
ABSTRACT
This study determined the effect of feeding synthetic zeolite A prepartum to reduce periparturient hypocalcaemia risk on the eating, lying, and activity behaviours of multiparous grazing dairy cows. Animals were randomly allocated to either a Zeolite treatment group (n = 21; individually fed 500 g/d zeolite for two weeks prepartum) or a Control group (n = 22). Behaviour data obtained from accelerometers were analysed to determine the effects of treatment, parity (Parity 2−3 vs. 4+), and their interactions during 3 periods: PRE (−21 to −3 d), PERI (−2–2 d) and POST (3–28 d) relative to the day of calving (day 0). Parity 2–3 Zeolite cows ate for 18 min/day less than Parity 2–3 Control cows PRE. Zeolite-fed cows also ate for 24 min/day less than Control cows during the PERI period. There were no treatment differences in lying time or activity PRE or PERI, but Zeolite-fed cows spent more time lying POST. Regardless of treatment, Parity 2–3 cows were generally more active and ate for longer than Parity 4 + cows. Our results suggest zeolite may subtly decrease appetite or alter eating behaviour during the supplementation period, particularly in younger cows, and may improve cow comfort postcalving.
Introduction
Periparturient hypocalcemia is a common metabolic disorder that occurs in dairy cows when they fail to adapt to the sudden increased physiological demand for calcium (Ca) to support milk production after parturition. Blood Ca concentrations can decline below the normal physiological range of between 2.1 and 2.5 mmol/L (Goff Citation2008) and, in severe cases, clinical milk fever can occur where a lack of muscle contractility can lead to paresis and risk of mortality. Although only 2–5% of grazing dairy cows experience clinical hypocalcemia (Ca <1.4 mmol/L; McDougall Citation2001; Roche Citation2003), a far greater percentage experience subclinical hypocalcemia (approximately 30%–40% of cows with blood Ca <2.0 mmol/L, and 50% with blood Ca ≤2.14 mmol/L; Roche Citation2003; Roberts and McDougall Citation2019). Similar incidences of clinical and subclinical hypocalcemia have been reported in higher-yielding cows managed in housed systems (Wilkens et al. Citation2020). Furthermore, numerous studies in housed cows indicate associations between hypocalcemia and other periparturient health disorders such as mastitis (Goff Citation2008) and metritis (McArt and Neves Citation2020), or early removal from the herd (McArt and Neves Citation2020).
Supplementing between 500–1000 g/d of synthetic zeolite A for 2–4 weeks prepartum can effectively reduce the risk of hypocalcemia in both housed (Thilsing-Hansen and Jørgensen Citation2001; Grabherr et al. Citation2009; Kerwin et al. Citation2019) and grazing (Roche et al. Citation2018; Crookenden et al. Citation2020) dairy cows, likely due to its ability to bind dietary Ca and phosphorus (P) in the gut, thereby reducing their absorption and triggering homeostatic mechanisms (Thilsing-Hansen et al. Citation2002; Grabherr et al. Citation2009). Previous studies using zeolite in dairy cows have focused on its effects on blood mineral concentrations and, to a lesser extent, on milk yield and composition. Researchers conducting studies in housed systems have also noted reduced dry matter intakes (DMI) during the prepartum supplementation period, which they hypothesised to be related to zeolite’s poor palatability or zeolite-induced hypophosphatemia (Thilsing-Hansen et al. Citation2002; Grabherr et al. Citation2009). Due to the current lack of reliable measures of DMI in grazing cows, it is important that the relationship between zeolite and eating behaviours is investigated to contribute to understanding its effects and modes of action under different management systems.
Periparturient hypocalcemia without paresis has also been associated with longer lying times and reduced activity in grazing dairy cows (Hendriks et al., Citation2020). These behaviours are important indicators of cow welfare (Munksgaard and Simonsen Citation1996), and therefore, the effect of zeolite supplementation on lying and activity behaviours should be assessed. Given that parity is independently associated with both hypocalcemia risk and behaviour, whereby older cows are at increased risk of developing hypocalcemia (Roche and Berry Citation2006; Roberts and McDougall Citation2019) and are typically less active (Hendriks et al. Citation2019b), it is important that parity is also considered when investigating the effects of zeolite on behaviour.
We hypothesized that (1) grazing cows supplemented with synthetic zeolite A during the prepartum period would spend less time eating than non-supplemented control cows, and (2) peripartum lying times would be shorter, and activity greater, in zeolite-supplemented cows, particularly in older cows who are at greater risk of developing hypocalcemia. Therefore, the objective of the current study was to determine the effect of prepartum supplementation of synthetic zeolite A on the eating, lying and activity behaviours of multiparous grazing dairy cows during the transition period.
Materials and methods
Animal handling and experimental design
This experiment was conducted at Lye Farm, Hamilton, New Zealand (37°46′S, 175°18′E) between 21 June and 16 August 2016, with approval from the Ruakura Animal Ethics Committee (RAEC #13871). The experimental design has been previously described (Roche et al. Citation2018; Crookenden et al. Citation2020). Briefly, 50 predominantly Holstein-Friesian cows of mixed age (mean ± standard deviation (SD): 5.2 ± 1.8 years) and in late gestation were randomly allocated to 1 of 2 treatment groups (Control, n = 25; or Zeolite, n = 25). Treatment groups were balanced for expected calving date (range: 5–12 July 2016), age, previous milk production, genetic merit (New Zealand Breeding Worth; provided by Livestock Improvement Corporation Ltd., Hamilton, New Zealand), body condition score, and body weight before enrolment.
Cows were separated into two cohorts with staggered treatment start dates to achieve a target treatment duration of 14 d before their expected calving date (actual duration relative to calving; mean ± SD: Zeolite = 18.2 ± 3.6 d, Control = 20.6 ± 4.1 d). All cows were brought to a covered facility between approximately 0700–1000 h and individually supplemented with 5 kg (wet weight) of maize silage once daily, with the Zeolite treatment group receiving 500 g/cow per d of zeolite A (80% sodium aluminosilicate, synthetic embedded in starch; XZelit, Optimate MF+, Blue Pacific Minerals, Tokoroa, New Zealand) mixed into the maize silage. Supplementation with zeolite stopped at calving.
Cows grazed fresh pasture as a single herd both pre- and post-calving and were milked twice daily following parturition with an average (± standard error of the difference [SED]) energy-corrected milk yield of 25.0 (±1.12) kg/cow per d, and had a mean body weight and body condition score of 515.0 (± 4.59) kg and 4.55 (± 0.05) units, respectively. No differences (P > 0.05) were detected between treatment groups for these parameters, although Parity 4 + cows had greater (P < 0.05) mean energy-corrected milk yields than Parity 2–3 cows (26.2 vs 23.8 (± 1.03) kg/cow per d, respectively). After calving, cows were group supplemented with maize silage and pasture silage if target grazing residuals could not be met due to insufficient pasture (Crookenden et al. Citation2020). Consistent with standard farm practice, cows were supplemented with 80–100 g/cow per d magnesium by dusting pasture with magnesium oxide daily (Causmag; Causmag International, Young, NSW, Australia; 55% magnesium), and they received 200 g/cow per d of ground limestone (calcium carbonate) for the first 4 d after parturition.
Blood sampling and analyses
Blood samples were collected from each cow via coccygeal venepuncture immediately pre-treatment (mean ± SD: 19.4 ± 4.0 d precalving) and then on d −17 (± 3 d range) and −10 (± 3 d) precalving, the day of calving (d 0), and d 1, 4 (± 2 d), 7 (± 2 d), 14 (± 2 d), 21 (± 2 d), and 28 (± 2 d) postcalving. The handling of blood samples and laboratory analyses for plasma Ca concentrations were as described by Crookenden et al. (Citation2020).
Behavioural measurements
Lying time, lying bout duration, number of lying bouts and step count data were measured in 15-min intervals for each cow from 21 d precalving to 28 d postcalving using IceQube triaxial accelerometers (IceRobotics Ltd., Edinburgh, Scotland). These devices, described in detail by Hendriks et al. (Citation2019b), measure lying and activity behaviour through the position and movement of the hind leg through the three-dimensional space.
Cows were fitted with CowScout neck sensors (GEA, New Zealand) to measure eating time in 15-min intervals through recording the position and movement of the animal’s head. The use of CowScout neck sensors for measuring eating time has been validated for grazing cows using visual observations and is described in detail by Dela Rue et al. (Citation2020).
Of the 50 cows enrolled, 43 cows precalving and 42 cows postcalving were included in the final analysis. Of these animals, 22 were in the Control group (n = 11 of parity 2-3, and n = 11 of parity 4+) and 21 were supplemented with zeolite (n = 8 of parity 2-3, and n = 13 of parity 4+). Six cows were removed from analyses due to inaccessible files or incomplete data (>10 d of missing data), one cow removed due to missing treatment before calving, and another removed after inadvertently receiving 350 ml Calform Plus (≥10 to ≤50% calcium diformate; Bayer New Zealand Ltd., Auckland, NZ) for milk fever prevention without exhibiting clinical signs, which could influence her subsequent data. A further two cows in the CowScout dataset were excluded from the eating time behaviour analyses, as they lacked data needed as a pre-treatment covariate. Data were removed on particular days for individual cows if eating time was equal to 0 h (n = 6 observations), due to probable device recording failure as it is unlikely a cow will not eat within a 24-h period without any recorded clinical observations of illness.
For ease of data interpretation, the sampling dates for all data were assigned an experimental day relative to the date of calving (d 0). The day of calving for 51% of the cows was the recorded day of calf collection, which was once daily in the morning, with the day of calving altered for the remaining 49% of cows when the number of lying bouts suggested the actual calving date was the day before, using the criterion described by Hendriks et al. (Citation2020).
Statistical analyses
Data analysis was performed using SAS 9.4 (SAS Institute Inc., Cary, NC) with repeated-measures ANOVA to investigate the main and interactive effects of treatment (categorical: Control or Zeolite), parity (categorical: Parity 2–3 or 4+), and experimental day. The cohort treatment start date was included as an additional fixed effect, and cow included as a random effect in all analyses. All models were pairwise comparison adjusted using Tukey–Kramer and used a compound symmetry covariance structure unless stated otherwise. Least squares mean, SED, and Tukey-adjusted P-values are presented in the text.
Eating and lying time, the number and duration of lying bouts, and step count were summarised into 24-h periods for the analysis of behaviour across experimental days. Lying time and step count were also summarised into 4-h intervals (starting at 0200 h) for the analysis of behaviour within day. Behaviour data were categorised into three periods (PRE: −21 to −3 d precalving, PERI: −2 to +2 d, and POST: 3–28 d postcalving) and were analysed separately. The across- and within-day behaviour data measured during the 3 d before treatment start were averaged for each individual cow to create a pre-treatment covariate, included in the models as a fixed effect. Weather variables (daily rainfall, minimum air temperature at 0900 h, and maximum daily wind speed) retrieved from retrieved from The National Climate Database (NIWA, Citation2018) were included as fixed effects, as inclement weather is noted to affect the lying, activity and feeding behaviour of dairy cows (Schütz et al. Citation2010; Hendriks et al. Citation2019a).
For the analysis of behaviour across experimental days, large within- and across-day variation in behaviour existed between individual animals and it was determined that a 3-d moving average for all weather and behaviour variables during the PRE and POST periods should be used for analysis. However, raw daily data were used for the PERI period due to the known disruptive effects of the parturition event on cow behaviour (Hendriks et al. Citation2019b).
For the within-day analyses, intervals were averaged across days of the PRE and POST periods, while individual days during the PERI period were analysed separately and included weather variables on these days as fixed effects. Interval was the repeated measure, and heterogenous Toeplitz was identified as the appropriate covariance structure.
Results and discussion
The current study investigated the effects of zeolite on the behaviours of multiparous grazing dairy cows to test our hypotheses that eating time would be reduced during the period of prepartum supplementation, but the lying times would be shorter and activity greater in the zeolite-supplemented cows during the peripartum period due to reduced risk of hypocalcaemia.
The interactive effect of treatment and parity on eating time
In support of our first hypothesis, supplementing zeolite precalving altered eating time in grazing dairy cows, but these effects were only minor and differed with parity. There was a significant 3-way interactive effect of treatment, parity and day relative to calving (P = 0.01; ). Although this effect varied between days, in general the older (Parity 4+; mean ± SD 6.4 ± 1.44 years) cows spent a similar amount of time eating each day during the PRE period irrespective of whether they were fed zeolite (Control mean 6.9 h/d [range: 6.5–7.3 h/d] vs. Zeolite mean 6.9 h/d [range: 6.4–7.6 h/d]; ). The eating time of younger (Parity 2−3; mean ± SD 3.5 ± 0.51 years) cows fed zeolite ranged between 6.7 and 9.3 h/d (mean 7.6 h/d), but was generally less than Controls (mean 7.9 h/d, range: 7.4–8.5 h/d; ). It is unclear why this interaction with parity occurred. Nevertheless, all zeolite-supplemented cows had shorter eating times averaged across the PERI period (−0.4 h/d; ) when feeding behaviours varied widely and were disrupted by the calving process, in agreement with other studies (e.g. Schirmann et al. Citation2013; Neave et al. Citation2017). Within-day eating time was not affected by zeolite during PRE, PERI, or POST periods (all P > 0.30; Supplemental Table 1), and the effects of treatment on eating time did not extend into early lactation once the supplementation of zeolite stopped at calving (), consistent with previous studies in TMR-fed housed cows (Kerwin et al. Citation2019).
Figure 1. The effect of feeding zeolite precalving and parity on daily eating time during the transition period. Daily eating times (h/d) of the four treatment x parity groups (Control 2-3; Control 4+; Zeolite 2-3; Zeolite 4+) across −14 to 28 d relative to the day of calving (d 0) are presented. Days −21 to −15 precalving are not presented in the figure due to low cow numbers (≤ 10 per group) at these time points. The average pretreatment daily eating time for all cows (filled circle at d −15) was measured during the 2 d before treatment start (overall mean ± SD: −21.4 ± 4.6 d relative to calving). Error bars represent 1 × mean standard error of the difference. The overall treatment × parity × day interaction was significant during the PRE period (d −21 to −3; P = 0.01), and days when there was a treatment × parity interaction are represented by * (P < 0.05) and + (P < 0.15).
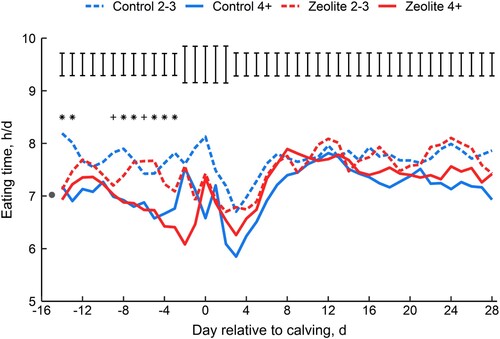
Table 1. The effect of feeding zeolite precalving and parity on eating, lying, and activity behaviours during the transition period. Least squares mean and standard error of the difference (SED) for the main effects of treatment (Control vs. Zeolite) and parity (2–3 vs. 4+) within the PRE (−21 to −3 d), PERI (−2 to 2 d), and POST (3–28 d) periods relative to the day of calving (d 0) are presented along with P-values for the effects of treatment (Treat), parity and day relative to calving (Day), and their 2- and 3-way interactions.
The slightly shorter eating times precalving may indicate a possible anorexic effect of zeolite in grazing cows that could result in small reductions in DMI. Unfortunately, this effect cannot be confirmed as DMI was not measured in the current study due to the complexities of obtaining accurate intakes for individual cows in a grazing system. Further, more efficient eating behaviour (e.g. greater bite rate) cannot be ruled out as a contributing factor. Nevertheless, the lack of any treatment effects on body condition score, body weight, milk production parameters, or plasma concentrations of non-esterified fatty acids or β-hydroxybutyrate (previously reported by Crookenden et al. (Citation2020) for a subset of cows (n = 10 per treatment) in the present study) indicates that if reductions in prepartum DMI did occur due to zeolite supplementation, they were relatively minor. The exact cause for reductions in DMI in zeolite-supplemented cows is currently unknown; however, previous researchers have hypothesised that reduced DMI may be related to zeolite’s suppressive effect on circulating concentrations of P (Thilsing et al. Citation2006), palatability issues (Thilsing-Hansen et al. Citation2002; Thilsing et al. Citation2007; Grabherr et al. Citation2009), or a reduction in digestion rate (Kerwin et al. Citation2019). Further work is required to determine if these effects are associated with the decrease in eating time in grazing dairy cows supplemented with zeolite.
Lying time and activity behaviours
We hypothesized that zeolite-supplemented grazing cows would have shorter lying times and be more active during the peripartum period because of improved blood Ca status and its association with muscle function (Goff Citation2008). Consistent with the literature in housed cows (Thilsing-Hansen and Jørgensen Citation2001; Grabherr et al. Citation2009; Kerwin et al. Citation2019), zeolite improved Ca concentrations around the time of calving and markedly reduced the risk of hypocalcemia in the grazing animals used for the current study (Roche et al. Citation2018; Crookenden et al. Citation2020); however, we observed few effects of treatment on lying and activity behaviours ().
There was a treatment × day interaction in the PERI period (P = 0.05; ) whereby the zeolite-fed cows engaged in more lying bouts on the day after calving (+1.8 per day; P = 0.03; Supplemental Figure 2), which could be a result of improved skeletal muscle contractility due to improved Ca balance immediately postpartum. However, the number of lying bouts did not differ enough to significantly alter the total daily lying time, nor was step count affected by treatment (). Within-day lying time and activity were also unaffected by treatment during the PERI period (all P > 0.13; Supplemental Table 2). Differences in lying time or activity in the immediate periparturient period may only be distinguished between groups with more divergent blood Ca concentrations. For example, longer lying times and reduced activity on the day before and the day of calving were previously reported in cases of clinical hypocalcemia (Ca ≤ 1.4 mmol/L) without paresis in dairy cows grazing pasture; however, the lying time and activity of subclinically hypocalcemic (Ca < 2.0 mmol/L) and normocalcemic (Ca ≥ 2.0 mmol/L) cows did not differ (Hendriks et al. Citation2020). In the current study, although there was a high prevalence of subclinical hypocalcemia (Ca ≤ 2.15 mmol/L on day 0 or 1 postcalving) in the Control 2−3 (70%) and 4+ (80%) groups, there were only two cases of clinical hypocalcemia without paresis (Ca < 1.4 mmol/L on day 0 or 1 postcalving) in the Control 4 + group, and none of the cows experienced clinical hypocalcemia with paresis at calving (i.e. milk fever). Studies in a larger population with a higher prevalence of clinical hypocalcemia with and without paresis in the controls may be required to examine the effects of zeolite on lying and activity behaviours.
Interestingly, supplementing cows with zeolite prepartum was associated with altered lying behaviours after calving. The zeolite-fed cows consistently spent about 30 min longer lying each day in the POST period, which was driven by longer lying bout durations of, on average, 7.8 min per bout (; Supplemental and ). This is an interesting response considering a previous study in grazing dairy cows indicated that the effects of blood Ca status at calving on lying behaviour were short-lived, with no differences observed beyond 3 d postcalving (Hendriks et al. Citation2020). Other studies in housed cows have reported longer lying times (between 0.6 and 2.1 h/d) in clinically and subclinically hypocalcemic cows during the first 21 days in milk (Barraclough et al. Citation2020; Tsai et al. Citation2021). In the current study, the lack of differences in eating time and activity between treatment groups postcalving indicates that the Control cows were likely spending more time standing idle, grooming, or ruminating while standing; however, it is difficult to disentangle the reasons for these differences between the treatment groups. Lying down is a biologically important behaviour in cows (Munksgaard and Simonsen Citation1996), and may become more important as the time available to lie down is limited due to postcalving management. The lying times of the Control cows in the current study (7.5 h/d) were at the lower end of the range of what is reported as ‘typical’ postcalving lying times in grazing cows (range: 7.50–8.58 h/d; Sepúlveda-Varas et al. Citation2014; Hendriks et al. Citation2019b). As adequate lying time is an important indicator of cow comfort and welfare, it is possible that the longer lying times in the zeolite-supplemented cows may indicate they have improved recovery and rest, rather than being indicative of disease. This hypothesis requires further investigation.
Figure 2. The effect of feeding zeolite precalving or parity on hourly profiles of eating and lying times within day during the PRE and POST periods. The temporal pattern of eating time (min/h) and lying time (min/h) within day for the two treatment groups [(a) and (b); Control vs Zeolite] or the two parity groups [(c) and (d); Parity 2–3 vs 4+] during the PRE [(top); −21 to −3 d] and POST [(bottom); 3 to 28 d] periods relative to the day of calving (d 0). Data are presented as raw means on an hourly basis. The average time spent off pasture is represented by the horizontal black lines. Time off pasture for sampling, measurements, and feeding PRE was approximately between 0700 and 1000 h, and for morning sampling/measurements and the morning and afternoon milkings POST was approximately between 0645–0830 h and 1445–1600 h, respectively. Average sunrise and sunset times across the study were 0730 h and 1719h, respectively (Timeanddate.com: Hamilton, New Zealand – Sunrise, Sunset, and Daylength Citation2021).
![Figure 2. The effect of feeding zeolite precalving or parity on hourly profiles of eating and lying times within day during the PRE and POST periods. The temporal pattern of eating time (min/h) and lying time (min/h) within day for the two treatment groups [(a) and (b); Control vs Zeolite] or the two parity groups [(c) and (d); Parity 2–3 vs 4+] during the PRE [(top); −21 to −3 d] and POST [(bottom); 3 to 28 d] periods relative to the day of calving (d 0). Data are presented as raw means on an hourly basis. The average time spent off pasture is represented by the horizontal black lines. Time off pasture for sampling, measurements, and feeding PRE was approximately between 0700 and 1000 h, and for morning sampling/measurements and the morning and afternoon milkings POST was approximately between 0645–0830 h and 1445–1600 h, respectively. Average sunrise and sunset times across the study were 0730 h and 1719h, respectively (Timeanddate.com: Hamilton, New Zealand – Sunrise, Sunset, and Daylength Citation2021).](/cms/asset/2a2a776d-f233-4b32-93dc-19f10313a3e2/tnza_a_2283052_f0002_oc.jpg)
Although we observed few effects of feeding zeolite on lying time or step count in this study, these behaviours were affected by parity and had a reciprocal relationship with eating time. Precalving, the younger (Parity 2-3) cows were more active than the older (Parity 4+) cows, taking 338 more steps/d and lying down for ∼30 fewer min/d (; Supplemental Figure 1). We hypothesise that these differences were driven by the longer time spent eating in the younger animals (; ). A treatment x parity x day interaction was detected PRE and, although this effect varied between days, on average, the Parity 2–3 Controls spent 55.8 min/d longer eating compared with Parity 4+ Controls, which was then restricted slightly to 42.6 min/d when animals were fed zeolite. These precalving parity differences in eating and lying times were driven by hourly differences during the night, particularly between 2200 and 0159 h (both P < 0.01; Supplemental Table 2; ). The parity-driven difference in behaviours continued into the PERI period, whereby there was a more pronounced difference in lying time and activity, and a smaller, but significant, difference in eating time (; Supplemental Figure 1). However, no within-day differences were observed PERI (Supplemental Table 2). While there were no effects of parity on lying behaviours POST, the Parity 2–3 cows tended to be more active than Parity 4 + cows, averaging 200 more steps/d (P = 0.13) and spending 24 min longer eating/d (P = 0.11; ), with differences in eating time occurring between 1800 and 0159 h (both P < 0.05; Supplemental Figure 1). The parity effects on eating time may reflect that the older cows had better grazing efficiency and were more experienced (Dunn et al. Citation1988; Gregorini et al. Citation2013), or possibly that the younger cows were avoiding competition with older cows by grazing at different times.
Shorter lying times PRE and PERI in younger cows could mostly be explained by their greater eating times in the current study but may also reflect the influence of comfort and welfare on the time budgets and motivation to lie in dairy cows (Munksgaard and Simonsen Citation1996; Hendriks et al. Citation2019b). This could particularly be the case during the PERI period, due to increased restlessness in younger cows during calving and increased difficulty coping with the physiological changes during this time (Wehrend et al. Citation2006). Other factors influencing lying behaviour during the PRE and PERI periods may include interactions with other behaviours (e.g. rumination time and social interactions) as well as individual traits (e.g. genetics, personality, other underlying conditions). Further research is required to investigate the factors that influence parity-driven behaviour differences in multiparous grazing dairy cows as parity may have interactive effects on transition cow health management strategies as observed in the current study.
Conclusions
Prepartum supplementation of dairy cows with synthetic zeolite A reduced the daily eating time of younger multiparous grazing dairy cows, indicating that it may have a subtle anorexic effect or affect eating behaviours during the time of supplementation. Although we hypothesised that lying times would be shorter and activity greater immediately postpartum, we did not detect any differences in the immediate PERI period, possibly reflecting a lack of animals in this study experiencing clinical hypocalcemia with or without paresis. Zeolite also increased the daily lying time postpartum across all multiparous age groups, which is potentially associated with improved cow comfort and welfare during early lactation. Furthermore, this study provides evidence that the effects of parity should be considered when investigating the eating, lying, and activity behaviour of grazing dairy cows.
Supplemental material
Download MS Word (770.6 KB)Acknowledgements
The authors gratefully acknowledge the DairyNZ technical and farm teams for their valuable assistance.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Barraclough RAC, Shaw DJ, Thorup VM, Haskell MJ, Lee W, Macrae AI. 2020. The behavior of dairy cattle in the transition period: effects of blood calcium status. Journal of Dairy Science. 103(11):10604–10613. doi: 10.3168/jds.2020-18238.
- Crookenden MA, Phyn CVC, Turner SA, Loor JJ, Smith AI, Lopreiato V, Burke CR, Heiser A, Roche JR. 2020. Feeding synthetic zeolite to transition dairy cows alters neutrophil gene expression. Journal of Dairy Science. 103(1):723–736. doi: 10.3168/jds.2019-17097.
- Dela Rue B, Lee JM, Eastwood CR, Macdonald KA, Gregorini P. 2020. Short communication: evaluation of an eating time sensor for use in pasture-based dairy systems. Journal of Dairy Science. 103(10):9488–9492. doi: 10.3168/jds.2020-18173.
- Dunn RW, Havstad KM, Ayers EL. 1988. Grazing behavior responses of rangeland beef cows to winter ambient temperatures and age. Applied Animal Behaviour Science. 21(3):201–207. doi: 10.1016/0168-1591(88)90109-8.
- Goff JP. 2008. The monitoring, prevention, and treatment of milk fever and subclinical hypocalcemia in dairy cows. The Veterinary Journal. 176(1):50–57. doi: 10.1016/j.tvjl.2007.12.020.
- Grabherr H, Spolders M, Fürll M, Flachowsky G. 2009. Effect of several doses of zeolite A on feed intake, energy metabolism and on mineral metabolism in dairy cows around calving. Journal of Animal Physiology and Animal Nutrition. 93(2):221–236. doi: 10.1111/j.1439-0396.2008.00808.x.
- Gregorini P, Dela Rue B, Pourau M, Glassey C, Jago J. 2013. A note on rumination behavior of dairy cows under intensive grazing systems. Livestock Science. 158(1-3):151–156. doi: 10.1016/j.livsci.2013.10.012.
- Hendriks SJ, Huzzey JM, Kuhn-Sherlock B, Turner SA, Mueller KR, Phyn CVC, Donaghy DJ, Roche JR. 2020. Associations between lying behavior and activity and hypocalcemia in grazing dairy cows during the transition period. Journal of Dairy Science. 103(11):10530–10546. doi: 10.3168/jds.2019-18111.
- Hendriks SJ, Phyn CVC, Turner SA, Mueller KM, Kuhn-Sherlock B, Donaghy DJ, Huzzey JM, Roche JR. 2019b. Lying behavior and activity during the transition period of clinically healthy grazing dairy cows. Journal of Dairy Science. 102(8):7371–7384. doi: 10.3168/jds.2018-16045.
- Hendriks SJ, Phyn CVC, Turner SA, Mueller KR, Kuhn-Sherlock B, Donaghy DJ, Huzzey JM, Roche JR. 2019a. Effect of weather on activity and lying behaviour in clinically healthy grazing dairy cows during the transition period. Animal Production Science. 60(1):148–153. doi: 10.1071/AN18569.
- Kerwin AL, Ryan CM, Leno BM, Jakobsen M, Theilgaard P, Barbano DM, Overton TR. 2019. Effects of feeding synthetic zeolite A during the prepartum period on serum mineral concentration, oxidant status, and performance of multiparous Holstein cows. Journal of Dairy Science. 102(6):5191–5207. doi: 10.3168/jds.2019-16272.
- McArt JAA, Neves RC. 2020. Association of transient, persistent, or delayed subclinical hypocalcemia with early lactation disease, removal, and milk yield in Holstein cows. Journal of Dairy Science. 103(1):690–701. doi: 10.3168/jds.2019-17191.
- McDougall S. 2001. Effects of periparturient diseases and conditions on the reproductive performance of New Zealand dairy cows. New Zealand Veterinary Journal. 49(2):60–67. doi: 10.1080/00480169.2001.36204.
- Munksgaard L, Simonsen HB. 1996. Behavioral and pituitary adrenal-axis responses of dairy cows to social isolation and deprivation of lying down. Journal of Animal Science. 74(4):769–778. doi: 10.2527/1996.744769x.
- National Climate Database. 2018. [NIWA] National Institute of Water and Atmospheric Research. [accessed 2018 August 16]. https://cliflo.niwa.co.nz/.
- Neave HW, Lomb J, von Keyserlingk MAG, Behnam-Shabahang A, Weary DM. 2017. Parity differences in the behavior of transition dairy cows. Journal of Dairy Science. 100(1):548–561. doi: 10.3168/jds.2016-10987.
- Roberts KI, McDougall S. 2019. Risk factors for subclinical hypocalcaemia, and associations between subclinical hypocalcaemia and reproductive performance, in pasture-based dairy herds in New Zealand. New Zealand Veterinary Journal. 67(1):12–19. doi: 10.1080/00480169.2018.1527732.
- Roche JR. 2003. The incidence and control of hypocalcaemia in pasture-based systems. Acta Veterinaria Scandinavica Supplementum. 97:141–144.
- Roche JR, Berry DP. 2006. Periparturient climatic, animal, and management factors influencing the incidence of milk fever in grazing systems. Journal of Dairy Science. 89(7):2775–2783. doi: 10.3168/jds.S0022-0302(06)72354-2.
- Roche JR, Heiser A, Crookenden MA, Burke CR, Turner SA, Kuhn-Sherlock B, Phyn CVC. 2018. The effect of feeding synthetic zeolite A prepartum on indices of mineral and metabolic status, milk production and reproduction in grazing dairy cows. Journal of Dairy Science. 101(Suppl. 2):175. [Abstract]
- Schirmann K, Chapinal N, Weary DM, Vickers L, von Keyserlingk MAG. 2013. Short communication: rumination and feeding behavior before and after calving in dairy cows. Journal of Dairy Science. 96(11):7088–7092. doi: 10.3168/jds.2013-7023.
- Schütz KE, Clark KV, Cox NR, Matthews LR, Tucker CB. 2010. Responses to short-term exposure to simulated rain and wind by dairy cattle: time budgets, shelter use, body temperature and feed intake. Animal Welfare. 19(4):375–383. doi: 10.1017/S0962728600001858.
- Sepúlveda-Varas P, Weary DM, von Keyserlingk MAG. 2014. Lying behavior and postpartum health status in grazing dairy cows. Journal of Dairy Science. 97(10):6334–6343. doi: 10.3168/jds.2014-8357.
- Thilsing T, Jørgensen RJ, Poulsen HD. 2006. In vitro binding capacity of zeolite A to calcium, phosphorus and magnesium in rumen fluid as influenced by changes in pH. Journal of Veterinary Medicine Series A. 53(2):57–64. doi: 10.1111/j.1439-0442.2006.00798.x.
- Thilsing T, Larsen T, Jørgensen RJ, Houe H. 2007. The effect of dietary calcium and phosphorus supplementation in zeolite A treated dry cows on periparturient calcium and phosphorus homeostasis. Journal of Veterinary Medicine Series A. 54(2):82–91. doi: 10.1111/j.1439-0442.2007.00887.x.
- Thilsing-Hansen T, Jørgensen RJ. 2001. Hot topic: prevention of parturient paresis and subclinical hypocalcemia in dairy cows by zeolite A administration in the dry period. Journal of Dairy Science. 84(3):691–693. doi: 10.3168/jds.S0022-0302(01)74523-7.
- Thilsing-Hansen T, Jørgensen RJ, Enemark JMD, Larsen T. 2002. The effect of zeolite A supplementation in the dry period on periparturient calcium, phosphorus, and magnesium homeostasis. Journal of Dairy Science. 85(7):1855–1862. doi: 10.3168/jds.S0022-0302(02)74259-8.
- Timeanddate.com: Hamilton, New Zealand – Sunrise, Sunset, and Daylength. 2021. [accessed 2021 March 13]. https://www.timeanddate.com/sun/new-zealand/hamilton.
- Tsai IC, Mayo LM, Jones BW, Stone AE, Janse SA, Bewley JM. 2021. Precision dairy monitoring technologies use in disease detection: differences in behavioral and physiological variables measured with precision dairy monitoring technologies between cows with or without metritis, hyperketonemia, and hypocalcemia. Livestock Science. 244(3):104334. doi: 10.1016/j.livsci.2020.104334.
- Wehrend A, Hofmann E, Failing K, Bostedt H. 2006. Behaviour during the first stage of labour in cattle: influence of parity and dystocia. Applied Animal Behaviour Science. 100(3-4):164–170. doi: 10.1016/j.applanim.2005.11.008.
- Wilkens MR, Nelson CD, Hernandez LL, McArt JAA. 2020. Symposium review: Transition cow calcium homeostasis—Health effects of hypocalcemia and strategies for prevention. Journal of Dairy Science. 103(3):2909–2927. http://doi.org/10.3168/jds.2019-17268.