greater water use efficiency and deeper root penetration that allow it to extract moisture from a greater depth in the soil compared with PR (Milne et al. Citation1997).
While DM yield is an important indicator of the productive potential of the sward, it is important to consider other plant parameters such as botanical composition, tiller density and herbage quality to understand how defoliation management can affect the persistence and nutritive value of the sward.
Nie et al. (Citation2004) suggested that a measure of persistence is the ability of a pasture to maintain a composition of greater than 70% sown species. In the present study, the proportions of sown species in both TF and PR swards generally increased with defoliation interval. However, the TF plots were only able to maintain above 50% sown species (TF plus legumes) in the sward at the four-leaf stage defoliation interval. When defoliated at the one- or two-leaf stages, the TF plots were not able to maintain this proportion of sown species. The PR plots were able to maintain above 50% sown species in the sward at the PR-2 leaf stage defoliation interval when subject to high defoliation intensity and at the three-leaf stage defoliation interval under both defoliation intensities. Similar trends in persistence were observed in an adjacent paddock-scale experiment (Hendriks et al. Citation2016) and reflect anecdotal farmer experience. Additionally, during the study period, the endophyte status of the TF and PR swards was tested and observed to be at a satisfactory level. Therefore, it is unlikely that endophyte choice had an impact on persistence. Results from the present study indicate that PR has a superior competitive ability against weeds compared to TF under the same management, in agreement with the findings of Bell (Citation1985). In support of this, it was observed (though not measured) that a large proportion of the ‘other species’ that had colonised the TF plots was PR, along with some less productive species (broadleaved weeds and stoloniferous grasses). This indicates that a high proportion of the yield in the TF-1 and TF-2 plots, especially towards the end of the study period, was in fact due to PR.
The ability to maintain a high proportion of sown species is heavily dependent on the defoliation interval and species (Brougham Citation1961; Tozer et al. Citation2011). Since TF produces longer and thicker leaves which have a slower elongation rate and a longer lifespan than those of PR (Kemp et al. Citation2001) then defoliation at the two-leaf stage and below may limit WSC replenishment, and subsequent plant regrowth and persistence of TF (Donaghy et al. Citation2008). Persistence of TF defoliated at the three-leaf stage therefore warrants investigation.
According to Nie et al. (Citation2004), a tiller density of >3000 tillers/m2 for PR-based pastures and >2300 tillers/m2 for TF is indicative of a productive and persistent pasture. At the start of the present study, tiller density was 3070 tillers/m2 for the PR plots and 1190 tillers/m2 for the TF plots. Overall, tiller density decreased in both PR and TF plots during the experiment by 44% and 31%, respectively. Despite the overall lower tiller density in the TF plots, TF plots cumulatively out-yielded PR plots in all treatment combinations over the 13-month experimental period, although as previously mentioned, some of the ‘TF’ yield would have actually been PR in the more frequently defoliated treatments. A study undertaken by Hendriks et al. (Citation2016) compared TF grazed at a four-leaf stage rotation to PR under a three-leaf stage grazing rotation in paddocks adjacent to the present study. Hendriks et al. (Citation2016) reported lower tiller densities in TF compared with PR, despite a greater yield and leaf area index (LAI) in the TF sward. On average, TF tillers are 1.6 times heavier than those of PR (Tavakoli Citation1993) and tiller appearance rate is slower than PR (Kemp et al. Citation2001), which indicates that TF allocates greater resources towards individual tiller development, but less towards tiller initiation, than does PR. Therefore, it is difficult to draw conclusions regarding potential persistence and productive status of pasture species based on tiller density data alone, due to differences in the morphological characteristics influencing tillering activity and subsequent plant regrowth (Jewiss Citation1972). Tiller density is a mechanism for LAI adjustment by which swards optimise light interception in response to defoliation, therefore Matthew et al. (Citation2013) suggested that LAI may be a better measure of productivity and persistence than tiller density alone. The observed trend in the present study of a decline in tiller density for both species with more frequent defoliation is in agreement with studies undertaken by Bell (Citation1985).
Herbage quality has a significant effect on animal production (Litherland et al. Citation2002). Kolver (Citation2000) suggests that herbage quality parameters of >11.5 MJ/kg DM and 18–24% CP are considered to be good quality. The differences in herbage quality components when comparing the TF plots with the PR plots were generally small and indicate that irrespective of species, the defoliation interval had the greatest effect on nutritive value (Fulkerson et al. Citation1998). In addition, the ingress of PR into frequently defoliated TF plots may have positively influenced the herbage quality of these treatments. In the present study, ME and CP contents decreased with increasing defoliation interval, due to plant material becoming more lignified as tissue aged. This is in agreement with studies by Lowe et al. (Citation1999), where herbage quality decreased as the forage matured. The lower CP content under the longer defoliation intervals is in line with results from a number of studies where irrespective of grass species, N levels and therefore CP content decreases with plant maturity (Fulkerson et al. Citation1998). In five out of six measures, the PR plots maintained ME values >10.2 MJ ME/kg DM between the two and three-leaf defoliation intervals. The TF plots maintained ME values >10.2 MJ ME/kg DM when defoliated at the two-leaf stage and ME content declined with less frequent defoliation at the four-leaf stage, in agreement with the findings of Donaghy et al. (Citation2008). During the summer period (2013), ME was <10.2 MJ ME/kg DM in the PR-3 plot, however, the reason for this decline is unclear. The onset of heading results in stem elongation and a subsequent decline in herbage quality (Hunt and Easton Citation1989). Studies have shown that reproductive growth can be related to temperature (Hunt and Field Citation1976). Temperature records from the time of sampling indicate that soil temperature was lower in the summer period during 2014 compared with 2013 (14°C vs. 17°C), and this higher temperature may have caused the decline in ME expressed in PR-3.
This emphasises the dichotomy of TF, with production and persistence being maximised at the four-leaf stage, while defoliation at the two-leaf stage is required to maintain ME contents >10.2 MJ ME/kg DM (Donaghy et al. Citation2008). The most effective defoliation interval for a pasture species is a balance between optimum pasture production, persistence and quality. DM yields, botanical compositions and tiller densities of both species were all generally greatest at the longest defoliation intervals (TF-4 and PR-3). This consistently resulted in the poorest herbage quality in TF-4 while herbage quality was maintained in PR-3. The herbage quality was greater in TF-2, but the DM yield, tiller density and proportion of sown species in the sward were all reduced.
The results from the present study indicate that DM yield, botanical composition and tiller density were compromised when defoliation of TF occurred at or below the two-leaf defoliation interval. Longer defoliation interval (at the four-leaf stage) achieved the highest yield and persistence, but compromised herbage quality. These results support the findings of Fulkerson and Donaghy (Citation2001) and Donaghy et al. (Citation2008) that for optimum growth, persistence and quality, defoliation of PR must occur between the two-leaf and three-leaf stages, whereas TF is more susceptible to defoliation interval.
Conclusion
Overall, results from the present study indicated that the longest defoliation interval (four-leaf stage) resulted in high cumulative DM yield and adequate botanical composition and tiller density; however, herbage quality was compromised compared with shorter defoliation intervals (one- and two-leaf stages). The present study highlighted that under field conditions, TF is more susceptible to defoliation interval than PR. Under the range of defoliation treatments applied (one-, two- or four-leaf stages), it was not possible to achieve an adequate compromise between DM yield, botanical composition, tiller survival and herbage quality in TF, and thus, defoliation at the three-leaf stage requires further investigation.
Acknowledgements
The authors would also like to thank and acknowledge the funding received from the Colin Holmes Dairy Scholarship. Particular thanks to Massey staff involved in the setting up and running of this study.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Allo AV, Southon T. 1967. Experiences with tall fescue. Proceedings of the New Zealand Grassland Association. 29:128–134.
- Bell CC. 1985. Effect of defoliation frequency on simulated swards of ryegrass, tall fescue, and a 50/50 mixture of the 2 species. New Zealand Journal of Agricultural Research. 28:307–312. doi: 10.1080/00288233.1985.10430432
- Brougham RW. 1957. Pasture growth rate studies in relation to grazing management. Proceedings of the New Zealand Society of Animal Production. 17:46–55.
- Brougham RW. 1961. Some factors affecting the persistency of short-rotation ryegrass. New Zealand Journal of Agricultural Research. 4:516–522. doi: 10.1080/00288233.1961.10431609
- Charlton JFL, Stewart AV. 1999. Pasture species and cultivars used in New Zealand – a list. Proceedings of the New Zealand Grassland Association. 61:147–166.
- Clark CEF, Clark DA, Waugh CD, Roach CG, Glassey CB, Woodward SL, Minneé EMK, Woodfield DR. 2010. Systems to increase grazeable forage production in the Waikato: a progress report on the tall fescue and perennial ryegrass components of these systems. Proceedings of the New Zealand Grassland Association. 72:49–54.
- Donaghy DJ, Turner LR, Adamczewski KA. 2008. Effect of defoliation management on water-soluble carbohydrate energy reserves, dry matter yields, and herbage quality of tall fescue. Agronomy Journal. 100:122–127. doi: 10.2134/agronj2007.0016
- Easton HS, Pennell CG. 1993. Breeding tall fescue for establishment vigour. Proceedings of the New Zealand Grassland Association. 55:151–154.
- Fulkerson WJ. 1994. Effect of redefoliation on the regrowth and water soluble carbohydrate content of Lolium perenne. Australian Journal of Agricultural Research. 45:1809–1815. doi: 10.1071/AR9941809
- Fulkerson WJ, Donaghy DJ. 2001. Plant-soluble carbohydrate reserves and senescence – key criteria for developing an effective grazing management system for ryegrass-based pastures: a review. Australian Journal of Experimental Agriculture. 41:261–275. doi: 10.1071/EA00062
- Fulkerson WJ, Slack K, Hennessy DW, Hough GM. 1998. Nutrients in ryegrass (Lolium spp.), white clover (Trifolium repens) and kikuyu (Pennisetum clandestinum) pastures in relation to season and stage of regrowth in a subtropical environment. Australian Journal of Experimental Agriculture. 38:227–240. doi: 10.1071/EA97161
- Hart RH, Carlson GE, McCloud DE. 1971. Cumulative effects of cutting management on forage yields and tiller densities of TF and orchardgrass. Agronomy Journal. 63:895–898. doi: 10.2134/agronj1971.00021962006300060022x
- Hendriks SJ, Donaghy J, Matthew C, Bretherton MR, Sneddon NW, Cosgrove GP, Christensen CL, Kaufononga S, Howes J, Osborne MA, et al. 2016. Dry matter yield, nutritive value and tiller density of tall fescue and perennial ryegrass swards under grazing. Journal of New Zealand Grassland. 78:149–156.
- Hunt WF, Easton HS. 1989. Fifty years of ryegrass research in New Zealand. Proceedings of the New Zealand Grassland Association. 50:1–23.
- Hunt WF, Field TF. 1976. Growth characteristics of perennial ryegrass. Proceedings of the New Zealand Grassland Association. 40:104–113.
- Jewiss OR. 1972. Tillering in grasses – its significance and control. Journal of the British Grassland Society. 27:65–81. doi: 10.1111/j.1365-2494.1972.tb00689.x
- Kays S, Harper JL. 1974. The regulation of plant and tiller density in a grass sward. Journal of Ecology. 62:97–105. doi: 10.2307/2258882
- Kemp PD, Tavakoli H, Hodgson J. 2001. Crop Physiology and Management: physiological and morphological responses of tall fescue and perennial ryegrass to leaf defoliation. 10th Australian Agronomy Conference, Jan 29–Feb 1; Hobart, Australia.
- Kolver ES. 2000. Nutritional guidelines for the high producing dairy cows. Proceedings of the Ruakura Farmers’ Conference. 51:78–87.
- Litherland AJ, Woodward SJR, Stevens DR, McDougal DB, Boom CJ, Knight TL, Lambert MG. 2002. Seasonal variations in pasture quality on New Zealand sheep and beef farms. Proceedings of the New Zealand Society of Animal Production. 62:138–142.
- Lowe KF, Bowdler TM, Casey ND, Moss RJ. 1999. Performance of temperate perennial pastures in the Australian subtropics. 1. Yield, persistence and pasture quality. Australian Journal of Experimental Agriculture. 39:663–676. doi: 10.1071/EA98021
- Matthew C, Agnusdei MG, Assuero SG, Sbrissia AF, Scheneiter O, Silva SCD. 2013. State of knowledge in tiller dynamics. Proceedings of the 22nd International Grasslands Congress. 1041–1043.
- McCallum DA, Thomson NA, Thom ER. 1992. The place of tall fescue in intensive dairying. Proceedings of the Ruakura Farmer’s Conference. 44:93–97.
- Milne GD, Shaw R, Powell R, Pirie B, Pirie J. 1997. Tall fescue use on dairy farms. Proceedings of the New Zealand Grassland Association. 59:163–167.
- Neal JS, Fulkerson WJ, Lawrie R, Barchia IM. 2009. Difference in yield and persistence among perennial forages used by the dairy industry under optimum and deficit irrigation. Crop and Pasture Science. 60:1071–1087. doi: 10.1071/CP09059
- Nie ZN, Chapman DF, Tharmaraj J, Clements R. 2004. Effects of pasture species mixture, management, and environment on the productivity and persistence of dairy pastures in south-west Victoria. 2. Plant population density and persistence. Australian Journal of Agricultural Research. 55:637–643. doi: 10.1071/AR03175
- Roughan PG, Holland R. 1977. Predicting in-vivo digestibilities of herbages by exhaustive enzymic hydrolysis of cell walls. Journal of the Science of Food and Agriculture. 28:1057–1064. doi: 10.1002/jsfa.2740281204
- Tavakoli H. 1993. Physiological and morphological responses of TF (Festuca arundinacea Schreb.) and perennial ryegrass (Lolium perenne L.) to defoliation [unpublished PhD thesis]. Palmerston North, New Zealand: Massey University. 54 p.
- Tozer KN, Bourdot GW, Edwards GR. 2011. What factors lead to poor pasture persistence and weed ingress. Proceedings of the New Zealand Grassland Association. 15:129–138.
- Turner LR, Donaghy DJ, Lane PA, Rawnsley RP. 2006. Effect of defoliation management, based on leaf stage, on perennial ryegrass (Lolium perenne L.), prairie grass (Bromus willdenowii Kunth.) and cocksfoot (Dactylis glomerata L.) under dryland conditions. 1. Regrowth, tillering and water-soluble carbohydrate concentration. Grass and Forage Science. 61:164–174. doi: 10.1111/j.1365-2494.2006.00523.x
- Zarrough KM, Nelson CJ, Coutts JH. 1983. Relationship between tillering and forage yield of tall fescue. I. Yield. Crop Science. 23:333–337. doi: 10.2135/cropsci1983.0011183X002300020036x