
ABSTRACT
Agriculture is becoming increasingly intensified through irrigation, increased nitrogen inputs and a change in grazing animal from sheep to cattle. These changes have the potential to affect earthworms, an important component of the soil biology. Over 4 years, this study compared earthworm populations under sheep or cattle grazing, mowing or no harvesting of pasture and with or without combinations of nitrogen fertiliser and irrigation. Irrigation had the greatest influence on both pasture production and earthworm abundance, also changing the earthworm species composition, with epigeic Lumbricus rubellus becoming more abundant. Nitrogen fertiliser increased pasture production under irrigation in both the sheep grazed and mown treatments, but not under cattle grazed treatments. The opposing influences of increasing pasture production and physical pressures mean that moving from dryland sheep to irrigated cattle grazing may have a minimal overall effect on earthworm numbers, while irrigated sheep grazing provides an ideal earthworm habitat.
Introduction
Land use intensification of pastoral grazing systems has occurred in many regions of New Zealand (Williams Citation2004; MacLeod and Moller Citation2006). In summer-dry regions, this intensification has often been achieved by the installation of irrigation systems and accompanied by increased nitrogen fertiliser inputs and a change of grazing animal from sheep to cattle. All of these changes have the potential to affect the capacity of the soil to function and hence impact the soil quality (Herrick Citation2000). New Zealand soil types vary greatly in their resilience, with some soils being quite susceptible to damage from increased intensification pressures (Hewitt and Shepherd Citation1997).
The importance of earthworms in maintaining soil quality in agricultural soils has been widely documented (Edwards and Bohlen Citation1996). Earthworms provide benefits such as organic matter decomposition, mixing of soil and improved soil physical properties (Springett Citation1983; Lee Citation1985; Francis and Fraser Citation1998). While intensification, such as the increase in nitrogen application (Edwards and Lofty Citation1982), stimulates organic matter inputs into the soil food web to enhance earthworm populations (Curry et al. Citation2008), intensification can also reduce the size and diversity of earthworm populations as earthworm habitats become compromised (Fraser et al. Citation1996). It is often difficult to determine which factors are most important in driving changes within earthworm populations, with field studies restricted to only one or two factors of intensification. For example, while earthworms were found to be more abundant under higher sheep stocking rates (Schon et al. Citation2008), they declined under more intensive cattle grazing (Schon et al. Citation2011). Irrigation too has the ability to stimulate earthworm populations during the drier summer with little differences in earthworm populations during winter observed (Fraser et al. Citation2012).
This study examined the effects of sheep and cattle grazing, mown and undefoliated pasture and the interaction of these defoliation regimes with and without irrigation and nitrogen over 4 years on earthworm population size and biomass. The aim was to investigate land-use change impacts on earthworm populations and the potential implications of these changes. We hypothesise that the greatest impact or intensification will be from a change in animal grazing as the soil physical structure becomes compromised.
Materials and methods
The experiment was located on a flat site on a commercial sheep, beef and dairy grazing farm in North Otago, New Zealand (45.02 S, 170.76 E). The experimental design and treatments are fully explained in White and Knight (Citation2007). Briefly, the experiment was established in 2004 as a split–split plot design on a Timaru silt loam (Pallic soil, NZSC; Hewitt Citation2010). Twenty-four main plots were randomly allocated factorial combinations of four types of pasture defoliation regimes (sheep, cattle, mowing, no harvest/undefoliated), two sown pasture species (perennial ryegrass, Lolium perenne cv. Samson with the AR1 endophyte and tall fescue, Festuca arundinacea cv. Advance with the MaxP endophyte) and two levels of irrigation (with and without irrigation).
The ryegrass and tall fescue were direct drilled into barley stubble in March 2004 and lightly grazed before treatment applications began in January 2005. The main plot factorial combinations were replicated two times. Each main plot was divided into two split plots, each 10 × 16.6 m in area, giving a total of 48 split plots. These split plots pairs were fenced individually—one of the pair received nitrogen fertiliser, the other did not. Split plots allocated to the mowing treatment were further divided into two halves, one half being mown and the other left undefoliated.
When above ground biomass reached 4000 kg DM/ha, the mown plots were harvested to 50 mm above ground level and the herbage removed from plots. The pasture mass on grazed treatments was determined pre- and post-grazing using a capacitance pasture probe, and the differences summed to calculate the total annual herbage harvested. Sheep grazed at greater than or equal to 2500 kg DM/ha down to 1000 kg DM/ha over a 24–36-hour period. Growing lambs were typically used. Cattle grazed at greater than or equal to 3000 kg DM/ha down to 1500 kg DM/ha over a 6–8-hour period. Mature dairy cows were typically used. Pasture composition was assessed by taking 10 random pasture samples in each paddock three to four times per year. Herbage material was subsampled, hand sorted to species, dried and weighed.
Irrigation was applied using impact sprinklers with the timing aligned to that occurring on the commercial farm schedule. Approximately 25 mm of water was applied during each irrigation event and applied once every 7–14 days from October to April, which resulted in average volumetric soil moisture of 27% for irrigated and 17% for dryland paddocks. The nitrogen application rate differed between treatments as the aim was to apply sufficient N to remove any limitation to pasture growth but without causing excessive leaching losses as determined by the biophysical simulation model EcoMod (White and Knight Citation2007). This meant applying different amounts to different harvest and irrigation treatments; specifically, 50, 75, 200, 125, 150 and 600 kg N/ha/year to the dryland grazed, dryland undefoliated, dryland mown, irrigated undefoliated, irrigated grazed and irrigated mown treatments, respectively.
Soil sampling for earthworms occurred each winter from 2006 to 2009 (28 June 2006, 9 July 2007, 15 August 2008, 21 July 2009). Eight 100 mm wide × 200 mm deep round cores were taken from each split plot in 2006, with four samples being collected from each of the mown and undefoliated treatments. Four square samples (140 mm wide × 200 mm deep) were taken from each split plot in 2007, 2008 and 2009. Earthworms were manually extracted from the soil and left in water overnight to remove all soil from their gut. They were then identified to species (Martin Citation1977), counted and subsequently dried using paper towels and weighed. Soil pH was measured in 2007 and again in 2009 using 1:2.1 v/v water slurry. Soil porosity was assessed from cores (100 mm diameter, 50 mm deep) using volumetric soil water content (Danielson and Sutherland Citation1986). Bulk density was determined by oven drying at 105°C for 48 hours and weighing. Total porosity (v/v) was calculated using bulk density data and assuming a particle density of 2.65 g/cm3. Soil pores <30 μm (microporosity), 30–300 μm and >300 μm (macroporosity) were distinguished.
Data in each year were analysed and compared separately from other years, using a split-plot analysis of variance (ANOVA). Each split-plot ANOVA consisted of full factorial combinations of two whole plot factors: defoliation treatment (three levels: cattle, sheep or mown) and irrigation (two levels: irrigated or dry), and one sub-plot factor, nitrogen (two levels: nitrogen or no nitrogen).
Data values from all 4 years were analysed and compared using another split-plot ANOVA, which consisted of not only the same terms as the ANOVA for within year comparison but also an additional factor, year, for blocking whole plot. Earthworm number was loge(n + 1) transformed prior to the split-plot ANOVA, in order to meet ANOVA’s assumption of normally distributed data. All split-plot ANOVAs were carried out with statistical software Genstat version 18. There was no statistically significant influence of pasture species on earthworms, so data are presented as the mean of the ryegrass and fescue plots.
Results
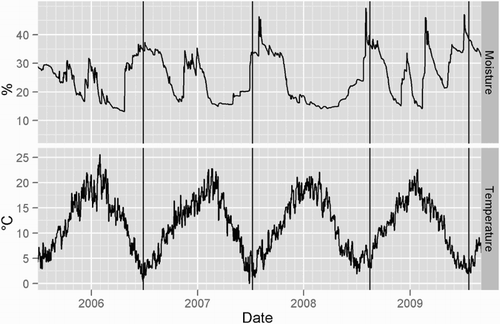
overlays the sampling dates, daily 10 cm soil temperature and dryland soil moisture content from the nearby Windsor climate station. Earthworm populations were sampled during the winter. In 2006, 2007 and 2009, sampling occurred within 11 days of the coldest day of the year, and 35 days after the coldest day in 2008. The annual rainfall was 523, 445, 486 and 692 mm for each respective year (starting 1 July 2005 to 30 June 2006) prior to sampling. In 2008, there was a particularly dry summer (December and January), while the 2009 summer was wetter than normal. In all years the soil had reached field capacity prior to earthworm sampling.
Figure 1. Soil moisture content without irrigation (vol %) and 10 cm soil temperature (°C) over the experimental period starting July 2005. Data collected from the Windsor climate station (www.cliflo.niwa.co.nz). Vertical lines denote earthworm sampling dates.
Average annual pasture production ranged from 8440 to 9650 kg DM/ha across the 4 years and the differences between treatments were consistent. Pasture production was greater under irrigated than dryland plots ( and ). Nitrogen application increased pasture production under irrigated treatments in both the mown and sheep grazed pastures but not the cattle grazed pastures. Pasture production was greatest in the irrigated, N fertilised and sheep grazed pastures and was also high under mown, irrigated and fertilised plots. Clover content was lower with N application, and this was significant for mown treatments, being higher under irrigation (). The undefoliated treatment had very low levels of white clover present.
Figure 2. Average annual pasture production (from 2006 to 2009) for the different defoliation treatments under dryland (striped bar) or irrigated (solid bar), receiving nitrogen (▪) or no nitrogen (▪). Error bars represent 1 SEM. a and b indicate significant irrigation difference for each nitrogen and grazing combination. Significant differences with nitrogen application were found under irrigated sheep, dry mown and irrigated mown treatments.
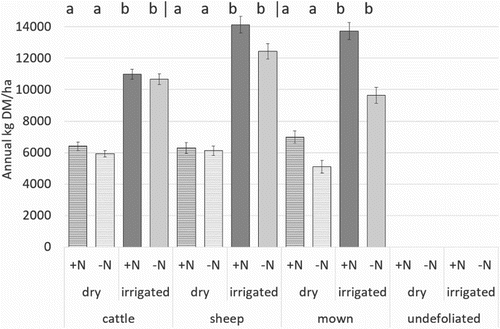
Table 1. ANOVA table for pasture production across years.
Table 2. Clover content (%) in all the treatments from July 2006 to June 2009 (mean ± SEM).
Soil pH in 2009 was significantly higher in nitrogen fertilised plots compared to those receiving no nitrogen (average 5.6 vs 5.3, respectively). The mown treatments had the lowest soil pH, dropping to 4.8 with N application, whereas other treatments receiving N fertiliser had an average soil pH of 5.2–5.6 ().
Table 3. Soil pH (1:2.1 v/v water slurry) in all the treatments in 2009 (mean ± SEM).
Total porosity in the final year (2009) was greater in the surface 0–5 cm (range 58.6–63.5%) compared to 5–10 cm depths (range 51.7–54.7%). Micropores made up the majority of porosity with microporosities ranging from 37% to 42%. In the surface 0–5 cm, there were few treatment effects on soil porosity ( and ), although the mown plots had a higher porosity under dryland than irrigated pastures when nitrogen was applied. Macroporosity was lowest under the undefoliated treatment at 0–5 cm depths, this was significant for dry mown plots in comparison to dry undefoliated plots receiving nitrogen application at 0–5 cm. At 5–10 cm depths, soil porosity and micropores were lowest under cattle grazed pastures with nitrogen and irrigation, micropores were also lower in dryland plots without nitrogen application when grazed by cattle than sheep or mown. Total soil porosity at 5–10 cm depths was also lower in undefoliated pastures in comparison to mown pastures receiving irrigation and no nitrogen.
Figure 3. Total soil porosity in 2009 at A, 0–5 cm and B, 5–10 cm depths for the different defoliation treatments under dryland or irrigated, receiving nitrogen or no nitrogen. Soil pores <30 μm (▪), 30–300 μm (□) and >300 μm (▪). Error bars represent 1 SEM for total soil porosity. a and b indicate significant irrigation difference for each nitrogen and grazing combination. Significant differences with nitrogen application were found for total porosity under mown irrigated plots at 0–5 cm depths.
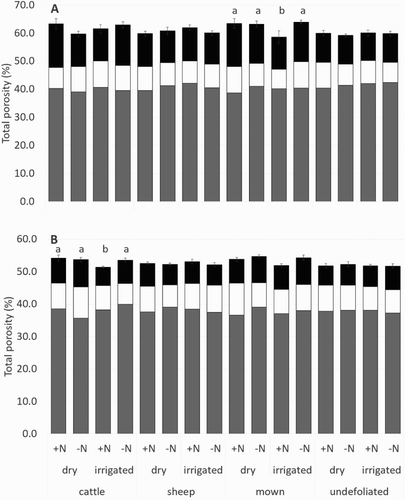
Table 4. ANOVA table for total porosity in 2009.
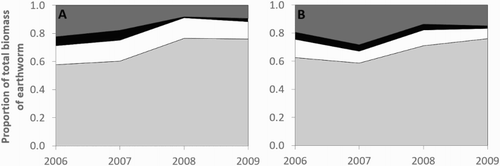
The four earthworm species identified were the endogeic Aporrectodea caliginosa (Savigny), Aporrectodea trapezoides (Duges) and Octolasion cyaneum (Savigny), and the epigeic Lumbricus rubellus (Hoffmeister). The dominant species under all treatments was A. caliginosa (). L. rubellus contributed a higher proportion of the population with irrigation (p = .016).
Figure 4. The proportional contribution of each species to the total mass of earthworms (m2) sampled in each year, under A, dryland and B, irrigation. Earthworm species include L. rubellus (▪), O. cyaneum (▪), A. trapezoides (□) and A. caliginosa (▪).
Irrigation had a larger influence on earthworm abundance and biomass than the nitrogen treatment ( and ). Total earthworm abundance was greater in irrigated than dryland plots, reflecting the dominant species A. caliginosa and L. rubellus. In contrast, O. cyaneum and A. trapezoides were less abundant and were less influenced by treatment. Although earthworm abundance varied throughout the years, their abundance was consistently greater under irrigated sheep pastures (). The addition of nitrogen in irrigated plots reduced earthworm abundance in the mown and cattle treatments, particularly with respect to A. caliginosa ().
Figure 5. Average earthworm A, abundance and B, biomass for each treatment across the 4 years sampled (winter 2006–2009). Earthworm species include L. rubellus (□), O. cyaneum (▪), A. trapezoides (▪) and A. caliginosa (▪). Error bars represent 1 SEM for total earthworm abundance or biomass. a and b indicate significant irrigation difference for each nitrogen and grazing combination. Significant differences with nitrogen application were found under irrigated cattle for earthworm abundance and irrigated mown plots for earthworm abundance and biomass.
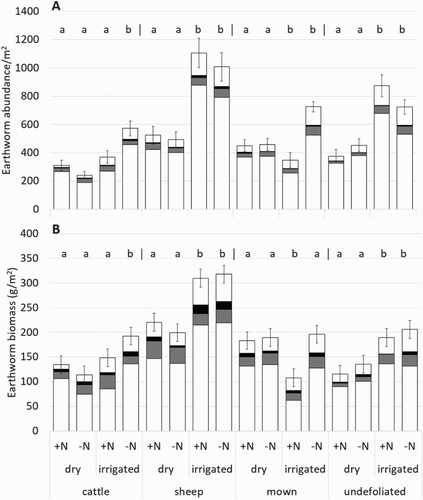
Figure 6. Average total earthworm abundance under A, dry and B, irrigated pastures, and average earthworm biomass under C, dry and D, irrigated pastures across all years sampled. Harvest type includes cattle (![]()
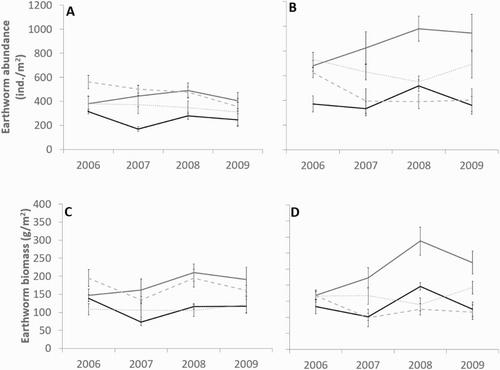
Table 5. ANOVA table for earthworm abundance and biomass (m2) across years.
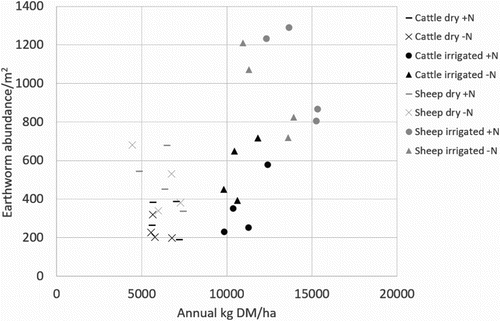
There was a strong positive relationship between annual earthworm populations and pasture production (). Differences in pasture production reflect treatment differences, with higher pasture production under irrigation than dryland, and also under sheep grazing compared to cattle grazing.
Figure 7. Relationship between earthworm abundance and pasture production in the different treatments.
Discussion
Intensification of pastures involves increasing inputs into the system in order to increase productivity. In this study, these inputs were water (irrigation), nutrients (fertiliser) and a change in livestock. Contrary to our hypothesis, irrigation had the greatest influence on both pasture production and earthworm abundance in our study, while the other practices of N application and defoliation regimes caused more subtle changes to the soil physical and chemical environment that influenced earthworms. By reducing soil moisture limitations on pasture growth, irrigation increased herbage yield in all recorded treatments. This corresponded to higher earthworm abundances under irrigation than dryland pastures during the winter period, not just during the drier summer as reported by Fraser et al. (Citation2012). The greatest pasture production was recorded under irrigated, sheep grazed pastures, which also had the largest earthworm populations as organic matter inputs were increased to the soil food web (Curry et al. Citation2008).
Mown, irrigated and fertilised treatments also had high pasture production. In the mown treatments where all physical pressures were removed, earthworm populations were not stimulated as herbage litter was removed from the plots, preventing the increases in yield under irrigation to be incorporated into earthworm populations. Similar results were also observed by Schon et al. (Citation2010) under a cut and carry system. If the litter had been returned to the mown treatment, we would have expected earthworms to increase as observed by Watkin and Wheeler (Citation1966), especially considering the high percentage of clover in these systems (van Eekeren et al. Citation2010). The addition of N in the mown treatments also depressed earthworm populations further. Although N application does not necessarily decrease earthworm abundance, soil acidification can be detrimental to earthworms (Edwards and Lofty Citation1982; Curry et al. Citation2008), which prefer a pH of 6–7 (Fraser Citation1994). The mown and irrigated treatment in this study received a high N application (600 kg N/ha), and without application of lime, the pH dropped to 4.8 over the course of the experiment compared to all the other treatments receiving smaller amounts of N where the pH remained between 5.2 and 5.6. A drop in pH to below 5 appears to limit earthworm populations in this study.
As typical of New Zealand pastures (Fraser et al. Citation1996), A. caliginosa was the most common earthworm in this study. Changes in A. caliginosa abundance were reflected in differences in total earthworm abundance. Hence, this species benefited from irrigation. However, irrigation also altered the earthworm community, with epigeic L. rubellus becoming more prevalent under irrigation. An increase in this epigeic species was also observed by Lobry de Bruyn and Kingston (Citation1997), presumably since moisture fluctuations are reduced within the surface soil under irrigation and there may be a greater accumulation of surface litter.
The addition of N with irrigation increased pasture growth under all defoliation treatments, except cattle grazing. Under irrigated and fertilised, cattle grazed pastures macroporosities were lower at 5–10 cm depths (5.7%) than 0–5 cm depths (11.5%). Although differences in macroporosity between irrigated cattle and dryland sheep were not as pronounced as those reported by Houlbrooke et al. (Citation2011), structural degradation has implications when intensifying from sheep to cattle grazing, and these impacts remain evident at deeper soil depths (Houlbrooke et al. Citation2011). The higher treading pressures of cattle (c. 138 kPa), which are more than double that of sheep (c. 66 kPa) (Greenwood and McKenzie Citation2001), compact the physical environment and impact both pasture growth and the habitat for earthworms to function in (Kretzschmar Citation1991). While earthworms are able to adjust to their environment, ingesting more soil and reducing the impact of compaction (Greenwood and McKenzie Citation2001; Jouquet et al. Citation2012), depending on the severity and frequency of compaction, will determine how quickly the soil and earthworm populations are able to recover from compaction. In this study, the increase in physical degradation and restricted pasture growth under irrigated, fertilised, cattle grazed pastures resulted in lower earthworm populations.
Soil macroporosity was lower under the undefoliated treatments in comparison to the mown treatment which also received no physical pressure from grazing. This observation is opposite to that observed after 3 years by Roach and Morton (Citation2005). In the current study, earthworm abundances between mown and undefoliated treatments were similar (with the exception of irrigated fertilised plots). Mown treatments received more than double the application of nitrogen than the undefoliated treatments. Although pasture production was not measured under the undefoliated treatments, it is likely root growth and turnover would have been higher under the mown treatment, leading to a greater influence of plant roots to soil macroporosity (Beven and Germann Citation1982). It may be difficult to distinguish between macropores produced by plant roots or earthworms as both structures are of tubular nature (Beven and Germann Citation1982). Although macropores created by earthworms are typically 2.5–3 mm diameter (Springett Citation1983), and larger than ryegrass root diameter of 0.25 mm (Nichols and Crush Citation2007), the clover tap root is a similar size to the earthworm burrow at 2.8 mm diameter (Caradus and Woodfield Citation1998). With clover content being lower in undefoliated pastures than mown pastures, this suggests that the coarse clover tap roots may play an important role in macropore formation as also observed by Bodner et al. (Citation2014).
The relationship between earthworms and pasture production under the different managements highlights the importance of organic matter inputs in stimulating earthworm populations (Curry et al. Citation2008). The presence of earthworms in soils has been reported to increase pasture production by 20% (van Groenigen et al. Citation2014), and hence stimulating pasture production should simultaneously provide positive feedback to pasture production. However, as the physical pressures from cattle grazing limit earthworm populations, this has the potential to reduce the influence of earthworms on pasture production. Treading pressures of cattle have been reported to limit earthworm populations in other studies (Schon et al. Citation2012), although is not always observed (Norris et al. in preparation), this may be a result of differences in soil type with some soil types more vulnerable to compaction than others (Hewitt and Shepherd Citation1997). Understanding the level of grazing on particular soils that limit earthworm populations needs to be determined.
Irrigation had the greatest effect on earthworm populations but nitrogen application and harvest method (including grazing) also had varying effects. In order to maximise the benefits provided by earthworms in terms of ecosystem services, including enhancing pasture production, we need management practices which enhance dry matter production while minimising soil physical degradation. While irrigated sheep grazing provides an ideal environment for earthworms, intensifying pasture land by converting dryland sheep to irrigated cattle grazing may have a minimal overall effect on earthworm abundance.
Acknowledgements
We gratefully acknowledge Grant and Elle Ludemann for providing the land and Duncan Kingan and Sandy Harper who managed the grazing and took pasture probe measurements. Cath Goulter provided technical help with experiments. David Baird provided the design and statistical analysis.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Beven K, Germann P. 1982. Macropores and water flow in soils. Water Resources Research. 18:1311–1325. doi: 10.1029/WR018i005p01311
- Bodner G, Leitner D, Kaul H-P. 2014. Coarse and fine root plants affect pore size distributions differently. Plant and Soil. 380:133–151. doi: 10.1007/s11104-014-2079-8
- Caradus JR, Woodfield DR. 1998. Genetic control of adaptive root characteristics in white clover. Plant and Soil. 200:63–69. doi: 10.1023/A:1004296707631
- Curry JP, Doherty P, Purvis G, Schmidt O. 2008. Relationships between earthworm populations and management intensity in cattle-grazed pastures in Ireland. Applied Soil Ecology. 39:58–64. doi: 10.1016/j.apsoil.2007.11.005
- Danielson RE, Sutherland PL. 1986. Porosity. In: Methods of soil analysis: physical and mineralogical methods, volume 1. Madison (WI): American Society of Agronomy; p. 443–460.
- Edwards CA, Bohlen PJ. 1996. Biology and ecology of earthworms. London (UK): Chapman & Hall.
- Edwards CA, Lofty JR. 1982. Nitrogenous fertilizers and earthworm populations in agricultural soils. Soil Biology & Biochemistry. 14:515–521. doi: 10.1016/0038-0717(82)90112-2
- Francis GS, Fraser PM. 1998. The effects of three earthworm species on soil macroporosity and hydraulic conductivity. Applied Soil Ecology. 10:11–19. doi: 10.1016/S0929-1393(98)00045-6
- Fraser PM. 1994. Effects of management practices on earthworm populations. In: Soil biota: management in sustainable farming systems. Melbourne: CSIRO; p. 125–134.
- Fraser PM, Schon NL, Piercy JE, Mackay AD. 2012. Influence of summer irrigation on soil invertebrate populations in a long-term sheep irrigation trial at Winchmore (Canterbury). New Zealand Journal of Agricultural Research. 55: 165–180. doi: 10.1080/00288233.2012.662902
- Fraser PM, Williams PH, Haynes RJ. 1996. Earthworm species, population size and biomass under different cropping systems across the Canterbury plains, New Zealand. Applied Soil Ecology. 3:49–57. doi: 10.1016/0929-1393(95)00062-3
- Greenwood KL, McKenzie BM. 2001. Grazing effects on soil physical properties and the consequences for pastures: a review. Australian Journal of Experimental Agriculture. 41:1231–1250. doi: 10.1071/EA00102
- Herrick JE. 2000. Soil quality: an indicator of sustainable land management? Applied Soil Ecology. 15:75–83. doi: 10.1016/S0929-1393(00)00073-1
- Hewitt AE. 2010. New Zealand Soil Classification. Landcare Research Science Series No. 1, Third Edition. Manaaki Whenua Press.
- Hewitt AE, Shepherd TG. 1997. Structural vulnerability of New Zealand soils. Australian Journal of Soil Research. 35:461–474. doi: 10.1071/S96074
- Houlbrooke DJ, Paton RJ, Littlejohn RP, Morton JD. 2011. Land-use intensification in New Zealand: effects on soil properties and pasture production. Journal of Agricultural Science. 149:337–349. doi: 10.1017/S0021859610000821
- Jouquet P, Huchet G, Bottinelli N, Thu TD, Duc TT. 2012. Does the influence of earthworms on water infiltration, nitrogen leaching and soil respiration depend on the initial soil bulk density? A mesocosm experiment with the endogeic species Metaphire posthuma. Biology and Fertility of Soils. 48:561–567. doi: 10.1007/s00374-011-0652-3
- Kretzschmar A. 1991. Burrowing ability of the earthworm Aporrectodea-longa limited by soil compaction and water potential. Biology and Fertility of Soils. 11:48–51. doi: 10.1007/BF00335834
- Lee K. 1985. Earthworms: their ecology and relationships with soils and land use. Sydney: Academic Press.
- Lobry de Bruyn LA, Kingston TJ. 1997. Effects of summer irrigation and trampling in dairy pastures on soil physical properties and earthworm number and species composition. Australian Journal of Agricultural Research. 48:1059–1079. doi: 10.1071/A94132
- MacLeod C, Moller H. 2006. Intensification and diversification of New Zealand agriculture since 1960: an evaluation of current indicators of land use change. Agriculture, Ecosystems and Environment. 115:201–218. doi: 10.1016/j.agee.2006.01.003
- Martin NA. 1977. Guide to lumbricid earthworms of New Zealand pastures. New Zealand Journal of Experimental Agriculture. 5:301–309. doi: 10.1080/03015521.1977.10425985
- Nichols SN, Crush JR. 2007. Selecting forage grasses for improved nitrate retention – a progress report. Proceedings of the New Zealand Grassland Association. 207–211.
- Norris T, Guernion M, Schon NL, Pronger J, Wall A, Sparling G, Schipper LA. in preparation. A comparison of earthworm abundance in adjacent paired pasture sites used for dairy or drystock farming, at farm and regional scale in Waikato, New Zealand.
- Roach CG, Morton JD. 2005. Impacts of intensive dairying on soil and pasture sustainability in Taranaki. Proceedings of the New Zealand Grassland Association. 67:109–116.
- Schon NL, Mackay AD, Minor MA. 2011. Effects of dairy cow treading pressures and food resources on invertebrates in two contrasting and co-occurring soils. Soil Research. 49:703–714. doi: 10.1071/SR11119
- Schon NL, Mackay AD, Minor MA. 2012. Relationship between food resource, soil physical condition and invertebrates in pastoral soils. Soil Science Society of America Journal. 76:1644–1654. doi: 10.2136/sssaj2011.0375
- Schon NL, Mackay AD, Minor MA, Yeates GW, Hedley MJ. 2008. Soil fauna in grazed New Zealand hill country pastures at two management intensities. Applied Soil Ecology. 40:218–228. doi: 10.1016/j.apsoil.2008.04.007
- Schon NL, Mackay AD, Yeates GW, Minor MA. 2010. Separating the effects of defoliation and dairy cow treading pressure on the abundance and diversity of soil invertebrates in pastures. Applied Soil Ecology. 46:209–221. doi: 10.1016/j.apsoil.2010.08.011
- Springett JA. 1983. Effect of five species of earthworm on some soil properties. Journal of Applied Ecology. 20:865–872. doi: 10.2307/2403131
- van Eekeren N, Bokhorst J, Brussaard L. 2010. Roots and earthworms under grass, clover and a grass-clover mixture. 9th World Congress of Soil Science, Soil Solutions for a Changing World; Brisbane, Australia.
- van Groenigen JW, Lubbers IM, Vos HMJ, Brown GG, De Deyn GB, van Groenigen KJ. 2014. Earthworms increase plant production: a meta-analysis. Scientific Reports. 4:1–7.
- Watkin BB, Wheeler JL. 1966. Some factors affecting earthworm populations under pasture. Grass and Forage Science. 21:14–20. doi: 10.1111/j.1365-2494.1966.tb00441.x
- White TA, Knight TL. 2007. Pasture yield and composition under different land uses in North Otago Rolling Downlands. Proceedings of the New Zealand Grassland Association. 69:65–71.
- Williams JM. 2004. Growing for good: intensive farming, sustainability and New Zealand’s environment. Wellington, New Zealand: Parliamentary Commissioner for the Environment.