Abstract
Knowledge of available variability in a crop species provides a guide for the utilization and conservation of useful variation. Seventeen agro-morphological traits were studied in 184 accessions of cacao, Theobroma cacao L. collected from farmers' fields (138) and field genebank collections (46). Descriptive statistics—univariate analysis of variance (ANOVA)—were used to determine variation among accessions. Multivariate analysis with principal component analysis (PCA) was done and a cluster dendrogram was generated based on the unweighted pair group mean with arithmetic average (UPGMA). The first three principal component axes accounted for 43.6% of total variation observed among accessions studied. Quantitative bean traits—dry bean weight, nib weight, fresh weight, cotyledon length and cotyledon width—and qualitative fruit traits—basal constriction, apex form, ridge colour, fruit shape, flush colour and cotyledon shape—were the most important traits accounting for the variability observed. Six groups of accessions were obtained from cluster analysis. Fruit and bean traits of Upper Amazon Forasteros observed in farmers' accessions provided evidence of a shift from previously grown local ‘West African Amelonado’ from the Lower Amazon Forastero population. In conclusion, this study revealed some accessions with useful trait combinations in farmers' plantations and indicated a need to conserve the local ‘Amelonado’ landrace variety to avoid extinction and to preserve the unique flavour of ‘West African Amelonado’ in cocoa breeding programmes.
Introduction
It is estimated that the world's cocoa exports, which amount to US$5–6 billion/year and provide cocoa butter, powder and liquor used in the manufacture of chocolate, cosmetics and other products, drive a market worth approximately US$70 billion and provide more than 60,000 jobs in the USA alone (Guiltinan Citation2007). Cocoa is a major foreign exchange-earning commodity crop for West and Central African countries such as Ivory Coast, Ghana, Nigeria and Cameroon which together produce more than 70% of the world's cocoa export of 3.592 million metric tons (ICCO Citation2008). In West Africa, more than 96% of cocoa production is carried out by smallholder farmers who rely on proceeds from the sale of cocoa beans as a major source of family income. Proceeds from export also provide substantial revenue for the governments’ capital development projects (Aikpokpodion Citation2007). The presence of significant genetic diversity in genetic resources maintained in farmers' fields and field genebanks could provide insurance against any present or future threat to cultivation and contribute to the sustainability of the world's cocoa economy.
Cacao is an understorey tree crop species native to the Amazon Basin belonging to the Sterculiaceae family (Purseglove Citation1974). However, recent molecular studies have revealed that Theobroma cacao belongs to the Malvaceae family (Alverson et al. Citation1999) and was introduced into Africa by Spanish and Portuguese seafarers—first to São Tome in 1822 and then to Fernando Po (now Bioko in Equatorial Guinea) in 1855 (Wood and Lass Citation1985; Bartley Citation2005). In São Tome, two cacao trees, which were successfully established from a batch of plants brought in from Bahia, Brazil in 1822, became the parents of the subsequent cacao trees of the island (Toxopeus Citation1973). Swiss missionaries also made other introductions, from Suriname, and the first cocoa seeds were sown on the African mainland in 1857 (Lanaud et al. Citation2000).
Cacao was first introduced by Chief Squiss Ibaningo into Bonny in present-day Rivers State of Nigeria in 1874 from Fernando Po (Opeke Citation1969). Apart from Chief Squiss Ibaningo, missionaries, slave traders and the former Nigeria Department of Agriculture also introduced several cacao germplasm materials. As a result of these introduction efforts, a fair range of variability was then present but all within the ‘Amelonado’ population (Aikpokpodion Citation2007). However, the cultivation and development of the cocoa industry in Nigeria only began in the 1890s with 95% of the production export of 21 tonnes in 1895 coming from the Western Region (Opeke Citation1969). The first materials planted were of the Lower Amazon Forastero population (‘Amelonado’), but this was followed by the introduction of the hybrid ‘Trinitario’ and ‘Criollo’ types in 1920 which formed hybrids with the original ‘Amelonado’ type (Toxopeus Citation1972). In 1944, several accessions from Upper Amazon Forastero and ‘Trinitario’ populations were introduced from Trinidad to widen the genetic base of the germplasm which was lacking in variability at the time (Posnette & Todd Citation1951). Between 1965 and 1967, a large-scale introduction of Upper Amazon cacao materials was made from Trinidad as part of the Trinidad–Nigeria Cacao Introduction Scheme sponsored by the Cocoa Alliance (Atanda Citation1977). This was also documented in the Annual Reports of Cacao Research, ITCA, 1966 and 1967 . In this scheme, some 701 progenies and 313 clones of intra-Nanay, intra-Parinari, intra-Iquitos and inter-P (Pound's selections) crosses from a total of 350 crosses were introduced (Olatoye and Esan Citation1992). Some materials were also acquired from Costa Rica, Indonesia, Fernando Po, Kew Gardens (UK), Wageningen (The Netherlands) and Miami (USA) (Jacobs et al. Citation1971). These materials formed the basis of cultivars developed in the breeding programmes and planting materials distributed to farmers.
Although molecular markers are more commonly used in the assessment of genetic diversity of cacao (N'Goran et al. Citation1994; Sounigo et al. Citation2005; Cryer et al. Citation2006; Aikpokpodion et al. Citation2009), highly heritable morphological and agronomic characters are still useful for germplasm characterization (Klug & Cummings Citation1994; Sounigo et al. Citation1997). In a recent study, Bekele et al. (Citation2006) showed that classification of accessions in the International Cocoa Genebank, Trinidad (ICG,T) based on morphological variation was congruent with the traditional classification of the cacao types.
Morpho-agronomic characteristics of the pods, seeds and flowers have also been used to evaluate relationships among cacao genotypes (Engels Citation1986; Bekele & Bekele Citation1996; Lachenaud et al. Citation1999; Lachenaud & Oliver Citation2005). Previous diversity studies have often concentrated on field genebank collections (Bekele et al. Citation2006; Engels Citation1986; Lachenaud & Oliver Citation2005), however, this study focused on cacao accessions and diversity maintained on-farm in field plantations by farmers.
As an introduced tree crop species, cacao has been grown as an important cash crop by the West African farmers for more than a century and cultivation has witnessed several challenges including the devastating effect of the cocoa swollen shoot virus disease particularly in the 1930s and 1940s (Posnette Citation1947). Since the mid-1950s, new varieties of cacao, largely of Upper Amazon Forastero population, had been released and distributed for planting both as a replacement for the genetically uniform local ‘Amelonado’ types that had been cut out as a remedy for the virus disease attack and as new plantings for production expansion. In Nigeria, Toxopeus (Citation1964) reported that up to 1961, more than 20 million seedlings of the newly developed synthetic variety had been distributed. Since then, there has been little or no information on the extent of diversity in field-cultivated cacao. This study was therefore necessary to determine the level of variation in field-grown cacao in order to provide information that could be useful for future breeding strategies, on-farm selection and conservation efforts.
The main objective of this study therefore, was to determine variation in agro-morphological characteristics of cacao grown in farmers' fields and determine their relationships with some genebank accessions used in the development of varieties distributed for planting.
Materials and methods
Plant materials
A total of 184 samples used for this study was comprised of 138 accessions collected from farmers' plantations in cocoa growing areas of Nigeria () and 46 accessions from seed gardens and multiplication units maintained in the field genebank of the Cocoa Research Institute of Nigeria, Owena in Ondo State (07.20° N, 5.03° E; 267 m above sea level) that are used to develop hybrid varieties for distribution to farmers.
Table 1 Accession codes, local government area (LGA) and geographical locations of cocoa plantations used in this study
Twenty-three cocoa plantations were sampled and the number of trees sampled ranged from 4 to 12 per farm, depending on the tree population and criteria defined with farmers. Samples were randomly selected from trees considered by farmers as their ‘best trees’ in terms of productivity; ‘choice’ trees from which pods were collected to raise new seedlings for planting (‘mother tree’); and ‘worst trees’ that would not be used for planting materials.
From the field genebank, accessions with small bean sizes (less than 1.0 g) like C77 (T85/799) and large bean sizes such as T17/11 derived from ‘Iquitos Mixed Calabacillo’ (IMC) population were included with other hybrid genotypes distributed to farmers such as CRIN Selected Accessions (CSA), CRIN Elite Selected Accessions (CESA), CRIN Series 2 hybrids (CS2H), Trinidad Introduction (CTIS and T-Clones).
Data collection
Data were collected on cacao accessions following the Short Descriptor List of the Cocoa Research Unit, Trinidad and Tobago (Bekele & Butler Citation2000) adapted from IBPGR Cacao descriptor list (IBPGR Secretariat Citation1981). The morphological descriptors used were carefully selected from the International Board for Plant Genetic Resources descriptor list for cacao (IBPGR Secretariat Citation1981) and have been found to be most discriminative, taxonomically useful and precluded redundancy (Engels Citation1986; Lachenaud et al. Citation1999; Bekele & Butler Citation2000; Bekele et al. Citation2006). Each farmer's selection was characterized in terms of the 17 traits listed in . Only healthy, mature pods were selected for characterization. Fruit shape, surface texture (rugosity), anthocyanin pigmentation (colour) on the ridge of mature pods, ridge disposition, basal constriction and apex form were scored, while fruit length and width were measured. Fruits were considered mature when their colour had turned (or were about to turn) yellow (in the case of green pods) or orange (in the case of red pods). Wet bean mass was separated from the central placenta and number of beans per pod was counted. For each accession, three fresh beans were collected from the middle section of each of the 10 pods and bulked. Cotyledon length and width were measured and the fresh weight of 20 randomly selected beans was determined using a Sartorius balance (BP 610 model). Dry weight was determined after samples had been oven-dried at 105°C for 24–30 hours and nib weight was recorded after the seed-coat had been removed from the dried beans.
Table 2 Characters and states of 17 agro-morphological traits studied in cacao accessions from farmers' plantation and the field genebank of the Cocoa Research Institute of Nigeria (CRIN)
Data analysis
Descriptive statistics were derived for all traits observed and all eight quantitative variables used for analysis showed no departure from normal distribution. Principal component analysis (PCA) with all 17 traits was carried out to determine the relative importance of the traits responsible for variation among cacao accessions. The aforementioned statistical analyses were conducted using SAS v.9.1 (CitationSAS Institute Inc, NC, USA). Cluster analysis with all 17 traits was used to explore relationships among the accessions based on the unweighted pair group method with the arithmetic mean (UPGMA) implemented with the NTSYSpc v. 2.02 package (Applied Biostatistics, Inc). The general linear model (GLM) procedure was specified for the analysis of variance (ANOVA) and mean separation was performed on the a priori clusters to determine cluster characteristics.
Results
The descriptive statistics of the quantitative traits of cacao accessions from field genebank and farmers' plantations are shown in while the qualitative traits are shown in . Test of goodness of fit showed no significant deviation from normal distribution in all the quantitative traits. Correlation analysis () showed significant positive correlation among all traits except between number of beans per pod on the one hand, and cotyledon length, cotyledon width, dry weight and nib weight, on the other.
Table 3 Mean, standard error (in parenthesis), coefficient of variation (%) and levels of significance from analysis of variance (ANOVA) of 184 cacao accessions from field genebank and farmers plantation in Nigeria
Table 4 Frequency distribution of nine qualitative traits in cacao accessions from CRIN's field genebank and farmers' plantation
Table 5 Correlation matrix of eight quantitative traits of cacao accessions
Principal component analysis of all 17 morphological traits revealed that the first three principal component (PC) axes accounted for 43.6% of total variation among cacao accessions (). Examination of the first PC axis which accounted for 24.5% of total variation showed that it was composed mainly of bean traits—dry bean weight, fresh bean weight, cotyledon length and cotyledon width. The second PC axis accounting for 10.8% of total variation was mainly composed of fruit traits—fruit length, fruit shape, rugosity and apex form. Although the fruit length and rugosity were positively correlated, fruit shape and apex form were negatively correlated to this axis. The third PC axis which made up 8.3% of total variation was composed mainly of qualitative traits of flush colour, ridge colour and cotyledon shape. The fourth PC axis which accounted for 8.0% of variation observed was composed mainly of fruit rugosity, basal constriction, apex form and cotyledon colour. Qualitative characteristics including fruit shape, basal constriction, apex form, cotyledon shape, rugosity (wartiness), ridge pair disposition and cotyledon colour were important traits which accounted for 23.29% of total variation in PC5, PC6, PC7 and PC8 axes. Cotyledon length, fruit width and number of beans per pod were also important quantitative traits accounting for the variance found in these PC axes.
Table 6 Eigenvectors of 17 agro-morphological traits of cacao accessions in first three principal component (PC) axes explaining 43.6% of total variation
Scatter plots of the first two principal component axes () showed the importance of some characteristics in showing relationships among the cacao accessions. Cluster analysis showing relationships among all accessions () showed three distinct groups corresponding to six clusters due to sub-clustering. Accessions with the largest nib, dry and fresh weights, fruit length and fruit width, cotyledon length and cotyledon width were grouped into cluster 3 (). Although fresh bean weight was greater in cluster 3A, cotyledon length was greater in cluster 3B. Accessions with the least bean weight, cotyledon length, cotyledon width and smallest fruit size were grouped into cluster 2. Fruits of accessions in cluster 3 were largely elongate in shape with a broad base (‘Angoleta’) and usually have slight to intense anthocyanin pigmentation in the ridge of their mature fruits. On the other hand, accessions in clusters 1 and 2 appeared to be mostly ‘Cundeamour’ and ‘Amelonado’ as evident from the fruit length and fruit width. Fruits in these clusters were always green and rarely had anthocyanin pigmentation present on the ridges of their mature fruits. Nib weight, fresh and dry bean weights of accessions in cluster 3 were almost twice those of accessions in cluster 2. There was however, no significant difference in the number of beans per pod, flush colour, basal constriction, apex form, rugosity, shape and colour of cotyledon among the six clusters.
Fig. 1 Scatter plot of cacao accessions on principal component axes 1 and 2 (PC1 and PC3) accounting for 35.3% of total variation. i – cotyledon fresh weight (g), ii – cotyledon dry weight (g), iii – nib weight (g), iv – fruit length (cm), v – cotyledon colour, vi – ridge pair disposition, vii – fruit shape, viii – apex form, ix – cotyledon width (cm), x – cotyledon length (cm).
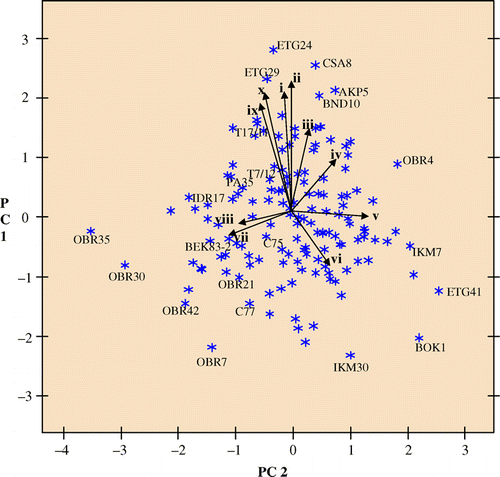
Fig. 2 Dendrogram based on the unweighted pair group arithmetic mean average (UPGMA) showing six clusters of on-farm accessions and field genebank accessions (in boxes) of cacao in Nigeria.
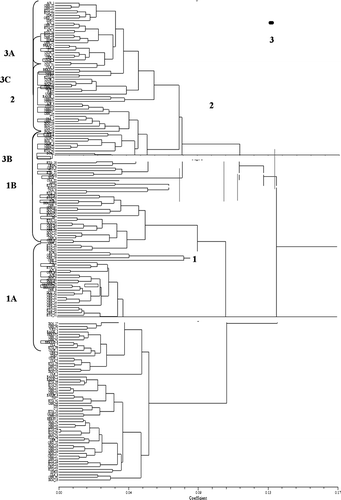
Table 7 Group characteristics of the six clusters based on 17 morphological traits of cacao accessions
The genebank clones included in the analysis were useful as checks to confirm observations made. Clones T17/11 (a progeny of Imperial Mixed Calabacillo ‘IMC’ IMC53 known for large beans and pod size), PA35 (‘Parinari’) and T7/12 (progeny of M 8 [SUR] from the Maraboen River, Suriname) were classified into cluster 3, which represented 40.8% of samples studied. Cluster 3A was mainly composed of red fruited ‘Trinitarios’ also known for large beans, and clusters 3B and 3C most likely represented ‘Upper Amazon’ hybrid cocoa with large beans and fruits. In contrast, clone C77 (T85/799) with small beans was classified into cluster 2. Cluster 2, which made up 14.7% of the accessions studied consisted of accessions with small beans (0.82 g mean dry bean weight) and included many of the accessions known as ‘local Amelonado’ by the farmers. Accessions in cluster 1 had intermediate trait values and made up the rest of the 44.6% of the samples studied.
ANOVA of the a priori groups showed significant differences (P < 0.001) among clusters in all quantitative traits except number of beans per pod (). There were also significant differences (P < 0.001) in fruit shape, ridge colour and ridge pair disposition among the six clusters. However, ANOVA also showed no significant differences between field genebank and farmers' plantation accessions among all quantitative traits except in fresh bean weight (P < 0.001), dry bean weight (P < 0.001) and number of beans per fruit (P < 0.001) ().
Discussion
In this study, the variation in morphological traits among cacao accessions grown in farmers' fields and their relationship with accessions from a field genebank collection was evaluated. This is unlike previous studies where the emphasis was on morphological variation of cacao in field genebank collections (Engels Citation1983; Bekele et al. Citation1994; Bekele & Bekele Citation1996; Iwaro et al. Citation2003; Bekele et al. Citation2006). Analysis of variance showed significant morphological variation in cacao accessions maintained in farmers' field collections and also between accessions from the field genebank and farmers' fields in bean and fruit traits. While nib, fresh and dry bean weights were greater in the field genebank accessions, the number of beans per pod was greater in the farmers' field accessions. It appeared that the higher number of beans per pod was related to the lower fresh bean weight in farmers' accessions as shown from the correlation values between these variables (r = − 0.131, P < 0.0001). A similar relationship (r = − 0.19; P < 0.001) was also reported among cacao accessions from a genebank in Trinidad (Iwaro et al. Citation2003). However, there was no significant difference among accessions in fruit length and cotyledon width.
The observed variation permitted the accessions to be classified into six clusters. From the classification obtained, it appeared that accessions with large bean and fruit size in cluster 3 displayed the characteristics of Upper Amazon derived varieties and ‘Trinitario’ materials distributed to farmers from seed gardens. On the other hand, the small bean and fruit size of accessions in cluster 2 were characteristic of the local ‘Amelonado’ cacao. Intermediate values for bean weight and fruit characteristics obtained for accessions in cluster 1 indicated possible hybridization that could have taken place in the field as a result of the introduction of Upper Amazon materials to farmers' fields which were initially dominated by the old ‘local Amelonado’ variety. In their study of some 600 accessions of cacao maintained in the International Cocoa Germplasm, Trinidad (ICG,T), Bekele et al. (Citation2006) found that phenotypic data showed distinct classification of accessions that were congruent with recognized genetic classes and were able to differentiate among the ‘Trinitarios’, ‘Forasteros’ and ‘Refractarios’. Engels (Citation1983, Citation1986) also showed the usefulness of bean characteristics in showing affinity between ‘Trinitario’ and ‘Criollo’ cacao populations.
Some accessions with large dry bean weight and number of beans per pod: ETG24 (1.6 g, 36.3/pod), ETG29 (1.5 g, 46/pod), AKP5 (1.5 g, 54/pod), IKM11 (1.4 g, 40.7/pod), ETG 40 (1.4 g, 42.8/pod), IKM7 (1.4 g, 43.9/pod) and OBR44 (1.4 g, 43.7/pod) identified in cluster 3 showed promise for selection as clones with a low pod index. On the other hand, accessions in cluster 2 with low dry bean weight such as OBR3 (0.67 g), BOK1 (0.68 g), OBR59 (0.68 g), AJS3 (0.75), ETG41 (0.76 g) and OBR22 (0.85) could be useful for further study as their characteristic low bean weight and size reflected the trait of ‘local Amelonado’, a traditional variety grown by farmers before the release of introduced Upper Amazon derived materials around 1956. Selection of some of these individual genotypes as individual clones could be justified; Bartley (Citation2005) also noted that cultivated populations that are products of recombination usually have greater genetic diversity, and consequently, a larger number of individual genotypes than the source populations.
The large variation observed in this study for cacao grown by farmers indicates a high level of heterogeneity in materials maintained on-farm. The current field composition of cacao indicates a shift from the situation preceding the 1950s, when uniform ‘Amelonado’ cacao types were mainly grown. This would have resulted from the use Upper Amazon-derived cacao varieties distributed to farmers through the seed gardens. As observed by Bartley (Citation2005), the introduction of materials from another source into an original population could result in hybridization between them, and subsequent recombination among these hybrids could lead to genotypes genetically distinct from the parental varieties but possibly having phenotypic ranges encompassing those of the parents.
Quantitative bean traits—dry bean weight, nib weight, fresh weight, cotyledon length and cotyledon width—and qualitative fruit traits—basal constriction, apex form, ridge colour and fruit shape—were among the most important traits accounting for the variation observed among the samples studied. The quantitative bean traits were however most useful for classification. These nine characteristics were also among the traits found by Bekele et al. (Citation2006) to explain more than 50% of phenotypic variation in the germplasm collection studied. However, the dispersion of variation among the first nine principal component axes, which explained 76.7% of total variation, indicated a need to use the full complement of the recommended morphological traits rather than these nine characters alone. Significant variation observed for bean and fruit characteristics among cacao accessions in this study indicated the importance of on-farm collections as a valuable reservoir of genetic diversity. Some of these traits are of commercial importance and have been used as selection criteria by farmers in the choice of parent trees for raising seedlings to make new plantings and used for farm expansion.
From this study, the complete absence or slight anthocyanin pigmentation on the ridge of the mature fruit of more than 88% of accessions indicated that most of the cacao now grown in Nigeria was apparently derived from the Amazonian Forastero origin. This observation may further confirm the report that Amazonian Forastero provides more than 95% of the world's cocoa output (Bekele et al. Citation2006). The preponderance of ‘Cundeamour’ fruit shape (>76%) with slight to strong basal constriction (>88%), obtuse to attenuate apex forms (95%) and intermediate to intense rugosity (86%) showed that, possibly, the Upper Amazon Forastero (UAF) ‘Parinari’ population, characterized by pronounced bottleneck, conspicuous apex form and the intermediate to intensely warty fruit (Bartley Citation2005; Bekele et al. Citation2005) and to a lesser extent, the ‘Nanay’ population, had the most impact on cacao grown in Nigeria. This could be explained by the fact that clones from these populations have been largely used in Nigerian cacao breeding programmes and form the basis of the released F3 Amazon variety and others (Toxopeus Citation1964; Olatoye & Esan Citation1992). It has also been reported that UAF accounted for about 60% of cacao cultivation worldwide (Wood & Lass Citation1985; Eskes & Lanaud Citation2001).
On the other hand, the low percentage (less than 15%) of red pigmentation in fruits, a trait known to be under the control of a single dominant gene (Bartley Citation2005) associated with some ‘Criollo’ populations indicated that ‘Criollo’ and red-podded ‘Trinitario’ populations have, at present, only a minimal influence on field-grown cacao in Nigeria. This could be related to the limited use of ‘Trinitario’ clones in the Nigerian cocoa breeding programme in developing improved materials distributed to farmers, a situation similar to that observed by Lockwood (Citation1976) in Ghana.
The low percentage of fruit traits that are typical of ‘Amelonado’ and ‘Trinitario’ types provides some evidence of variety replacement of ‘West Africa Amelonado’ (WAA) cacao types in farmers' fields with Upper Amazon-derived types and hybrids between them. This reported erosion of West African ‘Amelonado’ presents a serious case for the conservation of this population which may be completely lost if the present trend continues in farmers' fields. In a study of factors influencing farmers' choice of mother trees as sources of new planting on farms, Aikpokpodion et al. (Citation2003) showed farmers' preference for Upper Amazon Forastero-derived types as sources of seeds for new planting due to their good vigour allowing for easier field establishment, enhanced precocity (flowering three years after field establishment, compared with five to seven years for WAA), year-round fruit production and higher yields. These factors could have been responsible for this observation.
Although of low vigour, the WAA trees are amenable to high-density plant ing and the beans possess flavour attractive to chocolate manufacturers. Aikpokpodion et al. (Citation2003) also showed that farmers in Nigeria prefer green-podded trees as ‘mother trees’ or sources of planting materials rather than red-podded trees. An exception is in areas where farm sizes are small (e.g., marginal cocoa-growing areas) and the known high-yielding ability of the red-podded ‘Trinitario’ trees becomes the most important factor in spite of the high Phytophthora pod rot incidence associated with ‘Trinitario’.
The WAA or ‘bulk cocoa’ cocoa, known for its strong chocolate flavour, is commonly used for the production of high-volume chocolate lines. The shift in cacao types cultivated on farms observed in this study indicates the need to maintain the flavour quality profile characteristic of WAA in cocoa breeding programmes. In a flavour assessment done on UAF families used in developing the ‘F3 Amazon’ variety, Wadsworth (Citation1953) had shown that it was within the WAA flavour range, although it was harsher than a really good WAA cocoa. It is important to ensure that the WAA flavour profile used as standard for bulk cocoa is not compromised with particular reference to the Nigerian cocoa, often regarded as closest to the Ghanaian cocoa used as reference for ‘bulk cocoa’ on the world market (Anon Citation1991).
The shape of the cotyledon, largely ovate and elliptical, indicates a high level of uniformity for the shape of cocoa beans produced in Nigeria. The beans were purple coloured, varying from light to dark with more than half, medium purple. The mean dry weight of cocoa beans from Nigeria was slightly higher than the minimum weight of 1.0 g required in the international market (Wood & Lass Citation1985). In order to exploit variation in these cacao collections, more studies are needed to evaluate the quality characteristics such as butter fat content and flavour of accessions within each group. This should have emphasis on the preservation of the flavour profile of WAA bulk cocoa.
Acknowledgements
The field assistance of Messrs. Lateef Raji, Marcus Efunla, Segun Adeyanju, Sunday Njoku, Victor Enagu and Sunday Taiwo; financial support from the USAID/USDA-ARS, Mars Inc and three anonymous referees are gratefully acknowledged.
References
- Aikpokpodion PO 2007 . Genetic diversity in Nigerian cacao, Theobroma cacao L. collections as revealed by phenotypic and simple sequence repeats marker . PhD thesis submitted to the University of Ibadan, Nigeria .
- Aikpokpodion PO , Badaru K , Kolesnikova-Allen M , Ingelbrecht I , Adetimirin VO 2003 . Farmer-researcher participatory on-farm selection of improved cocoa varieties: the Nigerian experience INGENIC International Workshop on Cocoa Breeding for Improved Production Systems , 19–21 October 2003, Accra, Ghana 183 188 .
- Aikpokpodion , PO , Motamayor , JC , Adetimirin , VO , Adu-Ampomah , Y , Ingelbrecht , I , Eskes , AB , Schnell , RJ and Kolesnikova-Allen , M . 2009 . Genetic diversity assessment of sub-samples of cacao, Theobroma cacao L. collections in West Africa using simple sequence repeats marker . Tree Genetics & Genomes , 5 : 699 – 711 .
- Anon 1991 . Influence of genetic factors and agroclimatic conditions on the quality of cocoa . 2nd International Congress on Cocoa and Chocolate, May 1991, Munich. Nestle Ltd, Agricultural Service 11 27 .
- Atanda OA 1977 . The Third Nigerian Cocoa Breeding Programme: A progress report . Proceedings of the 5th International Cocoa Research Conference, 1–9 September 1975, Ibadan, Nigeria 85 90 .
- Alverson , WS , Whitlock , BA , Nyfeller , R , Bayer , C and Baum , DA . 1999 . Phylogeny of the core Malvales: Evidence from ndhf squence data . American Journal of Botany , 86 : 1474 – 1486 .
- Bartley BGD 2005 . The genetic diversity of cacao and its utilization . Wallingford, , UK , CABI Publishing .
- Bekele , FL and Bekele , I . 1996 . A sampling of the phenetic diversity of cacao in the international cocoa gene bank of Trinidad . Crop Science , 36 : 57 – 64 .
- Bekele , FL , Bekele , I , Butler , DR and Bidaisee , GG . 2006 . Patterns of morphological variation in a sample of cacao (Theobroma cacao L.) germplasm from the International Cocoa Genebank, Trinidad . Genetic Resources and Crop Evolution , 53 : 933 – 948 .
- Bekele FL , Bidaisee GG , Persad N , Bhola J 2005 . Examining phenotypic relationships among Upper Amazon Forastero clones Annual Report 2004 . St Augustine, Trinidad and Tobago, Cocoa Research Unit, the University of the West Indies 27 42 .
- Bekele FL , Butler DR 2000 Proposed list of cocoa descriptors for characterization Eskes AB Engels JMM Lass RA Working procedures for cocoa germplasm evaluation and selection Proceedings of the CFC/ICCO/IPGRI Project Workshop, 1–6 February 1998, Montpellier, France: IPGRI 41 48 .
- Bekele , FL , Kennedy , AJ , McDavid , C , Lauckner , FB and Bekele , I . 1994 . Numerical taxonomic studies in cacao (Theobroma cacao L.) in Trinidad . Euphytica , 75 ( 3 ) : 231 – 240 .
- Cryer , NC , Fenn , MGE , Turnbull , CJ and Wilkinson , MJ . 2006 . Allelic size standards and reference genotypes to unify international cocoa (Theobroma cacao L.) microsatellite data . Genetic Resources and Crop Evolution , 53 : 1643 – 1652 .
- Engels , JMM . 1983 . A systematic description of cacao clones III. Relationships between clones, between characteristics and some consequences for cocoa breeding . Euphytica , 32 : 7119 – 733 .
- Engels , JMM . 1986 . The identification of cacao cultivars . Acta Horticultura , 182 : 195 – 202 .
- Eskes AB , Lanaud C 2001 . Cocoa Charrier A , Jacquot M , Hamon S , Nicolas D . (Razdan MK, translation ed) Tropical plant breeding . Plymouth, UK, Science Publishers and Montpellier, France, CIRAD 78 105 .
- Guiltinan MJ 2007 . Cacao: Biotechnology in agriculture and forestry, Vol 60 Pua EC , Davey MR Transgenic crops V Berlin Heidelberg, Springer-Verlag .
- IBPGR Secretariat 1981 . Report: International Board for Plant Genetic Resources (IBPGR) working group on genetic resources of cacao . ACP, IBPGR/80/56, March 1981, Rome .
- ICCO (International Cocoa Organization) 2008 . Quarterly Bulletin Cocoa Statistics . Vol. XXXV. 2007/2008 .
- Iwaro , AD , Bekele , FL and Butler , DR . 2003 . Evaluation and utilization of cacao (Theobroma cacao L.) germplasm at the International Cocoa Genebank, Trinidad . Euphytica , 130 : 207 – 221 .
- Jacob VJ , Atanda OA , Opeke LK 1971 . Cacao breeding in Nigeria. In Progress in Tree Crops Research in Nigeria . CRIN Commemorative Book 23 33 .
- Klug , WS and Cummings , MR . 1994 . Concepts of genetics , 4th edn , USA : Macmillan College Publishing Company .
- Lachenaud , Ph , Bonnot , F and Oliver , G . 1999 . Use of floral descriptors to study variability in wild cocoa trees (Theobroma cacao L.) in French Guiana . Genetic Resources and Crop Evolution , 46 : 491 – 500 .
- Lachenaud , Ph and Oliver , G . 2005 . Variability and selection for morphological bean traits in wild cacao trees (Theobroma cacao L.) from French Guiana . Genetic Resources and Crop Evolution , 52 ( 3 ) : 225 – 231 .
- Lanaud C , Motamayor JC , Risterucci AM 2000 . Implications of new insight into the genetic structure of Theobroma cacao L. for breeding strategies Proceedings of the International Workshop on New Technologies and Cocoa Breeding , 16–17 October, Kinabalu, Sabah, Malaysia, INGENIC 89 107 .
- Lockwood , G . 1976 . A comparison of the growth and yield during a 20 year period of Amelonado and Upper Amazon hybrid cocoa in Ghana . Euphytica , 25 : 647 – 658 .
- N'Goran , JAK , Laurent , V , Risterucci , AM and Lanaud , C . 1994 . Comparative genetic diversity of Theobroma cacao L. using RFLP and RAPD markers . Heredity , 73 : 589 – 597 .
- Olatoye ST , Esan EB 1992 . Recent innovation in cacao genetic resources conservation in Nigeria Proceedings of the International Workshop on Conservation, Characterization and Utilization of Cacao Genetic Resources in the 21st Century , 13–17 September 1992, Cocoa Research Unit, Port of Spain, Trinidad 281 291 .
- Opeke LK 1969 . Development of cacao industry in Nigeria . In: Proceedings of the 6th Conference of the Agricultural Society of Nigeria 1 5 .
- Posnette , AF . 1947 . Virus diseases of cocoa in West Africa. 1. Cacao viruses 1A, 1B, 1C and 1D . Annals of Applied Biology , 34 : 388 – 402 .
- Posnette , AF and Todd , JMA . 1951 . Virus diseases of cacao in West Africa. VIII. The search for virus resistant cacao . Annals of Applied Biology , 38 : 785 – 800 .
- Purseglove , JW . 1974 . Tropical crops, dicotyledons , New York : John Wiley & Sons .
- SAS Institute 1999 SAS/STAT User's Guide, ver. V8 . SAS Institute , Cary, NC .
- Sounigo O , Bekele F , Bidaisee G , Christopher Y , Umaharan R 1997 . Comparison between genetic diversity data obtained from morphological, biochemical and molecular studies Cocoa Research Unit, Report for 1997 . St Augustine, Trinidad, The University of the West Indies 20 29 .
- Sounigo , O , Umaharan , R , Christopher , Y , Sankar , A and Ramdahin , S . 2005 . Assessing the genetic diversity in the International Cocoa Genebank, Trinidad (ICG,T) using isozyme electrophoresis and RAPD . Genetic Resources and Crop Evolution , 52 : 1111 – 1120 .
- Toxopeus H 1964 . F3 Amazon cacao in Nigeria . Report of the Cacao Research Institute, Nigeria, 1963/64 13 22 .
- Toxopeus H 1972 . Cocoa breeding: a consequence of mating system, heterosis and population structure . In: Proceedings of the Conference on Cocoa and Coconuts in Malaysia. Kuala Lumpur, Malaysia, Incorporated Society of Planters .
- Toxopeus , H . 1973 . History of the Nigerian cocoa crop in the light of the socio-economic revolution of the 19th century in West Africa . Landbouwkundig Tijdschrift , 83 ( 12 ) : 485 – 490 .
- Wadsworth RV 1953 . Flavour of new types of cacao introduced into West Africa . In: Report of Cocoa Conference, London, 1953 68 75
- Wood , GA and Lass , RA . 1985 . Cacao , 4th edn , London, , UK : Longman .
- Zar , J . 1999 . Biostatistical analysis , 4th edn , New Jersey, , US : Prentice-Hall .