
Abstract
Tephra eruptions have significant long-lasting impacts on vegetation, and potentially explain extant vegetation patterns in volcanic landscapes. We quantified the effects of the AD 1655 Burrell Lapilli deposit, Mt Taranaki, on treeline vegetation. Where lapilli depth was 25–40 cm, a succession close to primary was initiated. Where lapilli depth was 5–25 cm, the canopy was opened, but some vegetation survived. Total tree basal area remains lower in affected vegetation (158 cf. 206 m2 ha−1), whereas total tree density is higher where a moderate disturbance stimulated regeneration (8433 cf. 6656 stems ha−1). Griselinia littoralis and Podocarpus cunninghamii dominate treeline vegetation (c. 51 and 39 m2 ha−1, respectively) within the lapilli distribution. Where disturbance was moderate, a cohort of Libocedrus bidwillii was also initiated. Weinmannia racemosa is absent from treeline vegetation within the lapilli distribution, despite being a dominant component elsewhere (basal area 48 m2 ha−1). Compositional patterns result from interspecific differences in morphology and resilience, as well as light, substrate and temperature tolerance. Light-demanding, cold-tolerant taxa were able to take advantage of the newly created open sites, whereas shade-tolerant, less hardy species lost their competitive advantage at the treeline elevation. The successional trajectory of the treeline vegetation has been set back and altered, and there is no evidence of convergence.
Introduction
Volcanic eruptions represent infrequent and unpredictable disturbance events that have the potential to significantly affect vegetation. During eruptions, vegetation can be modified or destroyed by volcanic processes, and new geological substrates may be created for plants to re-establish upon (Dale et al. Citation2005a). Of all the vegetation disturbances that may result from volcanic eruptions, tephra fall is the most widespread in the landscape (Antos & Zobel Citation2005). Tephra is fragmental material explosively ejected into the air by an eruption and, depending on the diameter of fragments, can be categorised as ash (< 2 mm diameter), lapilli (2–64 mm diameter) or blocks and bombs (> 64 mm diameter). Eruption plumes containing tephra are readily influenced by wind direction, with the erupted material being transported and deposited downwind. Tephra deposits are generally deepest in close proximity to the volcanic vent, and become progressively thinner with increasing distance from the source (Carey Citation2005). The effect of tephra on vegetation is primarily related to the depth (thickness) of the ejected material (Eggler Citation1963; Vucetich & Pullar Citation1963; Hotes et al. Citation2004; Antos & Zobel Citation2005; Tsuyuzaki & Hase Citation2005; Gomez-Romero et al. Citation2006).
There is a close link between vegetation burial by tephra and succession pattern and process, with burials being retrogressive events that initiate either primary and/or secondary succession, depending on the severity of the eruption and the proximity to the source. Various studies have indicated that emergent, canopy and senescent vegetation are most susceptible to damage by tephra, whereas juvenile, robust or vigorous individuals have better chances of survival and are able to exploit forest gaps, thus facilitating forest succession (Kent et al. Citation2001). Vucetich & Pullar (Citation1963) examined plant macrofossils in the volcanically active central North Island of New Zealand (Taupo Volcanic Zone), and attempted to determine critical tephra depths at which damage to vegetation would occur. They concluded that > 38 cm of tephra burial would result in complete destruction of trees, 20–38 cm in almost complete destruction, and 23–30 cm in partial destruction. On Mt St. Helens (Washington, USA), thinner 1–15 cm depositions of tephra were insufficient to kill canopy trees, but damage to the forest understory (i.e. shrubs, seedlings, groundcovers) could still be severe and influence the successional trajectory of the vegetation (Antos & Zobel Citation1985, Citation1986, Citation2005; Zobel & Antos Citation1997). Several confounding factors other than tephra depth also determine the effects that a tephra eruption will have on vegetation. These include differences in the stature, morphology and eco-physiology of the taxa involved, the availability of micro-sites for protection, tephra chemistry and temperature, and the occurrence of rain, snow and wind at the time of tephra deposition (Kent et al. Citation2001; Dale et al. Citation2005a). Antos & Zobel (Citation1987) commented that data on vegetation changes caused by volcanic eruptions were sparse and mostly anecdotal, but the 1980 eruption of Mt St. Helens stimulated numerous succession studies (Dale et al. Citation2005b), as have recent eruptions in Japan (Tsuyuzaki Citation1989, Citation1991; Nakashizuka et al. Citation1993; Titus & Tsuyuzaki Citation2003), and elsewhere (Clarkson Citation1990, Citation1998; Whittaker et al. Citation1992; Oner & Oflas Citation1997).
In the North Island of New Zealand, numerous geologically young volcanoes (Rangitoto Island, White Island/Whakaari, Mts Tarawera, Ruapehu, Ngauruhoe, Tongariro and Mt Taranaki/Egmont) provide sites for the study of both primary and secondary succession initiated by volcanic disturbance. The analysis of plant macrofossils and pollen preserved in sediments across the North Island has often been used to determine broad-scale vegetation changes associated with volcanic disturbance (Vucetich & Pullar Citation1963; McGlone et al. Citation1988; Lees & Neal Citation1993; Clarkson et al. Citation1988, Citation1995; Wilmshurst et al. Citation1997; Horrocks & Ogden Citation1998; Giles et al. Citation1999), though given that the lifespan of many New Zealand trees is potentially > 500 years, present-day spatial differences in forest composition and structure can also be related directly back to volcanic events (Clarkson Citation1990).
In this study, we quantified present-day treeline vegetation across a relatively recent (AD 1655) tephra depth gradient on the New Zealand volcano Mt Taranaki. We asked whether, and to what extent, tephra depth is associated with present-day differences in vegetation composition, structure and dynamics. We also sought to determine the critical tephra depth thresholds for plant survival and recovery and whether species' differential responses to damage by tephra, and interactions between key species after the event, may have shaped the current vegetation pattern.
Study area and vegetation
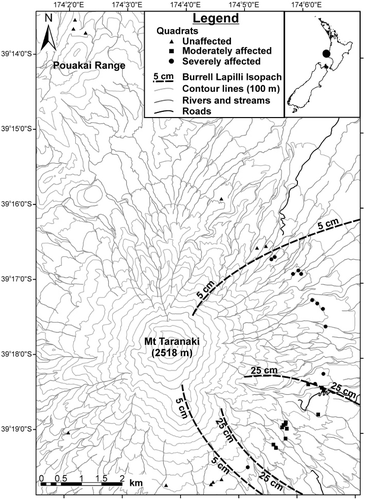
Mt Taranaki/Egmont is a dormant andesitic strato-volcano, which dominates the landscape of the Taranaki Region, North Island, New Zealand (39°18′S, 174°4′E). Mt Taranaki rises 2518 m above sea level (a.s.l.), and alongside the progressively older volcanic remnants of Pouakai, Kaitake and Paritutu, defines the Quaternary volcanic lineament known as the Taranaki Volcanic Succession. The present-day cone of Mt Taranaki has been built up by eruptions in the last 10,000 years, and since c. AD 1500, Mt Taranaki has had at least 10 distinct eruption events (Neall Citation1980; Neall et al. Citation1986; Platz et al. Citation2012); although nearly all occurred prior to the European occupation of New Zealand, and no accounts are known from the indigenous Māori people (Lowe et al. Citation2002). The present study focused on the AD 1655 Burrell Lapilli eruption. Druce (Citation1966) determined the year of the Burrell Lapilli eruption through dendrochronological dating of pre- and post-eruption trees, and was confident that the eruption occurred between AD 1654 and 1658, with the winter half of AD 1655 being the most probable date. The Burrell Lapilli deposit consists of up to 40 cm of loose white pumiceous blocks, lapilli and coarse ash (Topping Citation1972). Druce (Citation1966) and Topping (1972) mapped the extent and depth of the Burrell Lapilli over Mt Taranaki, producing almost identical isopach maps. These maps indicate that tephra fallout occurred over an area of c. 150 km2 to the east–southeast of the summit vent on account of the wind direction at the time, with a well-defined dispersal axis present from which the depth of lapilli gradually decreases from 40 to 0 cm. None of the other recent eruptions from Mt Taranaki have directly influenced the study site within the Burrell Lapilli distribution, other than the AD 1655 Burrell Ash and AD 1755 Tahurangi Ash, which are both distributed more or less evenly over the study area (Druce Citation1966). Soils on Mt Taranaki are predominantly recent/raw volcanic soils developed from andesitic tephra, and within the study area, Burrell Ash and Tahurangi Ash are the main parent materials (Tonkin Citation1970).
Mt Taranaki has a temperate maritime climate, with continually alternating calm and stormy weather conditions. Annual rainfall near the treeline elevation is c. 6000 mm, with an average summer temperature of 12–13 °C and an average winter temperature of 4–5 °C (Clarkson Citation1981). The slopes of Mt Taranaki are currently clothed with largely intact natural vegetation which has been described by Druce (Citation1966, Citation1974, Citation1976a, Citationb) and Clarkson (Citation1977, Citation1981, Citation1986). The most obvious vegetation pattern relates to altitude, with an uninterrupted sequence from lowland forest through to alpine vegetation represented. The current study investigated treeline vegetation specifically. Here ‘treeline’ refers to the narrow belt of transitional vegetation between upper montane forest and subalpine shrubland, whereby the canopy is typically between 2 and 3 m in height and sporadically interrupted with taller emergent trees. On Mt Taranaki, this treeline occurs at c. 1050 m a.s.l. (Efford Citation2012) and the main trees represented are Podocarpus cunninghamiiFootnote1 and Griselinia littoralis, either with or without Libocedrus bidwillii, overtopping tree-sized shrubland species Brachyglottis elaeagnifolia, Pseudopanax colensoi and Raukaua simplex. Treeline composition here is comparable with many areas in the North Island (e.g. Pouakai Range, Mt Hauhungatahi, north side Mt Tongariro, south and southwest Ruahine Range) where Nothofagus is absent (Druce Citation1966). Below the treeline exists montane forest (600–1000 m a.s.l.) characterised by a c. 10 m high canopy of Weinmannia racemosa and lesser amounts of G. littoralis, intermixed with emergent P. cunninghamii and L. bidwillii. Above the treeline, vegetation stature gradually decreases to c. 2.5 m by 1100 m a.s.l., and crowns merge closely to produce a compact hedge-like canopy. Between 1100 and 1400 m a.s.l., tree species are replaced in the canopy by shorter shrub species including B. elaeagnifolia, P. colensoi, R. simplex and Hebe stricta var. egmontiana. Volcanic eruptions and associated processes on Mt Taranaki are thought to have destroyed c. 6700 ha of vegetation and damaged an equivalent area in the last c. 500 years (Druce Citation1966). Both Druce (Citation1966) and Clarkson (Citation1981, Citation1986, Citation1990) speculated that the contributions of W. racemosa, G. littoralis, P. cunninghamii and L. bidwillii may vary across the Burrell Lapilli depth gradient, although these differences have never been comprehensively quantified until now.
Methods
Data collection
Treeline vegetation was measured across the extent of the AD 1655 Burrell Lapilli deposit with 36 permanently marked 10 × 15 m quadrats (). Nested quadrat minimal-area checks had previously been used to determine an adequate quadrat size of 150 m2 for surveying treeline vegetation in the study area (Clarkson Citation1981; J.T. Efford, unpubl. data). A stratified random sampling design was used to position the quadrats across three Burrell Lapilli depth categories (Druce Citation1966) at accessible treeline positions with 0–5, 5–25 and 25–40 cm lapilli depth. Twelve quadrats were located in each lapilli depth category. The 0–5 cm category included three quadrats from the treeline of the adjoining Pouakai Range, an area unaffected by the Burrell eruption. For ease of interpretation, the Burrell Lapilli depth categories of 0–5, 5–25 and 25–40 cm may henceforth be referred to as ‘unaffected’, ‘moderately affected’ and ‘severely affected’ areas, respectively; with the ‘eruption zone’ referring to the latter two categories only. For this study, ‘trees’ are defined as individuals > 2 cm diameter at ground height (dgh); tree ‘saplings’ are individuals < 2 cm dgh, but > 50 cm tall; and tree ‘seedlings’ are individuals 5–50 cm tall. In each 10 × 15 m quadrat, all trees were recorded by species and measured for dgh. The dgh of snags (standing dead trees; > 10 cm dgh) was also recorded. The physiognomy of the snags suggested they were either P. cunninghamii or L. bidwillii, however, because of their decayed state it was not always possible to distinguish the two, so these were grouped for analysis. In a randomly selected quarter of each quadrat (7.5 × 5 m), all tree and shrub saplings and seedlings were recorded by species and counted. Ground cover species in each quadrat were assigned a cover class based on the proportion of live foliage < 1 m, and all epiphyte species were recorded. A list of the vascular species present in each quadrat was also compiled.
Data analysis
Tree species and snag basal area and density were calculated first for each quadrat, and then average (mean) values and standard deviations (SD) were obtained for each of the three lapilli depth categories. Diameter frequency distributions (histograms) were derived for key species in each lapilli depth category from pooled quadrat dgh measurements; histograms used seedlings as the smallest size class, followed by saplings, then a 2–10 cm dgh class, and progressive 10 cm dgh increments thereafter.
Non-metric multidimensional scaling (NMS), an indirect ordination technique, was performed in PC-ORD Version 6.0 (McCune & Mefford Citation2011) to graphically arrange the quadrats according to similarities in species composition and abundance alone (not constraining the axes to secondary variables). In the ordination analysis, the primary data matrix comprised the density values of 30 tree species found in 36 quadrats. No data transformations were considered necessary. The secondary matrix was composed of site variables specific to each quadrat, and included thickness of the Burrell Lapilli, elevation, slope and aspect. Site aspect azimuths were assigned to one of four classes, 1 for north (316–45°), 2 for east (46–135°), 3 for west (226–315°) and 4 for south (135–225°). Burrell Lapilli thickness was recorded both quantitatively (0–40 depending on lapilli thickness in cm according to Druce's Citation1966 isopach map) and categorically (using the three Burrell Lapilli depth categories previously described). Sorenson (Bray–Curtis) distance measure was used in the ordination and the analytical recommendations of McCune & Grace (Citation2002) relating to stress value, stability criterion and number of iterations were followed. After ordination of the quadrats based on species composition and abundance, NMS independently assessed the relationship of the site variables to the species composition and abundance. A three-dimensional solution was selected and the ordination was rotated to obtain maximum correlation between axis 1 and Burrell Lapilli depth. Pearson correlation coefficients (r) calculated for each species and site variables, in relation to primary ordination axes, were used to assess variation in species composition and abundance across site variable gradients (Keith et al. Citation2010).
A permutational multivariate analysis of variance (perMANOVA) and post-hoc pair-wise comparison were used to test for significant differences in species composition and abundance between the three lapilli depth categories (Anderson Citation2001). The perMANOVA was conducted in PC-ORD using a Sorenson (Bray–Curtis) distance measure and the same primary data matrix used for the NMS ordination.
Results
NMS ordination
Some 57 vascular taxa were identified in the survey of treeline vegetation across the Burrell Lapilli deposit. All taxa were indigenous, constituting a mix of upper montane forest and subalpine shrubland species. Burrell Lapilli depth was the strongest influence on treeline vegetation composition (, ). Axis 1 represented most of the variation in composition (R2 = 0.42), and was strongly correlated to the Burrell Lapilli depth variable (r = 0.80). Topographic slope was also moderately correlated with axis 1 (r = 0.36). Axis 2 and axis 3 represented the remaining variation (R2 = 0.36; 0.15), and were both correlated with elevation (r = −0.53; −0.45). No correlation between the ordination axes and quadrat aspect was apparent. The three Burrell Lapilli quadrat groups (severely affected, moderately affected, unaffected) were largely separated from one another in ordination space, with minimal overlap occurring between the groups. Several species displayed moderate (Pseudopanax colensoi, Alseuosmia macrophylla, Cyathea smithii) or significant (Coprosma grandifolia, G. littoralis, Coprosma tenuifolia, Melicytus lanceolatus) positive correlations with axis 1, suggesting that these species have become more abundant in treeline vegetation affected by the Burrell Lapilli (). Conversely, significant negative correlation with axis 1 (Pseudowintera colorata, Myrsine salicina) suggests other species are suppressed from the Burrell eruption zone. A lack of correlation between axis 1 and several key species shown to differ in relation to lapilli depth results from nonlinear relationships.
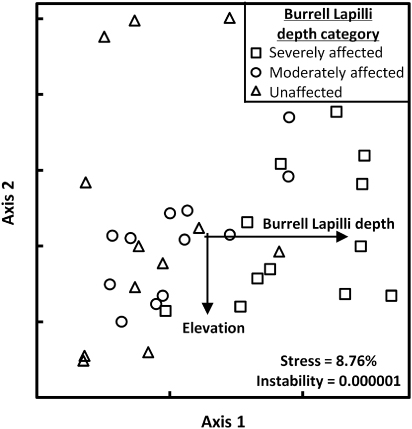

Table 1 Correlation (r) of species and site variables with NMS ordination axes.
perMANOVA
A perMANOVA provided a statistical test of species composition and abundance across the three Burrell Lapilli depth categories and indicated a significant difference in vegetation (). Pair-wise comparisons distinguished that significant differences existed between unaffected and severely affected vegetation, as well as between moderately affected and severely affected vegetation.
Table 2 perMANOVA on treeline species composition and abundance across three Burrell Lapilli depth categories, Mt Taranaki. Highly significant P values (P < 0.001) are given in bold.
Vegetation composition
Basal areas and densities of dominant tree species and snags are presented in . Across the three Burrell Lapilli depth categories, total basal area was greatest in the unaffected area (206 m2 ha−1), with progressively lower totals found across the moderately affected (180 m2 ha−1) and severely affected (158 m2 ha−1) areas. Total tree density was highest in the moderately affected area (8433 stems ha−1), compared with the severely affected (6972 stems ha−1) and unaffected (6656 stems ha−1) areas, which both exhibited similar densities. The contributions of several potential treeline canopy or emergent species were found to vary considerably in relation to Burrell Lapilli thickness. Using basal area as a measure of species dominance, treeline vegetation unaffected by the Burrell Lapilli was characterised by G. littoralis (56 m2 ha−1) and W. racemosa (48 m2 ha−1), with lesser amounts of L. bidwillii (31 m2 ha−1) and P. cunninghamii (25 m2 ha−1). In moderately affected areas, G. littoralis (51 m2 ha−1), P. cunninghamii (43 m2 ha−1) and L. bidwillii (26 m2 ha−1) were most dominant, while W. racemosa (0.1 m2 ha−1) was essentially absent. In the severely affected areas, G. littoralis (51.1 m2 ha−1) and P. cunninghamii (35.1 m2 ha−1) were dominant, while W. racemosa and L. bidwillii were absent from the vegetation (). Snags contributed to the vegetation in all three lapilli depth categories. Basal area of snags was greatest in the severely affected category (27.3 m2 ha−1) and decreased with lapilli depth. Conversely, the density of snags was the lowest in the severely affected category (44 stems ha−1) and did not correlate with lapilli depth; the moderately affected area had the highest snag density (122 stems ha−1). Results show the high snag basal area in the severely affected category is a result of large diameter snags, in contrast to the moderately affected area, which had numerous smaller diameter snags.
Table 3 Basal area (± SD) and density (± SD) values for treeline species across three Burrell Lapilli depth categories, Mt Taranaki.
A number of other less abundant tree and shrub species also displayed notable trends across the lapilli depth gradient. Fuchsia excorticata and Aristolelia serrata, for example, were only found within the severely affected area. Pseudopanax colensoi, C. grandifolia, C. tenuifolia, Carpodetus serratus and C. smithii were all more abundant in the severely affected area than elsewhere. The basal area of L. bidwillii and P. cunninghamii snags was greatest in the severely affected area. Brachyglottis eleagnifolia and H. stricta var. egmontiana were also more common in the eruption zone than the unaffected area. By contrast, M. salicina was only found in unaffected areas. Pseudowintera colorata, an important understory component across all lapilli depth categories, was least abundant in the severely affected area.
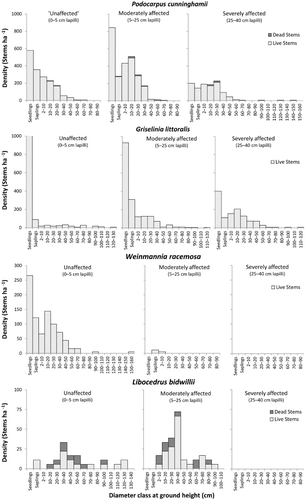
Diameter frequency distributions
The diameter frequency distributions (population structures) displayed by canopy species were variable across each of the lapilli depth categories (), and provide further insight into species response to the Burrell eruption. The area unaffected by the eruption serves as a reference point for populations that are unlikely to have had any recent influences from wide-scale disturbances. In the unaffected area, P. cunninghamii and W. racemosa both exhibited reverse-J type (continuous recruitment) population structures with high numbers of seedlings and progressively fewer stems in the larger diameter classes. Griselinia littoralis was represented by a range of diameters in the unaffected area (an all-aged population), but the majority of seedlings appeared to be failing to reach the subsequent sapling size class. Libocedrus bidwillii had a sporadic population structure in the unaffected area, which included a number of dead stems and an absence of seedlings. In the moderately affected area, P. cunninghamii and L. bidwillii both displayed cohort distributions, although only P. cunninghamii had representation in the seedling and sapling classes. Griselinia littoralis displayed a reverse-J distribution in the moderately affected area, with a higher level of seedling success apparent than in the unaffected area. Weinmannia racemosa was only represented by a small number of saplings and juvenile trees in the moderately affected area. In the severely affected area, W. racemosa and L. bidwillii were both absent, while P. cunninghamii and G. littoralis displayed cohort distributions, with several of the P. cunninghamii snags being > 1.0 m diameter.
Discussion
Response of treeline vegetation to tephra damage
As a result of recent volcanic disturbances, the majority of the treeline around Mt Taranaki currently occurs at a lower elevation than on the adjoining Pouakai Range, with upper altitudinal limits of species often being higher on Pouakai (Clarkson Citation1990). The treeline position will slowly continue to advance up the slopes of Mt Taranaki, until it is again inhibited by another eruption, or species finally reach their potential upper altitudinal limits. Suppression of the treeline position has also been reported on Mt St Helens. Prior to the 1980 eruption there, the treeline was still advancing up the mountain in response to an eruption in AD 1800, and it was again suppressed considerably by the 1980 eruption (Swanson et al. Citation2005). More than 357 years after the Burrell eruption, structural and compositional differences were strongly evident in treeline vegetation across the Burrell Lapilli gradient on Mt Taranaki. This sector of the mountain also received ash deposition from the Newall and Tahurangi eruptions. The Newall event preceded the Burrell eruption by c. 150 years and resulted in c. 5 cm of ash being uniformly deposited over the study site (Druce Citation1961). This event would have increased soil nutrients but was unlikely to have had devastating or lasting effects on the vegetation, although minor canopy disturbance and seedling suppression could have decreased vegetation resilience to the effects of the Burrell Lapilli. The Tahurangi Ash was distributed across the upper slopes of the mountain c. 100 years after the Burrell event (Platz et al. Citation2012), where it has contributed to the formation of the current soil (Clarkson Citation1986). Although this ash may have had some impact on vegetation, given its concentric distribution, the effects would have probably been equivalent across the study area.
Treeline vegetation in the affected areas is yet to recover to the pre-eruption volumes indicated outside the Burrell eruption zone, with lapilli depth correlating with the numbers of trees destroyed by the eruption and the rate at which the vegetation has been able to recover. Large snags were more dominant in the severely affected area of the treeline. Natural senescence or possum damage cannot be fully ruled out as possible explanations for the trees deaths, however, their larger size compared with those in less severely affected areas, and lack of obvious root flanges, suggests these pre date, and were deleteriously affected by the Burrell eruption. Currently, total tree basal area is greatest outside the lapilli distribution, and decreases with lapilli depth. Total tree density was found to be greatest where the vegetation was moderately affected by the eruption (5–25 cm lapilli). Tree regeneration initiated by the eruption may, therefore, have been most prolific where lapilli damage opened the canopy to the extent that existing seedlings and saplings were released, or where substrate remained favourable for a colonisation event. Greater lapilli depths would have also opened the canopy, but few survivors would have remained and the substrate may have been unsuitable for new colonists for some time. Self-thinning is yet to return the total tree densities to pre-eruption stocking rates, with the successional trajectory of the vegetation having been set back. Disturbance events in forests, particularly those that open the canopy, are known to create a wave of regeneration resulting in an increase in stem densities (Denslow Citation1995). Canopy damage inflicted by 5–15 cm of tephra during the 1980 eruption of Mt St. Helens, for example, resulted in the density of tree seedlings increasing twofold, with much greater seedling establishment and survival than in the original undisturbed forest (Antos & Zobel Citation1986).
Differential species responses to damage
Vegetation would have responded variably across the Burrell Lapilli gradient following the eruption of Mt Taranaki, with mosaics created by a combination of the survival, death and arrival of species. Compositional patterns are thus likely to be a result of interspecific differences in, for example, morphology, resilience, demography and light and substrate requirements. Weinmannia racemosa, an abundant upper montane species over much of Mt Taranaki and at the treeline on Pouakai Range (Clarkson Citation1986), was shown to be absent from treeline vegetation severely affected by the eruption, and represented only by juveniles in the moderately affected areas. Previous research has determined that at lower elevations on Mt Taranaki, W. racemosa proliferated following eruption events due to its ability to resprout epicormically, and the copious amount of light wind-dispersed seed it produces (McGlone et al. Citation1988; Clarkson Citation1990). Weinmannia racemosa has also displayed a similar recovery following vegetation damage by the AD 1886 eruption of Mt Tarawera (Nicholls Citation1963; Clarkson & Clarkson Citation1983). Conversely, in treeline areas most affected by the Burrell eruption on Mt Taranaki, W. racemosa has not displayed this success. Species approach their upper-altitudinal limits at the treeline, and in this instance, the competitive advantage of W. racemosa to recover from disturbance appears to have been lost to more cold-tolerant species. On Mt Taranaki, P. cunninghamii, G. littoralis and L. bidwillii all display higher altitudinal limits than W. racemosa (Clarkson Citation1986), and thus have better exploited the eruption event at treeline. In parallel with Mt Taranaki, L. bidwillii was also able to dominate upper montane forest at the expense of W. racemosa following the AD 232 Taupo eruption (Hogg et al. Citation2011) on Mt Hauhungatahi, because L. bidwillii was more tolerant of the cooler conditions associated with exposure (Horrocks & Ogden Citation1998). Simulation of the post-Taupo vegetation dynamics on Mt Hauhungatahi using a forest landscape model also suggested a major shift in the montane forest composition, with initial L. bidwillii dominated stands being progressively replaced by W. racemosa following 400–700 years, when shade became a more limiting factor (Thrippleton et al. Citation2014). Myrsine salicina has also been excluded from treeline vegetation affected by the Burrell eruption, although it is quite common outside the distribution of lapilli. Myrsine salicina only appears in late-successional forest (Smale & Smale Citation2003), and is usually more prevalent on nutrient-depleted soils (B. D. Clarkson, University of Waikato, pers. obs.); the eruption has thus set back the successional trajectory of affected vegetation, with M. salicina yet to fully establish. Pseudowintera colorata was also found to be less successful in the severely affected areas, a pattern paralleled on Mt Tarawera where P. colorata is still rare in forest affected by the AD 1886 eruption (B. D. Clarkson, University of Waikato, pers. obs.).
Libocedrus bidwillii was suppressed from treeline vegetation, but in severely affected areas only. In moderately affected areas, it has been more successful with cohort populations being initiated by the eruption. Libocedrus bidwillii periodically establishes in even-aged stands following massive disturbance (Ogden et al. Citation2005), with seedlings being intolerant of shade and unable to regenerate below a closed canopy (Efford Citation2012). The absence of L. bidwillii from severely affected areas confirms that it was deleteriously affected by the Burrell eruption, but it is not clear why it has not responded with a typical wave of regeneration (Veblen & Stewart Citation1982; Boase Citation1988) such as that seen in the moderately affected area. Some possible explanations include that: 1. the deep lapilli substrate was unsuitable for L. bidwillii seedling establishment, 2. competitive exclusion by other light-demanding pioneer species occurred, 3. prevailing southerly winds in the severely affected southeastern side of the mountain inhibited L. bidwillii seed dispersal into the devastated areas, or 4. the timing of the eruption did not coincide with seed availability. The emergent physiognomy of L. bidwillii at the treeline would have resulted in it suffering the full impact of the lapilli shower, whereas species below the canopy may have been afforded some protection. Most emergent L. bidwillii at the time of the eruption never recovered from the stripping of foliage by lapilli, with some still remaining as decayed snags in the vegetation (Druce Citation1966; Efford Citation2012). Other studies have reported that L. bidwillii does not recover well from browsing damage by the introduced possum (Rogers Citation1997; Husheer Citation2006), further evidence that it may have struggled to recover from the mechanical damage inflicted by lapilli. Podocarpus cunninghamii also grows as an emergent in treeline vegetation, but is capable of more rapid foliar recovery than L. bidwillii following possum browse (Rogers Citation1997; Husheer Citation2006). Following the deposition of the Taupo tephra on Mt Hauhungatahi, L. bidwillii increased rapidly after the eruption in areas where tephra was < 9 cm thick, which is similar to the pattern on Mt Taranaki. At Mt Hauhungatahi, this was attributed to an increase in canopy gaps (created by the eruption), which permitted the light-demanding L. bidwillii to proliferate (Horrocks & Ogden Citation1998). Violent rainstorms which cause landslides can also be associated with large volcanic eruptions, and they too provide ideal sites for L. bidwillii colonisation (Boase Citation1988).
In treeline areas most severely affected by the Burrell eruption, P. cunninghamii and G. littoralis have become dominant. Perhaps not coincidently, both these species have bird-dispersed seed; as prevailing winds could have greatly inhibited seed transport into the area most severely affected by the eruption. Another possible explanation for the success of G. littoralis is related to its ability to colonise epiphytically. Elevated snags and debris would have been available immediately after trees were killed by the eruption, and the current growth form of many G. littoralis suggests that they have originated epiphytically. Clarkson (Citation1981) also noted that G. littoralis had been establishing epiphytically on senescent L. bidwillii on Mt Taranaki, thereafter developing their own supporting stems to remain as canopy dominants. Griselinia littoralis is the sole coloniser of forest gaps in the montane zone at Waihaha and prefers epiphytic establishment (Smale & Smale Citation2003). Senescent G. littoralis commonly replaces itself in situ at Mt Pureora, whereby seedlings establish epiphytically in crowns of parent trees, thus prolonging dominance (Smale & Kimberley Citation1993). A reciprocal regeneration cycle between G. littoralis and P. cunninghamii similar to that discussed by Lusk & Ogden (Citation1992) could also be occurring on Mt Taranaki, with G. littoralis capturing gaps formed by P. cunninghamii windfall, the latter, in turn, regenerating beneath the thinning crowns of older G. littoralis. On Mt Tarawera vegetation recovery has been slow, with P. cunninghamii only just beginning to colonise c. 110 years after the AD 1886 eruption, with facilitation by other species including G. littoralis thought to be important factors aiding its establishment (Clarkson & Clarkson Citation1995). Estimated ages for P. cunninghamii within the study area suggest it did not colonise immediately after the eruption, instead requiring a period of c. 150 years to establish (Efford Citation2012); probably time for suitable substrate to form as some lapilli was removed by erosion and soil developed.
Vegetation succession
Considering both the critical tephra thicknesses reported by Vucetich & Pullar (Citation1963), Tsuyuzaki (Citation1989) and Dale et al. (Citation2005b), and the results of the current research, it is likely that where the effects of the Burrell Lapilli were most severe (i.e. 25–40 cm lapilli), the large majority of trees and shrubs were killed, and a succession close to primary was initiated. A suite of species found to be more common in treeline vegetation within the Burrell eruption zone (Brachyglottis elaeagnifolia, H. stricta var. egmontiana, Pseudopanaxcolensoi, Raukaua simplex, Coprosma spp., Carpodetus serratus, Fuchsia excorticata, Aristolelia serrata) are mostly subalpine shrubland vegetation constituents above treeline (Clarkson Citation1986) or seral light-demanding taxa. Pollen analysis of sediments from the eastern flank of Mt Taranaki suggest that seral shrub species, particularly Coriaria arborea var. arborea, increased in abundance following the Burrell eruption (Lees & Neall Citation1993). Shrubland vegetation is essentially a successional precursor to upper montane/treeline forest on the mountain (Boase Citation1988), and the Burrell eruption disturbance has thus acted to suppress the elevation of the treeline to an extent, by setting back and retarding the rates of species succession. Where the Burrell Lapilli depth approached 40 cm, compositional and structural differences are still markedly apparent in the treeline vegetation. Given the time that has passed without convergence it is likely these differences will persist. Similarly, during the AD 232 Taupo eruption (Hogg et al. Citation2011), < 10 cm of tephra was thought to have damaged vegetation via mechanical stripping of branches and foliage, with the crowns of canopy and emergent trees being most affected, and mechanical damage exacerbated further with chemical damage by the acidic tephra. Within 200 years of the eruption, revegetation of areas totally overwhelmed was completed, and post-eruption forests were similar to pre-eruption forests (Wilmshurst & McGlone Citation1996). However, closer to the source of the eruption, on the Hauhungaroa Range, tephra depth approached 50 cm and recovery of vegetation was slower, taking up to 450 years. Unlike the more distal sites, vegetation composition was changed more permanently, with, for example, P. cunninghamii becoming abundant and L. bidwillii becoming rare after the Taupo eruption (Clarkson et al. Citation1985). Seedling composition on the forest floor also changed following the addition of tephra on Mt St. Helens; a result of a partially intact tree canopy with a drastically altered substrate. Often following a disturbance, species not previously present in the vegetation will invade, but on Mt St. Helens, despite nearly 100% plant death in some locations, few species absent from the pre-disturbance forest established. Instead, there was a compositional divergence of the existing species, with some recovering rapidly while others failed to increase. This resulted in a complex mosaic of survival over the landscape (Dale et al. Citation2005a). In the decades after the event, this compositional shift has not been towards pre-disturbance conditions (Antos & Zobel Citation1986, Citation2005). The treeline vegetation on Mt St. Helens is now an unusual mix of conifers and hardwoods, which differs considerably from adjoining ranges which were not affected by such recent volcanic disturbance (Swanson et al. Citation2005).
The introduction of browsing mammals (goats, Capra hircus; brush-tailed possums, Trichosurus vulpecula) to Mt Taranaki has also had an effect on the successional trajectory of the vegetation in the study area, reducing the vigour and abundance of palatable species while promoting the proliferation of less palatable types (Clarkson Citation1990; Husheer Citation2006). Goats were first recorded in the park in 1910, with numbers peaking during the 1960s (New Zealand Forest Service 1975; Forsyth et al. Citation2003) when damage to the vegetation became so severe that ‘dead areas’ as great as 5 ha developed in lower altitude forest (Clarkson Citation1990). In the upper altitude forest, effects were not as great, however, compositional changes were still apparent, with depletion of G. littoralis, F. excorticata, P. colensoi and R. simplex, whereas the less palatable shrub P. colorata, as well as grasses and ferns became more abundant. Plant species capable of epiphytic regeneration such as W. racemosa and G. littoralis may have partially avoided goat browse, and this also has potential to change the vegetation profile; historically these species may have had much higher success establishing on the forest floor. Many plant species (e.g. W. racemosa, P. cunninghamii, L. bidwillii, F. excorticata, P. colensoi, M. robusta and R. simplex) present in the study area are also vulnerable to possum browse and, as elsewhere in New Zealand (Smale & Smale Citation2003), possums represent a contemporary influence on vegetation succession. Locally, heavy possum browse of W. racemosa occurred where Pseudopanax spp., F. excorticata and A. serrata were abundant and provided food variety (Clarkson Citation1990). This may have further confounded W. racemosa recovery in the areas severely affected by the Burrell eruption, where these palatable species were more dominant. Significant reductions in browsing mammal numbers recently occurred, primarily a result of intensive pest control beginning in the 1970s, including recent aerial broadcasting of 1080, and this led to dramatic recovery of vegetation in some areas (Clarkson Citation1990, Citation2011; Husheer Citation2006). However, despite pest control W. racemosa continued to decrease in tree density (Husheer Citation2006). Weinmannia racemosa could potentially regain dominance at the treeline within the eruption zone, although given the considerable time that has elapsed this is improbable. Herbivore pressure, not present prior to human arrival, coupled with W. racemosa having lower climatic tolerances compared to other species (e.g. L. bidwillii, P. cunninghamii), likely limits its recovery at the treeline within the Burrell eruption zone.
Our research has quantified differences in treeline composition and structure directly resulting from the AD 1655 Burrell Lapilli distribution. Emergent species appear to have been the most heavily affected by the eruption, and where tephra depth approached 40 cm, it is likely that almost all vegetation was killed, permitting seral and light demanding taxa to proliferate. Although the treeline position is expected to advance up the mountain until species reach their upper altitudinal limits, the successional trajectory of vegetation has been set back and altered by the eruption to the extent that differences may persist indefinitely and convergence is unlikely.
Acknowledgements
We thank the Department of Conservation for Permission to undertake this study on DOC-administered land. We also thank the George Mason Charitable Trust and the Environmental Research Institute, University of Waikato, for providing funding for this project, and two anonymous reviewers for providing constructive comments on an earlier draft of the manuscript. This research is dedicated to the memory of Anthony (Tony) Peter Druce, for his pioneering research on the vegetation of Mt Taranaki.
Notes
1. Nomenclature follows Ngā Tipu o Aotearoa (http://nzflora.landcareresearch.co.nz/).
References
- Anderson MJ 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecology 26: 32–46.
- Antos JA, Zobel DB 1985. Recovery of forest under-stories buried by tephra from Mt St. Helens. Vegetatio 64: 103–111. 10.1007/BF00044786
- Antos JA, Zobel DB 1986. Seedling establishment in forests affected by tephra from Mt St. Helens. American Journal of Botany 73: 485–499. 10.2307/2444253
- Antos JA, Zobel DB 1987. How plants survive burial: a review and initial response to tephra from Mt St. Helens. In: Bilderback DE ed. Mt St. Helens 1980: botanical consequences of an explosive eruption. California, University of California Press. Pp. 246–261.
- Antos JA, Zobel DB 2005. Plant responses in forests of the tephra-fall zone. In: Dale VH, Swanson FJ, Crisafulli CM eds. Ecological responses to the 1980 eruption of Mt St. Helens. New York, Springer. Pp. 47–58.
- Boase MR 1988. Succession of sub-alpine vegetation to Libocedrus bidwillii forest on a landslide-initiated chronosequence, Mt Taranaki, New Zealand. Unpublished MSc thesis. Hamilton, University of Waikato.
- Carey SN 2005. Understanding the physical behaviour of volcanoes. In: Marti J, Ernst G eds. Volcanoes and the environment. Cambridge, Cambridge University Press. Pp. 2–54.
- Clarkson BD 1977. Vegetation change along an altitudinal gradient, Mt Egmont, New Zealand. Unpublished MSc thesis. Hamilton, University of Waikato.
- Clarkson BD 1981. Vegetation studies in the Taranaki Land District, New Zealand. Unpublished PhD thesis. Hamilton, University of Waikato.
- Clarkson BD 1986. Vegetation of Egmont National Park, New Zealand. Wellington, Department of Scientific and Industrial Research.
- Clarkson BD 1990. A review of vegetation development following recent (<450 years) volcanic disturbance in North Island, New Zealand. New Zealand Journal of Ecology 14: 59–71.
- Clarkson BD 1998. Vegetation succession (1967–89) on five recent montane lava flows, Mauna Loa, Hawaii. New Zealand Journal of Ecology 22: 1–9.
- Clarkson BD 2011. Ecological heritage in the Taranaki Region. In: Ruru J, Stephenson J, Abbott M eds. Making our place: exploring land-use tensions in Aotearoa New Zealand. Dunedin, Otago University Press. Pp. 240.
- Clarkson BR, Clarkson BD 1983. Mt Tarawera: 2. Rates of change in the vegetation and flora of the high domes. New Zealand Journal of Ecology 6: 107–119.
- Clarkson BR, Clarkson BD 1995. Recent vegetation changes on Mount Tarawera, Rotorua, New Zealand. New Zealand Journal of Botany 33: 339–354. 10.1080/0028825X.1995.10412961
- Clarkson BD, Clarkson BR, McGlone MS 1985. Vegetation history of some west Taupo mires. In: Veale B, Innes J eds. Ecological research in the central North Island volcanic plateau region, Proceedings of a New Zealand Forest Service workshop. Rotorua, Forest Research Institute. Pp. 34–37.
- Clarkson BR, McGlone MS, Lowe DJ, Clarkson BD 1995. Macrofossil and pollen representing forests of the pre-Taupo volcanic eruption (c. 1850 yr BP) era at Pureora and Bennydale, central North Island, New Zealand. Journal of the Royal Society of New Zealand 25: 263–281. 10.1080/03014223.1995.9517489
- Clarkson BR, Patel RN, Clarkson BD 1988. Composition and structure of forest overwhelmed at Pureora, central North Island, New Zealand, during the Taupo eruption (c. AD 130). Journal of the Royal Society of New Zealand 8: 417–436. 10.1080/03036758.1988.10426466
- Dale VH, Delgado-Acevedo J, MacMahon J 2005b. Effects of modern volcanic eruptions on vegetation. In: Marti J, Ernst G eds. Volcanoes and the environment. United Kingdom, Cambridge University Press. Pp. 227–249.
- Dale VH, Swanson FJ, Crisafulli CM 2005a. Disturbance, survival and succession: understanding ecological responses to the 1980 eruption of Mt St. Helens. In: Dale VH, Swanson FJ, Crisafulli CM eds. Ecological responses to the 1980 eruption of Mt St. Helens. New York, Springer. Pp. 3–12.
- Denslow JS 1995. Disturbance and diversity in tropical rain forests: the density effect. Ecological Applications 5: 962–968. 10.2307/2269347
- Druce AP 1961. Mountain vegetation of the North Island. New Zealand Soil News 3: 95–107.
- Druce AP 1966. Tree-ring dating of recent volcanic ash and lapilli, Mt Egmont. New Zealand Journal of Botany 4: 3–41. 10.1080/0028825X.1966.10443951
- Druce AP 1974. Some notes on the flora of Mt Egmont. Newsletter of the Auckland Botanical Society 31: 1–4.
- Druce AP 1976a. The vegetation. In: Fullarton JH ed. Egmont National Park. 3rd edition. New Plymouth, Egmont National Park Board. Pp. 39–50.
- Druce AP 1976b. Egmont botanical studies. In: Seminar on science in national parks, 1976 proceedings. National Parks Series 6. Wellington, National Parks Authority. Pp. 53–57.
- Efford JT 2012. Treeline vegetation composition, structure and dynamics across the AD 1655 Burrell Lapilli deposit, Mt Taranaki, New Zealand. Unpublished MSc thesis. Hamilton, University of Waikato.
- Eggler WA 1963. Plant life of Paricutin volcano, Mexico, eight years after activity ceased. American Midland Naturalist 69: 38–68. 10.2307/2422843
- Forsyth DM, Hone J, Parkes JP, Reid GH, Stronge D 2003. Feral goat control in Egmont National Park, New Zealand, and the implications for eradication. Wildlife Research 30: 437–450. 10.1071/WR02116
- Giles TM, Newman RM, Lowe DJ, Munro AJ 1999. Impact of tephra fall and environmental change: a 1000 year record from Matakana Island, Bay of Plenty, North Island, New Zealand. In: Firth C, McGuire WJ eds. Volcanoes in the quaternary. Geological society, London, Special Publications 161: 11–26.
- Gomez-Romero M, Lindig-Cisneros R, Galindo-Vallejo S 2006. Effect of tephra depth on vegetation development in areas affected by volcanism. Plant Ecology 183: 207–213. 10.1007/s11258-005-9017-z
- Hogg A, Lowe D, Palmer J, Boswijk G, Ramsey C 2011. Revised calendar date for the Taupo eruption derived by 14C wiggle-matching using a New Zealand kauri 14C calibration data set. The Holocene 22: 1–11.
- Horrocks M, Ogden J 1998. The effects of the Taupo tephra eruption of c. 1718 BP on the vegetation of Mt Hauhungatahi, central North Island, New Zealand. Journal of Biogeography 25: 649–660. 10.1046/j.1365-2699.1998.2540649.x
- Hotes S, Poschlod P, Takahashi H, Grootjans AP, Adema E 2004. Effects of tephra deposition on mire vegetation: a field experiment in Hokkaido, Japan. Journal of Ecology 92: 624–634. 10.1111/j.0022-0477.2004.00901.x
- Husheer SW 2006. Changes to the forest of Egmont National Park 1977–2001. Department of Conservation Research & Development Series 257. Wellington, Department of Conservation.
- Keith RP, Veblen TT, Schoennagel TL, Sherriff RL 2010. Understory vegetation indicates historic fire regimes in ponderosa pine-dominated ecosystems in the Colorado Front Range. Journal of Vegetation Science 21: 488–499. 10.1111/j.1654-1103.2009.01156.x
- Kent M, Owen NW, Dale P, Newnham RM, Giles TM 2001. Studies of vegetation burial: a focus for biogeography and biogeomorphology?Progress in Physical Geography 25: 355–482.
- Lees CM, Neall VE 1993. Vegetation response to volcanic eruptions on Egmont Volcano, New Zealand, during the last 1500 years. Journal of the Royal Society of New Zealand 23: 91–127. 10.1080/03036758.1993.10721220
- Lowe DJ, Newnham RM, McCraw JD 2002. Volcanism and early Maori society in New Zealand. In: Torrence R, Grattan J eds. Natural disasters and cultural change. London, Routledge. Pp. 126–161.
- Lusk CH, Ogden J 1992. Age structure and dynamics of a podocarp-broadleaf forest, Tongariro National Park, New Zealand. Journal of Ecology 80: 379–393. 10.2307/2260684
- McCune B, Grace JB 2002. Analysis of ecological communities. Oregon, MjM Press.
- McCune B, Mefford MJ 2011. PC-ORD: multivariate analysis of ecological data Version 6.0. Oregon, MjM Software.
- McGlone MS, Neall VE, Clarkson BD 1988. The effect of recent volcanic events and climatic changes on the vegetation of Mt Egmont (Mt Taranaki), New Zealand. New Zealand Journal of Botany 26: 123–144. 10.1080/0028825X.1988.10410105
- Nakashizuka T, Iida S, Suzuki W, Tanimoto T 1993. Seed dispersal and vegetation development on a debris avalanche on the Ontake volcano, central Japan. Journal of Vegetation Science 4: 537–542. 10.2307/3236081
- Neall VE 1980. Volcanology. In: Tullett JS ed. Egmont National Park Handbook. 4th edition. New Plymouth, Egmont National Park Board. Pp. 51–58.
- Neall VE, Stewart RB, Smith IEM 1986. History and petrology of the Taranaki volcanoes. In: Smith IEM ed. Late Cenozoic volcanism in New Zealand. Royal Society of New Zealand Bulletin 23: 251–263.
- New Zealand Forest Service 1965. Preliminary report and plan for control of noxious animal, Egmont National Park 1966–1970. Palmerston North, New Zealand Forest Service.
- Nicholls JL 1963. Vulcanicity and indigenous vegetation in the Rotorua district. Proceedings of the New Zealand Ecological Society 10: 58–65.
- Ogden J, Fordham RA, Horrocks M, Pilkington S, Serra RG 2005. Long-term dynamics of the long-lived conifer Libocedrus bidwillii after a volcanic eruption 2000 years ago. Journal of Vegetation Science 16: 321–330.
- Oner M, Oflas S 1997. Plant succession on the Kula volcano in Turkey. Vegetatio 34: 55–62. 10.1007/BF00119886
- Platz T, Cronin SJ, Procter JN, Neall VE, Foley SF 2012. Non-explosive, dome-forming eruptions at Mt Taranaki, New Zealand. Geomorphology 136: 15–30. 10.1016/j.geomorph.2011.06.016
- Rogers G 1997. Trends in health of pahautea and Hall's totara in relation to possum control in central North Island. Science for Conservation 52. Wellington, Department of Conservation.
- Smale MC, Kimberley MO 1993. Regeneration patterns in montane conifer broadleaved forest on Mt Pureora, New Zealand. New Zealand Journal of Forestry Science 23: 123–141.
- Smale MC, Smale PN 2003. Dynamics of upland conifer/broadleaved forest at Waihaha, central North Island, New Zealand. Journal of the Royal Society of New Zealand 33: 509–528. 10.1080/03014223.2003.9517741
- Swanson FJ, Crisafulli CM, Yamaguchi DK 2005. Geological and ecological settings of Mt. St. Helens before May 18, 1980. In: Dale VH, Swanson FJ, Crisafulli CM eds. Ecological responses to the 1980 eruption of Mt St. Helens. New York, Springer. Pp. 13–26.
- Thrippleton T, Dolos K, Perry GLW, Groeneveld J, Reineking B 2014. Simulating long-term vegetation dynamics using a forest landscape models: the post-Taupo succession on Mt Hauhungatahi, North Island, New Zealand. New Zealand Journal of Ecology 38: 26–38.
- Titus JH, Tsuyuzaki S 2003. Distribution of plants in relation to microsites on recent volcanic substrates on Mt Koma, Hokkaido Japan. Ecological Research 18: 91–98. 10.1046/j.1440-1703.2003.00536.x
- Tonkin PJ 1970. The soils of the southeastern sector of Egmont National Park. Earth Science Journal 4: 36–57.
- Topping WW 1972. Burrell Lapilli eruptives, Mt Egmont, New Zealand. New Zealand Journal of Geology and Geophysics 15: 476–490. 10.1080/00288306.1972.10422345
- Tsuyuzaki S 1989. Analysis of revegetation dynamics on the volcano Usu, northern Japan deforested by 1977–1978 eruptions. American Journal of Botany 68: 1468–1477. 10.2307/2444434
- Tsuyuzaki S 1991. Species turnover and diversity during early stages of vegetation recovery on the volcano Usu, northern Japan. Journal of Vegetation Science 2: 301–306. 10.2307/3235920
- Tsuyuzaki S, Hase A 2005. Plant community dynamics on the volcano Mount Koma, Northern Japan, after the 1996 eruption. Folia Geobotanica 40: 319–330. 10.1007/BF02804282
- Veblen TT, Stewart GH 1982. On the conifer regeneration gap in New Zealand: the dynamics of Libocedrus bidwillii stands on South Island. Journal of Ecology 70: 413–436. 10.2307/2259912
- Vucetich CG, Pullar WA. 1963. Ash beds and soils in the Rotorua district. Proceedings of the New Zealand Ecological Society 10: 2–9.
- Whittaker RJ, Walden J, Hill J 1992. Post-1883 ash fall on Panjang and Sertung and its ecological impacts. Geojournal 28: 153–171.
- Wilmshurst JM, McGlone MS 1996. Forest disturbance in the central North Island, New Zealand, following the 1850 BP Taupo eruption. The Holocene 6: 399–411. 10.1177/095968369600600402
- Wilmshurst JM, McGlone MS, Partridge TR 1997. A late Holocene history of natural disturbance in lowland podocarp/hardwood forest, Hawke's Bay, New Zealand. New Zealand Journal of Botany 35: 79–96. 10.1080/0028825X.1997.10410671
- Zobel DB, Antos JA 1997. A decade of recovery of understory vegetation buried by volcanic tephra from Mt St. Helens. Ecological Monographs 67: 317–344. 10.1890/0012-9615(1997)067[0317:ADOROU]2.0.CO;2