Abstract
Chromosome numbers were determined for plants of five species and seven additional entities of uncertain taxonomic status of Brachyscome. We confirmed the chromosome numbers of several species but found new numbers in plants related to B. humilis, B. montana and B. radicata. Karyotype analysis showed a common karyotype in the diploids and suggested that the polyploids are autopolyploids. There is some evidence of aneuploidy in plants related to B. sinclairii and B. radicata. Karyotype analysis suggests that structural chromosome changes may have accompanied polyploidy in tetraploid B. humilis.
The genus Brachyscome Cass. (=Brachycome orth. var. Cass.) in the Asteraceae is amongst the chromosomally most variable genera of angiosperms (Weiss-Schneeweiss & Schneeweiss Citation2013). In Australia, where the majority of species occur, there are species with 15 different chromosome numbers that range from 2n=4 to 2n=36, large differences in total chromosome length and also in the degree of karyotype asymmetry (Smith-White et al. Citation1970; Watanabe et al. Citation1999). In New Zealand, Webb et al. (Citation1988) recognize five endemic species and more recently, de Lange & Rolfe (Citation2010) recognize seven species, two of which, B. humilis G.Simpson & J.S.Thomson and B. radicata Hook.f., they conclude are species aggregates. A recent phylogenetic analysis placed the New Zealand endemic B. humilis within the same clade as the Australian species which supports the idea that the New Zealand species have arrived by long distance dispersal from Australia (Brouillet et al. Citation2009).
Previous observations on the chromosomes of the New Zealand species reported a range of numbers from 2n=18 to 2n=90 (summarized in ). Brachyscome humilis is reported to have two chromosome numbers, 2n=36 and 2n=37 (Beuzenberg & Hair Citation1984; Dawson & Beuzenberg Citation2000); an interesting observation as examples of intraspecific chromosome number variation are relatively rare (< 3% of species) in the New Zealand flora (de Lange et al. Citation2008; Murray & de Lange Citation2013).
Table 1 Previously published chromosome numbers in the New Zealand species of Brachyscome.
Because B. humilis and B. radicata are considered to be species aggregates (de Lange & Rolfe Citation2010), in this paper we have analysed the chromosomes of plants matching the types of both species as well as morphologically divergent plants believed to be allied to them. In addition, we include several plants believed to be allied to B. sinclairii Hook.f. and B. montana G.Simpson. The taxonomic status of New Zealand species of Brachyscome is currently under review and this paper is a contribution toward the review describing the results of our cytological investigation of the genus.
Materials and methods
The plant material used in this study is listed in . All plants were collected as living specimens from the wild. Somatic chromosomes were obtained from root tip meristems that were pretreated with a saturated solution of paradichlorobenzene for c. 18 h at 4 °C, fixed in ethanol/acetic acid (3:1 v/v), hydrolysed in 1 M HCl for 8 min before staining with FLP orcein. A minimum of five cells per plant were observed and counted.
Table 2 Chromosome counts, collection details, Landcare Research Garden number and herbarium voucher number for the Brachyscome plants studied.
Results
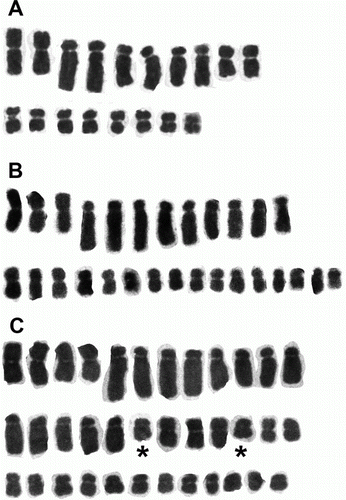
We found five different chromosome numbers in the 18 plants that we analysed. Twelve plants were diploid with 2n=18, one had 2n=28, three were tetraploids with 2n=36 (though one plant, the sole example of B. montana s.s. [G210/97, G] appeared to contain cells with 35 or 36 chromosomes), one was decaploid with 2n=90 (J) and one was decaploid but had two extra chromosomes (2n=92, K) ( and ). Karyotypes were constructed for three plants, the diploid B. longiscapa G.Simpson & J.S.Thomson (A), B. aff. sinclairii (G148/00, B) with 2n=28 and tetraploid B. humilis (G18306) (C). The diploid karyotype comprised eight small metacentric or sub-metacentric chromosomes and ten large, acrocentric chromosomes (Fig. 2A). All the diploid plants that we examined had a similar assortment of chromosomes. The majority of chromosomes of the B. aff. sinclairii plant (G148/00) can be arranged into sets of three presumed homologues with one additional small chromosome (Fig. 2B). In the tetraploid B. humilis most, although possibly not all, of the chromosomes can be arranged in sets of four, but as shown in C there appears to have been some chromosome rearrangement because not all presumed homologues can be arranged in groups of four. However, we appreciate that the placing of the chromosomes into groups of presumed homologues is somewhat subjective but it does appear in this plant that not all the chromosomes can be placed into groups of four. In B. radicata we have confirmed previous counts of 2n=90 in one plant and found 2n=92 in another that is allied to B. radicata. This latter plant is either an aneuploid or has two B chromosomes, we cannot confirm at this stage which of these possibilities is correct.
Figure 1 Mitotic metaphase in New Zealand Brachyscome. A, B. longiscapa 2n=18 (AK 233220). B, B. pinnata 2n=18 (G132/95). C, B.aff. sinclairii 2n=18 (G14292). D, B.sp. (indet.) 2n=18 (G212/97). E, B.aff. montana 2n=18 (G200/97). F, B. aff. sinclairii 2n=28 (G148/00). G, B. montana 2n=36 (G210/97). H, B. humilis 2n=36 (G18306). I, B. sp. (indet.) 2n=36. J, B. radicata 2n=90 (G66/9). K, B.aff. radicata 2n=92 (G497/98). Scale bar = 10 µm.
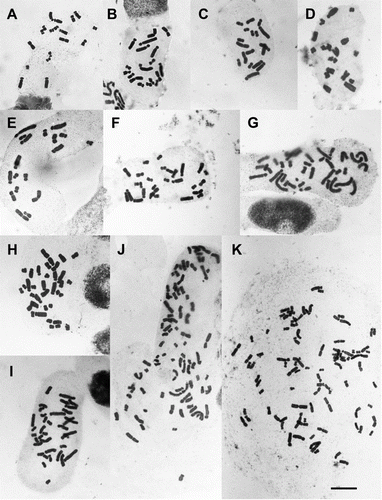
Figure 2 The karyotypes of three Brachyscome species of different ploidy levels. A, B. longiscapa 2n=18 (AK 233220). B, B. aff. sinclairii 2n=28 (G148/00). C, B. humilis 2n=36 (G18306). The chromosomes of B. humilis that do not appear to have obvious homologues are indicated by ★.
Discussion
We have confirmed previous chromosome counts for many of the New Zealand species of Brachyscome and found diploid numbers in two of the undetermined Brachyscome from Otago (G212/97 & G32/96) and one from Marlborough (G131/92). We found one tentatively new species currently allied to B. humilis (G263/98) had 2n=18, rather than the expected 2n=36 (Beuzenberg & Hair Citation1984; Dawson & Beuzenberg Citation2000). One plant morphologically allied to B. sinclairii had 2n=28 but the other plant had 2n=18, the same number as previously reported for B. sinclairii. In B. radicata, one plant had the expected number of 2n=90 but the other, which is similar to that species, had 2n=92. Neither 2n=28 nor 2n=92 have been reported previously in New Zealand species of the genus (). Our 2n=35/36 count for B. montana G.Simpson is the first for this species and the B.aff. montana plant from The Remarkables was diploid. Described by Simpson (Citation1945) B. montana was treated as ‘Incertae Sedis’ by Allan (Citation1961, p. 604–605) and the species was not discussed by Webb et al. (Citation1988) although it was accepted as distinct by Druce (Citation1993), Webb & Simpson (Citation2001) and de Lange & Rolfe (Citation2010). Of the two plants we counted, one (G210/97) comes from the type locality (Mt Cardrona, Central Otago) and matches Simpson's 1945 protologue as to description. The second plant (G200/97), from Doolan's Creek, The Remarkables, Central Otago is also a very close match to Simpson's description of B. montana but is clearly diploid. At this stage it is not clear whether there are diploid and tetraploid races of B. montana or whether the diploid plant represents another, potentially distinct segregate from B. montana. Central to this problem is that the type of B. montana appears to be missing (Allan Citation1961), and until the typification of B. montana is resolved any further taxonomic conclusions with respect to our diploid count would be premature.
Unlike in so many New Zealand native plants where chromosomes are small and often numerous, the chromosomes of Brachyscome are relatively large, in most cases relatively few in number and in addition individual chromosome morphology and size is variable. This makes basic karyotype analysis possible and this offers the potential to elucidate whether karyotype changes have accompanied speciation. All of our diploids appear to have the same basic karyotype and this is similar to a karyotype, presented as an idiogram, for a further undescribed entity (CHR 518417) from Ward Beach, Marlborough (Dawson & Beuzenberg Citation2000). Our undescribed plant from the Livingstone Mountains in Southland had a tetraploid number, the same number that we found in a plant, collected as B. humilis, from the Rock and Pillar Range, Otago. The karyotype of this Otago plant showed many chromosomes in groups of four, suggestive of autopolyploidy, though there are possibly some chromosomes in the complement that are not easily grouped in that way, thus suggesting that some structural rearrangements may have occurred associated with whole genome duplication. The plant with 2n=28 from Ward Beach appears to be an autotriploid, as those of its chromosomes that can be readily distinguished fall into groups of three apparent homologues, but it must also be aneuploid for one of the small chromosomes which we cannot differentiate unambiguously one from another. The plant of B.aff. radicata from the Umbrella Mountains also appears to be an aneuploid, although in this case of a decaploid, or to contain two B chromosomes. Aneuploids are not commonly found in natural plant populations but they are not unknown and a study of the rare Australian daisy, Rutidosis leptorrhynchoides F.Muell. as well as the New Zealand native Crassula ruamahanga A.P.Druce found extensive aneuploidy variation in several wild populations (Murray & Young Citation2001; de Lange et al. Citation2008).
The New Zealand species of Brachyscome, like their Australian congeners, are chromosomally variable showing evidence of both euploid and aneuploid variation. At this stage it is unclear whether any of this chromosome variation underlies the apparent taxonomic variation. The genus in New Zealand needs further detailed study at the population level to clarify the extent of chromosome variation and the application of molecular cytogenetic analyses to better understand genome evolution in the group. Further investigation is also needed to clarify the systematic position of the variants of the named species that we have studied.
Acknowledgements
We thank Nick Head, Cathy Jones and Neill Simpson for assistance in the field and David Purcell (Landcare Research, Lincoln Campus) for tending the plants examined in this study.
References
- Allan , HH . 1961 . Flora of New Zealand , 1085 Wellington : Government Printer .
- Beuzenberg , EJ and Hair , JB . 1984 . Contributions to a chromosome atlas of the New Zealand flora – 27. Compositae . New Zealand Journal of Botany , 22 : 353 – 356 . doi: 10.1080/0028825X.1984.10425266
- Brouillet , L , Lowrey , TK , Urbatsch , L , Karaman-Castro , V , Sancho , G , Wagstaff , S and Semple , JC . 2009 . “ Astereae ” . In Systematics, evolution, and biogeography of Compositae , Edited by: Funk , VA , Susanna , A , Stuessy , TF and Bayer , RJ . 589 – 629 . Vienna : International Association for Plant Taxonomy .
- Dawson , MI and Beuzenberg , EJ . 2000 . Contributions to a chromosome atlas of the New Zealand flora – 36. Miscellaneous families . New Zealand Journal of Botany , 38 : 1 – 23 . doi: 10.1080/0028825X.2000.9512671
- Druce AP 1993 . Indigenous vascular plants of New Zealand . Ninth revision. Unpublished checklist held at Landcare Research , Lincoln , , New Zealand .
- de Lange , PJ , Heenan , PB , Keeling , DJ , Murray , BG , Smissen , R and Sykes , WR . 2008 . Biosystematics and conservation: a case study with two enigmatic and uncommon species of Crassula from New Zealand . Annals of Botany , 101 : 881 – 899 . doi: 10.1093/aob/mcm294
- de Lange , PJ and Murray , BG . 2002 . Contributions to a chromosome atlas of the New Zealand flora – 37. Miscellaneous families . New Zealand Journal of Botany , 40 : 1 – 23 . doi: 10.1080/0028825X.2002.9512767
- de Lange , PJ , Murray , BG and Datson , PM. 2004 . Contributions to a chromosome atlas of the New Zealand flora – 38. Counts for 50 families . New Zealand Journal of Botany , 42 : 873 – 904 . doi: 10.1080/0028825X.2004.9512936
- de Lange , PJ and Rolfe , J . 2010 . New Zealand indigenous vascular plant checklist 2010 , 131 Wellington : New Zealand Plant Conservation Network .
- Murray BG , de Lange PJ 2013 . Contributions to a chromosome atlas of the New Zealand flora – 40. Miscellaneous counts for 36 families . New Zealand Journal of Botany 51 : 31–60 . doi: 10.1080/0028825X.2012.747969
- Murray , BG and Young , AG . 2001 . Widespread chromosome variation in the endangered grassland forb Rutidosis leptorrhynchoides F. Muell. (Asteraceae: Gnaphalieae) . Annals of Botany , 87 : 83 – 90 . doi: 10.1006/anbo.2000.1307
- Simpson , G. 1945 . Notes on some New Zealand plants and descriptions of new species (No. 4) . Transactions of the Royal Society of New Zealand , 75 : 187 – 202 .
- Smith-White , S , Carter , CR and Stace , HM . 1970 . The cytology of Brachycome I. The subgenus Eubrachycome: a general survey . Australian Journal of Botany , 18 : 99 – 125 . doi: 10.1071/BT9700099
- Watanabe , K , Yahara , T , Denda , T and Kosuge , K . 1999 . Chromosomal evolution in the genus Brachyscome (Asteraceae, Astereae): Statistical tests regarding correlation between changes in karyotype and habit using phylogenetic information . Journal of Plant Research , 112 : 145 – 161 . doi: 10.1007/PL00013869
- Webb , CJ and Simpson , MJA . 2001 . Seeds of New Zealand gymnosperms and dicotyledons , 428 Christchurch : Manuka Press .
- Webb , CJ , Sykes , WR and Garnock-Jones , PJ . 1988 . Flora of New Zealand IV. Naturalised pteridophytes, gymnosperms, dicotyledons , 1365 Christchurch : DSIR Botany Division .
- Weiss-Schneeweiss , H and Schneeweiss , GM . 2013 . “ Karyotype diversity and evolutionary trends in angiosperms ” . In Plant genome diversity, volume 2 , Edited by: Leitch , IJ , Greilhuber , J , Doležel , J and Wendel , JF . 209 – 230 . Wien : Springer-Verlag .