
Abstract
Ten new species are described from New Zealand, Cortinarius araniiti, C. artosus, C. canovestitus, C. chlorophyllus, C. cruentoides, C. cuphocyboides, C. cuphomorphus, C. medioscaurus, C. sciurellus and C. wallacei. Two interesting groups of species, designated ‘cuphocyboid’ and ‘scauroid’, are provided with keys.
Introduction
The genus Cuphocybe was erected by Heim (Citation1951) to accommodate two phlegmacioid fungi from New Zealand, distinguished by a missing cortina. Later a few more species were added (Horak Citation1973; Soop Citation1998), but the genus remained confined mainly to the South Pacific. Observing these common fungi in the native Nothofagus forests of New Zealand, one can nevertheless note that, even without a distinct cortina, the universal veil is usually composed of several filamentous layers that rupture simultaneously during the development of the fruit body (pers. obs.). The absence of a cortinal zone causes this veil to settle on the full length of the stipe, rather than on its lower part (cf. ), as is the case for most cortinars. Peintner et al. (Citation2002) showed that the genus nests in Cortinarius (Pers.) Gray, where it is polyphyletic, thus making Cuphocybe a synonym of Cortinarius.
Figure 1 Basidiocarps. A, Cortinarius canovestitus. B, Cortinarius cuphomorphus. C, Cortinarius cuphocyboides. D, Cortinarius wallacei. E, Cortinarius chlorophyllus. F, Cortinarius medioscaurus. G, Cortinarius artosus. H, Cortinarius cruentoides. I, Cortinarius sciurellus. J, Cortinarius araniiti.

Cortinarius section Defibulati was introduced by Moser (Citation1983) to contain a number of species lacking clamp connections in their hyphae, a very rare feature in the genus. About seven species in the northern hemisphere are currently included in the section, where they share most of their characters with sect. Myxacium (Fr.) Gillot & Lucand. Garnica et al. (Citation2005) showed that one sequestrate Cortinarius from Australia nests in the Defibulati clade together with two New Zealand taxa: C. gymnocephalus Soop and an undescribed species, C. aff. anauensis. Section Defibulati was hence confirmed to be monophyletic and bihemispherical.
The fruit bodies of southern Defibulati species evoke Cuphocybe by having a rudimentary or missing cortina, a feature connected with the gelatinous veil, which remains more or less confluent with the cortina in a way homologous to the universal veil of Cuphocybe outlined above. The likeness is strengthened by the ensuing velar pattern on the stipe, and by the fact that southern Defibulati tend to possess less viscid or even dry stipes when compared with their northern sister taxa.
It has long been evident to this author that the native forests of New Zealand host several undescribed species of Cuphocybe habit. This sixth instalment in a series on new Cortinarius taxa from New Zealand deals with this group in part. Other species described in the present study may be placed in subgenera Dermocybe (Fr.) Trog and Phlegmacium (Fr.) Trog, as morphologically defined by Brandrud et al. (Citation1989–1997). Species in these subgenera were partly considered in the previous instalments (Soop Citation2001, Citation2002, Citation2005, Citation2010, Citation2013), and by Horak & Taylor (Citation1981); Horak (Citation1987), Soop (Citation1998) and Gasparini & Soop (Citation2008), but have not been otherwise treated in publications aimed at the mycota of the country. The infrageneric geotaxonomy of Cortinarius, and in particular of the studied groups, was extensively discussed in Soop & Gasparini (Citation2011).
All taxa described in the present study are assumed to form obligatory mycorrhiza with native trees (Nothofagus and Leptospermum/Kunzea spp.). One would therefore seek to identify them primarily in descriptions from the corresponding habitats in Australia and Patagonia (South America), these being the only regions where analogous studies have been undertaken (among these, notably Cleland & Harris Citation1948; Moser & Horak Citation1975; Bougher & Hilton Citation1989; Gasparini Citation2001a,Citationb, Citation2007; Garnica et al. Citation2002).
Methods
In the descriptions, measurements of the fruit body pertain to adult specimens; these are the diameter of the pileus, the length of the stipe, and the diameter of the upper part of the stipe. Lamella colour pertains to immature individuals unless specified otherwise, the designation ‘L =’ means the number of lamellae reaching the stipe, and ‘l =’ indicates the number of lamellulae between two lamellae. The universal and partial veils of the fruit body are referred to as veil and cortina, respectively.
The alkaline reaction was made with a 30% NaOH solution. Fluorescence is reported under Macrochemical reactions; the test consists of irradiating the context of a fresh fruit body with ultraviolet (UV) light in an otherwise dark room. Microscopic observations were made principally on material mounted in 5% ammonia solution, with or without Congo red, and examined with an oil-immersion objective (1000 ×). The term ‘marginal elements’ refers to sterile cells that are found on the lamellar edge. The spore measurements are expressed as the mean value with one-sigma limits and, in parentheses, the observed maxima and minima. The Q value is the length/width quotient; its average and standard deviation are reported. The n value is the number of measurements.
Species mentioned for comparison under ‘Comments’ were described from New Zealand unless otherwise specified. All holotype material has been deposited in the PDD herbarium, Auckland, New Zealand. Some of the material has been sequenced by Landcare Research, Auckland. The GenBank accession numbers and herbarium numbers are given under ‘Type species’ and ‘Collections examined’. Unless stated otherwise, collections were made by the author and the descriptor ‘KS’ indicates the author's collection number. Collection localities are referred to the districts defined by Crosby et al. (Citation1998).
Taxonomy
Key to the cuphocyboid taxa of New Zealand
The group is characterised by a missing or rudimentary cortina, which causes the often abundant veil to settle as tufts and squamules on the full length of the stipe. Fruit bodies are medium-sized to large, and the stipe is cylindrical, often with a small rounded or piston-like bulb (cf. Horak Citation1973).
1 Pileus dry ........................................................................................................................................2
1* Pileus viscid to glutinous...........................3
2 Pileus pink to pale buff with pinkish squamules, with Nothofagus..............................................................................................................................Cortinarius alboroseus (R. Heim) Peintner et al.
2* Pileus greyish with grey to grey-brown hairs and fibres, with myrtaceous hosts............................................................................................................................................1. C. canovestitus sp. nov.
3(1) Odour in context strong, sweetish, like honey........................................................................................................................................C. dulciolens E. Horak et al. (≡ Cuphocybe melliolens Soop)
3* Odour strong like moth-balls........................................................................C. naphthalinus Soop
3** Odour different or faint...................................................................................................................4
4 Pileus golden fulvous................................................................... C. elaiochrous var. leontis Soop
4* Pileus dark brown to olive-brown, clamp connections present .............................................5
4** Pileus grey to greyish brown, clamp connections absent (sect. Defibulati) ..........................................6
5 Stipe with a greenish to citrinous sheen, fruit body often robust.......................................................................................................C. elaiochrous E. Horak et al. (≡ Cuphocybe olivacea R. Heim)
5* Stipe yellowish grey with ochraceous squamules, fruit body smaller.......................................................................................................................................C. phaeomyxa (E. Horak) Peintner et al.
6(4) Lamellae saturated violet, veil white ........................................................ C. gymnocephalus Soop
6* Lamellae grey-blue, veil greyish to ochraceous .......................................................................................7
7 Pileus glabrous, stipe viscid ...............................................................3. C. cuphocyboides sp. nov.
7* Pileus fibrillose-squamulose, stipe dry...............................................2. C. cuphomorphus sp. nov.
1. Cortinarius canovestitus Soop, sp. nov.
(, ), MycoBank 807549.
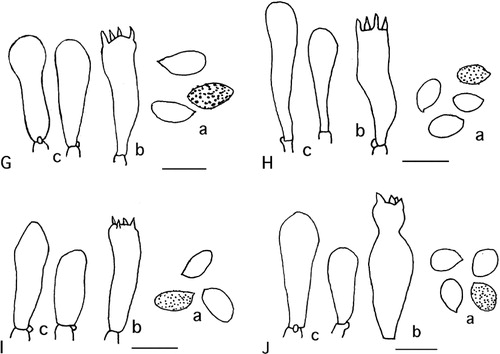
Figure 2 Microscopic details: a: spores, b: basidia, c: sterile marginal elements, scale = 10 µm. A, Cortinarius canovestitus. B, Cortinarius cuphomorphus. C, Cortinarius cuphocyboides. D, Cortinarius wallacei. E, Cortinarius chlorophyllus. F, Cortinarius medioscaurus. G, Cortinarius artosus. H, Cortinarius cruentoides. I, Cortinarius sciurellus. J, Cortinarius araniiti.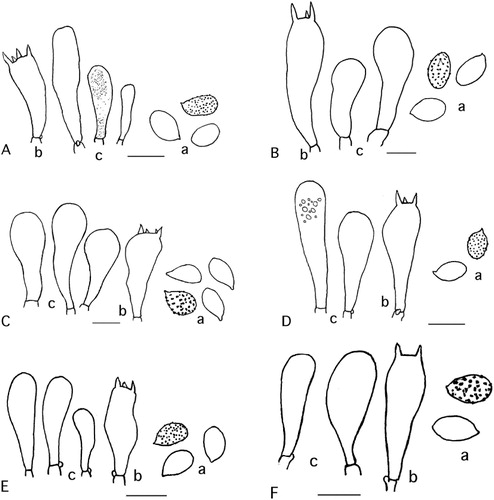
Pileo 25–50 mm diam., hemispherico, deinde convexo vel expanso, sicco, nec hygrophano, griseo-brunneo vel fulvo-pallido, fibrilllis cano-flavis raditatis vestito. Lamellis primo pallide cinnamomeis, subdistantibus. Stipite aequali vel subclavato, albo-griseo, copiose cano-cingulato. Velo pallido cano-flavo, subcopioso, cortina exigua vel abest. Carne grisea, fragile, odore debile raphanoide, sapore nullo. Sporis ellipsoideis 9–10.6 × 5.6–6.4 µm, moderate verrucosis. Reactionem ope NaOH nullam. In silvis cum Leptospermate, raro.
Typus. New Zealand. Buller, St Arnaud Campsite, in a mixed Leptospermum scoparium and Nothofagus forest, 4 May 2013, K. Soop KS-CO2095, holotype PDD 103667, isotype S F244778, GenBank KF727353 (ITS).
Description. Pileus 25–50 mm diameter, hemispherical, later convex to expanded, dry, hygrophanous, grey-brown to pale tan, irregularly covered by greyish yellow radial fibrils, sometimes with tufts and patches, elsewhere glabrous to finely innate fibrillose, margin not striate. Lamellae pale cinnamon when young, free, rather distant (L = 34–44, l = 1–2), edge concolorous. Stipe 35–75 × 5–9 mm, dry, cylindrical to slightly clavate, greyish-white with thick greyish girdles and tufts, apex pale yellow. Veil pale yellow to greyish yellow, fairly copious, cortina rudimentary or absent. Context greyish, marbled yellow in stipe-base, often fragile, exsiccata pale tan. Macrochemical reactions: NaOH trivial. Odour faint, raphanoid, taste none.
Spores (8.7−)9.1−9.9−10.6(−11.1) × (5.1−)5.6−6−6.4(−6.8) µm, Q = 1.65 ± 0.13 (n = 26), elliptic, moderately verrucose. Marginal elements crowded, clavate to vesiculose, 20–35 × 8–9 µm. Basidia 25–35 × 6–8 µm, four-spored. Pileipellis of repent hyphae 3–5 µm wide. Hypocutis of oblong elements, 20–40 × 9–16 µm. Clamp connections present.
Habitat. Gregarious, rare, associated with Leptospermum scoparium.
Other collections examined. New Zealand. Buller, Boyle River, 11 April 1997, KS-CO790 (PDD 103633), GenBank KF727351 (ITS); Buller, St Arnaud Campsite, 5 May 2008, KS-CO1790 (PDD 103644), GenBank KF727352 (ITS).
Etymology. From Latin cani ‘grey hair’ and vestis ‘clothing’, due to the general aspect.
Comments. This species is rather similar to a Cortinarius alboroseus (R. Heim) Peintner et al. without the pink coloration. Moreover, the latter is found in Nothofagus forests, whereas C. canovestitus is associated with myrtaceous hosts. The greyish veil remnants on the pileus often form radially oriented hairs, reminiscent of a balding head, and the fruit body is typically airy and brittle. Available molecular data (see above) indicate that C. canovestitus is closely related to C. subcastanellus E. Horak et al., a species of Rozites habit.
2. Cortinarius cuphomorphus Soop, sp. nov.
(, ), MycoBank 807550.
Pileo 35–55 mm diam., hemispherico, deinde convexo vel plano, viscido, parum vel haud hygrophano, pallide griseo-brunneo, dense fibrilloso-squamuloso, demum ad discum glabro, pallescente. Lamellis primo griseo-violaceis, subconfertis. Stipite sicco, aequali, albo, toto griseo-luteo squamuloso. Velo pallide luteo, subcopioso; cortina exigua vel abest. Carne alba vel subviolacea, griseo-violaceo marmorata; odore dulci, sapore nullo. Sporis ellipsoideis 11.4–12.6 × 7.4–8 µm, subgrosse verrucosis. Reactionem ope NaOH nullam. In silvis cum Nothofagete, raro.
Typus. New Zealand. Taupo, Clements Mill Rd, in Nothofagus forest, 10 May 2013, K. Soop KS-CO2108, holotype PDD 103680, isotype S F244807, GenBank KF727387 (ITS).
Description. Pileus 35–55 mm diameter, hemispherical, later convex to plane, viscid, weakly or not hygrophanous, pale grey-brown with dense fibrils and squamules, pallescent on disc which turns glabrous with age, margin not striate. Lamellae grey-blue when young, free, moderately crowded (L = 60, l = 2), edge concolorous. Stipe 30–70 × 6–13 mm, dry, cylindrical, sometimes with a small, piston-like bulb, white, with grey to yellowish squamules over the whole length, apex concolorous. Veil pale yellow to ochraceous grey, fairly copious, cortina rudimentary or absent. Context white to pale violet, marbled grey-blue, exsiccata pale brownish. Macrochemical reactions: NaOH weakly red-brown on cutis, elsewhere none. Odour sweetish, sometimes strongly, taste none.
Spores (10.9−)11.4−12−12.6(−13.1) × (7.1−)7.4−7.7−7.9(−8.1) µm, Q = 1.56 ± 0.09 (n = 26), elliptic, rather coarsely verrucose. Marginal elements crowded, vesiculose, 25–45 × 8–14 µm. Basidia 30–45 × 13 µm, four-spored. Pileipellis with an outer layer of sinuous gelified hyphae 3–5 µm wide, inner layer of repent hyphae 4–6 µm wide, many with a yellow pigmentation. Hypocutis of hyaline oblong elements, 40–70 × 14–20 µm. Clamp connections absent.
Habitat. Gregarious, rare, associated with Nothofagus spp.
Other collections examined. New Zealand. Taupo, Te Iringa Track, 10 April 2013, KS-CO2105 (PDD 103677), GenBank KF727386 (ITS); idem 11 April 2013, KS-CO2099 (PDD 103671), GenBank KF727385 (ITS).
Etymology. From Cuphocybe and Greek µoρϕη ‘shape’, due to the general aspect.
Comments. A largely greyish fungus, resembling Cortinarius canovestitus (above), but without the hairy aspect of the pileus and presenting a violet tint on lamellae and context. Cortinarius cuphomorphus is further characterised by the absence of clamp connections. Available molecular data (see above) indicate that the species belongs to the Defibulati clade, where it is the only known member of the section with a dry stipe. It is so far recorded only from the Kaimanawa mountains.
3. Cortinarius cuphocyboides Soop, sp. nov.
(, ), MycoBank 807551.
Pileo 35–70 mm diam., hemispherico, deinde convexo, glutinoso, parum hygrophano, griseo, ad discum subflavescente, glabro. Lamellis primo griseis, subviolaceis, subconfertis. Stipite viscido, aequali, albo-pruinato. Velo albo-griseo, copioso, cortina exigua vel abest. Carne griseo-alba, leviter violaceo-marmorata, odore saporeque subnullibus. Sporis amygdaloideis vel citriformis 12–13.6 × 7.3–8 µm, grosse verrucosis. Reactionem ope NaOH nullam. In silvis cum Nothofagete, valde raro.
Typus. New Zealand. Otago Lakes, Te Anau Downs, in Nothofagus forest, 16 April 1999, K. Soop KS-CO1018, holotype PDD 88293, GenBank AY669625 (ITS).
Description. Pileus 35–70 mm diameter, hemispherical, later convex, glutinous, weakly hygrophanous, cutis thick, gelatinous, tenacious, grey with a slightly flavescent disc, darker brown when old, glabrous. Lamellae grey with a faint violet tinge when young, free, moderately crowded (L = 64), edge concolorous. Stipe 55–70 × 12–14 mm, viscid, cylindrical, white frosty and thinly zoned, fulvescent with age, apex concolorous. Veil greyish white, copious, cortina rudimentary or absent. Context greyish white, slightly marbled violet, exsiccata red-brown to yellow-brown. Macrochemical reactions: NaOH and tincture of gum guaiac none. Odour none, taste none to slightly bitter.
Spores (11.7−)12−12.8−13.6(−14.7) × (7.1−)7.3−7.7−8.1(−8.7) µm, Q = 1.66 ± 0.15 (n = 26), amygdaloid to citriform, often with one papilla, rather coarsely to coarsely verrucose. Marginal elements crowded, clavate to vesiculose, many yellow encrusted, 30–40 × 10–15 µm. Basidia 30–38 × 12 µm, four-spored. Pileipellis with an outer thick layer of sinuous gelified hyphae 1.5–2.5 µm wide, inner layer of repent hyphae 4–5 µm wide. Hypocutis of irregular elements, 30–50 × 12–20 µm, many with a yellow pigmentation. Clamp connections absent.
Habitat. Gregarious, very rare, associated with Nothofagus spp.
Etymology. From Cuphocybe and Greek ειδoς ‘similar’, due to the general aspect.
Comments. Like Cortinarius cuphomorphus (above), this fungus is greyish and devoid of clamp connections, differing mainly by a glabrous pileus and a more viscid sporocarp. In addition, many spores exhibit an unusual shape, being papillate at one end only. Cortinarius cuphocyboides was included in a major study of the phylogenetic classification in Cortinarius by Garnica et al. Citation2005 (as ‘C. aff. anauensis’), where it was an exponent of southern Defibulati. The species has not been recorded again in New Zealand, but it was deemed worth publishing it in this study in view of its importance in the bihemispherical phylogeny of the genus.
4. Cortinarius wallacei Soop, sp. nov.
(, ), MycoBank 807552.
Pileo 25–55 mm diam., obtuse hemispherico, deinde convexo vel expanso, glutinoso, parum vel haud hygrophano, fervide rubro-rufo vel fulvo, glabro vel minute innato-fibrilloso. Lamellis primo pallide cinnamomeis, subconfertis. Stipite sicco, aequali, albo vel pallide griseo-brunneo, luteo-griseo annulato. Velo rufo, sparso. Carne pallide fulva, odore saporeque subnullibus. Sporis subobtuse ellipsoideis 8–9 × 5.2–6 µm, moderate verrucosis, forte dextrinoideis. Reactionem ope NaOH rubro-rufam. In silvis cum Nothofagete, subvulgare.
Typus. New Zealand. Taupo, Cascade Hut Track, in Nothofagus forest, 5 May 2009, K. Soop KS-CO1918, holotype PDD 97543, isotype S F140324, GenBank KJ635242 (ITS+LSU).
Description. Pileus 25–55 mm diameter, obtusely rounded, later convex to expanded, glutinous, weakly or not hygrophanous, warmly red-brown to yellow-brown, glabrous to finely innate-fibrillose, margin devoid of velar remnants, weakly striate, sometimes sulcate. Lamellae pale cinnamon when young, soon with an orange tinge, broadly emarginate, moderately crowded (L = 42–48, l = 2–3), edge concolorous. Stipe 30–70 × 5–8 mm, dry, cylindrical, often with a small, piston-like bulb, white to pale grey-brown with sparse red-brown fibrils and bands below the annulus, which is membranous, greyish yellow, striate, and placed high. Veil red-brown, sparse. Context pale tan. Macrochemical reactions: NaOH red-brown to red on cutis and context, tincture of gum guaiac strongly blue-green. Odour nil or faint, somewhat like wax candles, taste none.
Spores (7.6−)8−8.5−9(−9.3) × (4.6−)5.2−5.6−5.9(−6) µm, Q = 1.54 ± 0.12 (n = 28), rather obtusely elliptic, moderately verrucose, strongly dextrinoid. Marginal elements crowded, clavate to vesiculose, some with a hyaline granulose matter, 17–35 × 6–9 µm. Basidia 25–30 × 8–9 µm, four-spored. Pileipellis with an outer layer of sinuous gelified hyphae 2–4 µm wide, inner layer thin of ± repent hyphae with a brown-yellow pigment, most brown incrusted, 3–5 µm wide. Hypocutis of rectangular to irregular elements, 35–45 × 12–25 µm, with a pale-yellow pigmentation. Clamp connections present.
Habitat. Gregarious, fairly common, associated with Nothofagus spp.
Other collections examined. New Zealand. Taupo, Te Iringa Track, 13 May 2001, KS-CO1256 (PDD 73136), GenBank AY669623 (ITS); idem 8 May 2009, KS-CO1913 (PDD 97538, S F140320); idem 13 May 2013, KS-CO2043 (PDD103889, S F190009), GenBank KF727358 (ITS); Otago Lakes, Haast Pass, Cameron Creek, 2 May 2009, KS-CO1903 (PDD 97528, S F140312); Buller, Lake Daniel Track, 4 May 2011, KS-CO2029 (PDD101856, S F190001).
Etymology. In honour of Mike Wallace, a valued colleague who participated in the discovery of the species.
Comments. This fungus of Rozites habit recalls Cortinarius subcastanellus E. Horak et al., with which it has often been confused. In contrast to the latter, C. wallacei presents a handsome, mahogany-brown pileus that is almost devoid of veil remnants. It also differs by the placement of the stipital collar, the alkaline reaction, and the dextrinoid spores. Cortinarius gamundiae (E. Horak) E. Horak et al., described from Nothofagus habitat in Patagonia, differs by more oblong spores (up to 12.5 µm long) and yellow lamellae (see Horak Citation1979 sub Rozites gamundiae). Two other species are similar to C. wallacei: C. rugosiceps (E. Horak & G.M. Taylor) Peintner et al. and C. armeniacovelatus (Bougher et al.) Peintner et al., described respectively from New Zealand and Australia. They both produce significantly larger spores and also deviate in several smaller ways. As can be inferred from available molecular data (see above), C. wallacei forms a sister clade to C. subcastanellus. The latter name has also been used for several collections of C. wallacei in GenBank (GU222270, AY033112, AY669623; cf. collection CO1256 in Peintner et al. Citation2004; Garnica et al. Citation2005).
Key to the scauroid taxa of New Zealand
This phlegmacicoid group is characterised by a marginate bulbous stipe and violaceous tints at least in the context. Compared with the European mycota, it has relatively few representatives in the country, partly because the populous section Calochroi M.M. Moser & E. Horak appears to be missing in the South Pacific (cf. Garnica et al. Citation2005).
1 Lamellae violaceous.........................................................................................................................2
1* Lamellae dark green............................................................. 5. Cortinarius chlorophyllus sp. nov.
2 Pileus light tan, stipe cylindrical with a small bulb..................................... 7. C. artosus sp. nov.
2* Pileus darker, stipe bulb prominent, marginate.............................................................................. 3
3 Associated with myrtaceous trees ..................................................................................................4
3* Associated with Nothofagus ...........................................................................................................5
4 Fruit body small, pileus often < 30 mm, taste mild..................................... C. minoscaurus Soop
4* Fruit body usually larger, taste bitter .................................................6. C. medioscaurus sp. nov.
5(3) Pileus with greenish or olive tints .......................................................................C. turcopes Soop
5* Pileus mahogany brown to date brown.......................................................................................... 6
6 Pileus hygrophanous, reaction positive with iodine solutions .........................C. singularis Soop
6* Pileus not hygrophanous, iodine reaction absent ......................................C. fiordlandensis Soop
5. Cortinarius chlorophyllus Soop, sp. nov.
(, ), MycoBank 807553.
Pileo 20–45 mm diam., obtuse hemispherico, deinde expanso, viscido, haud hygrophano, fusco-fulvo vel umbrino-brunneo et olivaceo-tincto, innato-fibrilloso. Lamellis primo fusco-viridis vel olivaceo-viridis, confertis. Stipite sicco, marginato-bulboso, violaceo. Velo olivaceo-flavo, subsparso, cortina pallide luteo-viride. Carne griseo-violacea, odore submelleo, sapore debili. Sporis amygdaloideis vel subellipsoideis, 7.7–8.5 × 5–5.7 µm, moderate verrucosis. Reactionem ope NaOH et Lugol sanguinem. In silvis cum Nothofagete, raro.
Typus. New Zealand. Taupo, Te Iringa Track, in Nothofagus forest, 11 May 2013, KS-CO2109, holotype PDD 103681, isotype S F244810, GenBank KF727375 (ITS).
Description. Pileus 20–45 mm diameter, obtusely rounded, later expanded, viscid, not hygrophanous, dark yellow-brown to umber with an olive tinge, innate-fibrillose. Lamellae dark greyish green to olive green when young, narrowly emarginate, moderately crowded (L = 56. l = 1–2), edge paler. Stipe 35–45 × 9–12 mm, dry, with a wide, marginate bulb, violaceous, greying towards base with a yellowish coating on bulb margin. Veil ochraceous yellow, rather sparse, cortina pale yellow-green. Context grey-violet. Macrochemical reactions: NaOH blood red on stipital veil, Lugol blood red in context. Odour none or faint, like honey, taste faint, pleasant.
Spores (7.3−)7.7−8.1−8.5(−9.2) × (4.9−)5.1−5.4−5.7(−6) µm, Q = 1.50 ± 0.09 (n = 28), amygdaloid to subelliptic, moderately verrucose. Marginal elements crowded, clavate to cylindrical, 12–30 × 7–10 µm. Basidia 22–28 × 8–10 µm, four-spored. Pileipellis with an outer layer of sinuous gelified hyphae 2–4 µm wide, inner layer of ± repent and tangled hyphae with a brown-yellow pigment, 4–5 µm wide. Hypocutis of rectangular to irregular elements, 25–45 × 12–20 µm. Clamp connections present.
Habitat. Solitary, rare, associated with Nothofagus spp.
Etymology. From Greek χλωρoς ‘green’ and φυλλoν ‘leaf’, with reference to the colour of the lamellae.
Other collections examined. New Zealand. Fiordland, Lake Gunn Track, 24 April 2004, KS-CO1419 (PDD 103640), GenBank KF727374 (ITS).
Comments. This species is a typical exponent of section Scauri (Fr.) Henn., characterised by a marginate bulbous stipe, violaceous tints in the context, and a positive reaction with iodine solutions. It differs from Cortinarius singularis Soop, common in the same habitat, by handsomely dark green lamellae and smaller and markedly less verrucose spores. Available molecular data (see above) place C. chlorophyllus in the Scauri clade, where it differs from C. singularis by only six substitutions in the internally transcribed spacer region, a not uncommon inter-specific difference in phlegmacioid clades (T.E. Brandrud, pers. comm.). These two species are the only members of the section so far reported from New Zealand.
6. Cortinarius medioscaurus Soop, sp. nov.
(, ), MycoBank 807554.
Pileo 25–50 mm diam., obtuse hemispherico, deinde expanso, viscido, parum vel haud hygrophano, fulvo, ad discum rufo-tincto, innato-fibrilloso, sæpe velo pallide luteo-maculato. Lamellis primo griseo-violaceis, subconfertis. Stipite sicco, marginato-bulboso, violaceo at fulvescente. Velo pallide luteo, subcopioso. Carne alba griseo-violaceo marmorata, odore nullo, sapore perspicue amaro. Sporis amygdaloideis vel subellipsoideis, 10–11.5 × 5.5–6.5 µm, subgrosse verrucosis. Reactionem ope NaOH nullam. In silvis cum Leptospermate, subvulgare.
Typus. New Zealand. Auckland, Albany, Horton Reserve, in Kunzea copse, 17 May 2013, KS-CO2121, holotype PDD 103691, isotype S F244821, GenBank KF727397 (ITS).
Description. Pileus 25–50 mm diameter, obtusely rounded, later expanded, when mature somewhat upturned, viscid, weakly or not hygrophanous, warmly yellow-brown with a red-brown tinge on disc, innate-fibrillose, often with pale yellow veil patches. Lamellae greyish violet when young, fairly crowded. Stipe 25–55 × 6–13 mm, dry, with a marginate bulb, violaceous, turning yellow-brown with age, young with a yellowish coating on bulb margin, apex violet. Veil pale yellow to grey-yellow, fairly copious, cortina not noted. Context white, marbled violet, exsiccata yellow-brown, later red-brown. Macrochemical reactions: None with NaOH, phenol, Lugol. Odour none, taste distinctly bitter.
Spores (9.3−)10−10.7−11.5(−12) × (5.1−)5.5−5.9−6.4(−6.8) µm, Q = 1.82 ± 0.17 (n = 28), amygdaloid to subelliptic, rather coarsely verrucose. Marginal elements fairly crowded, clavate, 22–28 × 8–10 µm. Basidia 22–28 × 7–10 µm, four-spored, some two-spored or one-spored. Pileipellis with an outer layer of sinuous gelified hyphae 2–4 µm wide, inner layer of tangled hyphae with a brown-yellow pigment, 3–4 µm wide. Hypocutis of irregular elements, 35–55 × 20–25 µm. Clamp connections present.
Habitat. Gregarious, fairly common, associated with myrtaceous trees.
Etymology. From Latin medius ‘intermediary’ and Scauri, from its intermediate size among scauroid taxa.
Other collections examined. New Zealand. Auckland, Hunua Mountains, Mangatangi Track, 25 April 2001, KS-CO1174 (PDD 73138), GenBank KJ635224 (ITS+LSU); idem 25 April 2001, KS-CO1175 (S F46632).
Comments. Another species with scauroid habit, but not closely related to section Scauri (cf. the preceding Comments) as evidenced by its negative Lugol reaction. Cortinarius medioscaurus is almost identical to C. minoscaurus Soop, very rare in the same habitat. Their only differential morphological characters are the somewhat smaller fruit bodies of the latter, its mild taste, and its more grossly ornamented spores. As can be seen from available molecular data (see above); however, they are genetically well separated, Cortinarius medioscaurus occupying a basal position to the former, as well as to most other clades examined, including Scauri.
7. Cortinarius artosus Soop, sp. nov.
(, ), MycoBank 807555.
Pileo 55–70 mm diam., obtuse hemispherico, deinde expanso vel subplano, viscido, haud hygrophano, æque fulvo-pallido, innato-fibrilloso. Lamellis primo griseo-violaceis, confertis. Stipite sicco, bulboso, albo, apice violaceo, ad bulbam luteo-vestito. Velo luteo, sparso. Carne alba, violaceo-marmorata, odore saporeque nullis. Sporis amygdaloideis, 10–11 × 5.3–6.3 µm, subgrosse verrucosis. Reactionem ope NaOH nullam. In silvis cum Nothofagete, valde raro.
Typus. New Zealand. Auckland, Hunua Mountains, Workman Track, in Nothofagus forest, 8 May 2003, KS-CO1348, holotype PDD 77486, GenBank AY669644 (ITS).
Description. Pileus 55–70 mm diameter, obtusely rounded, expanded to almost plane, viscid, not hygrophanous, evenly pale tan, finely innate-fibrillose. Lamellae greyish violet when young, adnate, crowded. Stipe 50–90 × 11–14 mm, dry, cylindrical with a small bulb, white with a yellowish coating on bulb, apex with a violet tinge. Veil butter yellow, sparse, cortina not noted. Context white, marbled violet, exsiccata pale. Macrochemical reactions: None with NaOH and tincture of gum guaiac. Odour and taste none.
Spores (9.5−)9.9−10.4−11(−11.5) × (4.4−)5.3−5.8−6.3(−6.8) µm, Q = 1.81 ± 0.20 (n = 27), amygdaloid, rather coarsely verrucose. Marginal elements fairly crowded, clavate to capitate, 20–25 × 8–10 µm. Basidia 25–30 × 7–8 µm, four-spored. Pileipellis with an outer layer of sinuous gelified hyphae 2–3 µm wide, inner thick layer of repent hyphae 3–5 µm wide, many with a yellow pigmentation. Hypocutis of hyaline oblong to fusoid elements, 25–60 × 6–10 µm. Clamp connections present.
Habitat. Gregarious, very rare, associated with Nothofagus spp.
Etymology. From Greek αρτoς ‘bread’, with reference to the crust-like pileus colour.
Comments. A rather inconspicuous phlegmacioid fungus with a pale brown-yellow pileus and a violaceous tinge on the stipe. Cortinarius artosus was included in a major study of the phylogenetic classification in Cortinarius by Garnica et al. (2005; as ‘C. sp.’), where it was shown to be unique to the New Zealand mycoflora by nesting in the boreal section Percomes (Konrad & Maubl.) Melot. (The only other known southern Percomes is the Australian C. coelopus Gasparini) The species has not been recorded again in New Zealand, but it was deemed worth publishing it in this study in view of its importance in the bihemispherical phylogeny of the genus.
8. Cortinarius cruentoides Soop, sp. nov.
(, ), MycoBank 807556.
Pileo 10–30 mm diam., late conico, deinde subplano, sicco, parum hygrophano, fusco-cramesino, deinde pallescente et brunnescente, minute squamuloso vel granuloso, margine leviter rubrofibrilloso et striato. Lamellis primo intense cramesinis, distantibus. Stipite æquali, rubro, leviter rubrofibrilloso. Velo fusco-rubro vel pupurato, subcopioso. Carne griseo-alba, rubro-marmorata, odore saporeque nullis. Sporis ellipsoideis, 5.5–7.5 × 4–5.2 µm, moderate verrucosis. Reactionem ope NaOH ubique sanguinem præbet. In silvis cum Leptospermate, nec vulgare.
Typus. New Zealand. Auckland, Huia, Donald Mclean Rd, in Kunzea forest, 10 May 2011, KS-CO2038, holotype PDD 101864, isotype S F190005, GenBank KJ635217 (ITS+LSU).
Description. Pileus 10–30 mm diameter, broadly conical, later almost plane, dry, weakly hygrophanous, dark carmine red, later fading with a brownish tinge, minutely squamulose to granulose, later innate fibrillose, margin often paler with sparse red fibrils, striate. Lamellae intensely carmine red when young, edge often more saturated, free, distant (L = 28, l = 2–3). Stipe 20–40 × 2–4 mm, cylindrical, slender, red to pale red, with red tufts and squamules, apex pale red. Veil dark red to purple red, rather copious, cortina not noted. Context grey-white with a pink tinge, marbled red. Macrochemical reactions: NaOH blood red on cutis, stipital veil and lamellae, fluorescence in UV-light none. Odour and taste none.
Spores (6−)6.5−6.9−7.3(−7.6) × (3.8−)4.2−4.5−4.9(−5.4) µm, Q = 1.81 ± 0.20 (n = 27), elliptic, moderately verrucose. Marginal elements crowded, clavate, 15–30 × 7–9 µm, some with pink contents. Basidia 20–25 × 7–8 µm, four-spored. Pileipellis a rather thick layer of sinous and repent elements with a pale red pigmentation, 25–60 × 6–10 µm. Hypocutis of hyaline, rectangular elements, 17–40 × 12–14 µm. Clamp connections present.
Habitat. Gregarious, uncommon, associated with myrtaceous trees.
Etymology. From C. cruentus (Latin ‘bleeding’), a boreal Dermocybe, and Greek ειδoς ‘similar’, with reference to the deep-red overall hue.
Other collections examined. New Zealand. Auckland, Albany, Northwood Reserve, 17 May 2013, KS-CO2122 (PDD 103692), GenBank KF727363 (ITS).
Comments. A small deep red fungus found in Kunzea woods, resembling northern taxa in subgenus Dermocybe, section Sanguinei. Cortinarius cruentoides also resembles C. vinicolor (E. Horak) G. Garnier and C. rubripurpuratus Soop (= Dermocybe purpurata E. Horak & Gerw. Keller), growing in Nothofagus forests. These are even smaller in size and produce leaner and coarsely ornamented spores. Available molecular data (cf. above) indicate that C. cruentoides belongs to section Icterinula M.M. Moser & E. Horak, while several other small, red dermocyboid fungi in the South Pacific, such as Dermocybe kula Grgur., nest in sect. Splendidi Gasparini (Garnica et al. Citation2005; Stefani et al. Citation2013). The species apparently also grows in Tasmania (B. Gasparini, pers. comm.).
9. Cortinarius sciurellus Soop, sp. nov.
(, ), MycoBank 807557.
Pileo 12–35 mm diam., conico, deinde expanso angusto-umbonato, sicco vel leviter viscido, hygrophano, fulvo, ad umbum obscurior, minute innato-fibrilloso, margine interdum subolivaceo, striato vel sulcato. Lamellis primo olivaceo-griseis vel olivaceo-luteis, distantibus. Stipite æquali, luteo subcitrino, fuscescente. Velo ex citrino subfulvo, sparso. Carne luteo-olivaea vel luteo-viride, odore debile raphanoide, sapore interdum subacerbo. Sporis ellipsoideis, 7.5–8.5 × 4.7–5.3 µm, moderate vel subminute verrucosis. Reactionem ope NaOH cute veloque rubram. In silvis cum Nothofagete, subvulgare.
Typus. New Zealand. Buller, Reefton, Murray Creek Track, in Nothofagus forest, 8 May 2006, KS-CO1679, holotype PDD 103641, GenBank KF727369 (ITS).
Description. Pileus 12–35 mm diameter, broadly conical, later expanded with a narrow umbo, dry to slightly viscid, hygrophanous, yellow-brown with a darker umbo, finely innate-fibrillose, margin paler without obvious veil remnants, sometimes with an olive or citrinous tinge when young, striate to sulcate. Lamellae olive-grey to olive-yellow when young, distant (L = 30, l = 2), free. Stipe 40–65 × 2–4 mm, dry, cylindrical, slender, citrinous to yellow, darkening with age, with sparse greyish green fibrils. Veil citrinous to yellow-brown, sparse, cortina pale yellow. Context olive-yellow to greyish yellow. Macrochemical reactions: NaOH red on cutis and stipital veil, orange-red on lamellae, fluorescence in UV-light none. Odour weak, raphanoid, taste faint like ‘raw peas’, sometimes ± acerbic.
Spores (7.1−)7.4−8−8.5(−9) × (4.4−)4.7−5−5.3(−5.5) µm, Q = 1.60 ± 0.10 (n = 29), elliptic, moderately to rather weakly verrucose. Marginal elements scattered, clavate, 20−24 × 6–8 µm. Basidia 24–26 × 6–7 µm, four-spored. Pileipellis a thin layer of repent elements with a brownish yellow pigmentation, c.4 µm wide. Hypocutis a tangled layer of oblong elements with the same coloration, 35–70 × 8–12 µm. Clamp connections present.
Habitat. Gregarious, uncommon, associated with Nothofagus spp.
Etymology. From Latin sciurus ‘squirrel’, due to the general coloration.
Other collections examined. New Zealand. Nelson, Flora Saddle, 10 May 2004, KS-CO1465 (PDD 78800); Otago Lakes, Haast Pass, Bridle Track, 4 May 2006, KS-CO1665 (PDD 88280); idem 22 April 2011, KS-CO2002 (PDD 101832), GenBank KJ635212 (ITS+LSU); idem Cameron Creek, KS-CO2001 (PDD 101831).
Comments. A small yellow-brown fungus, easily mistaken in the field for a Telamonia, whereas the alkaline reaction indicates section Icterinula, a position corroborated by available molecular data. Cortinarius sciurellus resembles C. indotatus (E. Horak) G. Garnier in the same section, which, however, produces distinctly smaller spores. It has sometimes been mistaken for the latter species; for example GenBank GU222322, which in fact represents C. sciurellus. The present taxon also resembles C. olivaceoniger (E. Horak) Garnier, which is a larger species with a viscid stipe.
10. Cortinarius araniiti Soop, sp. nov.
(, ), MycoBank 807548.
Pileo 12–30 mm diam., conico, deinde late conico vel expanso, sicco, hygrophano, aurantio, ad discum aurantio-brunneo tincto, minutissimo innato-fibrilloso, margine substriato. Lamellis primo intense aurantiis, distantibus. Stipite æquali, griseo-luteo vel aurantio-luteo, ad basem leviter aurantio-fibrilloso. Velo aurantio-brunneo, subcopioso. Carne luteo-aurantia vel aurantio-brunnea, odore nullo, sapore subraphanoide. Sporis ellipsoideis, 7–8 × 5.7–6.4 µm, moderate verrucosis. Reactionem ope NaOH ubique sanguinem præbet. In silvis cum Nothofagete, valde raro.
Typus. New Zealand. Mid Canterbury, Craigieburn Res., in Nothofagus forest, 5 May 2009, KS-CO1907, holotype PDD 97532, isotype S F140316, GenBank KJ635237 (ITS+LSU).
Description. Pileus 12–30 mm diameter, conical, later broadly conical or expanded, dry, hygrophanous, orange with an orange-brown tinge on disk, very finely innate-fibrillose, margin concolorous, somewhat striate. Lamellae intensely orange when young, free, distant (L = 24, l = 1–2). Stipe 20–40 × 2–4 mm, dry, cylindrical, greyish yellow to orange-yellow, with sparse orange-brown fibrils near base. Veil orange-brown, sparse, cortina not noted. Context yellow-orange to orange-brown. Macrochemical reactions: NaOH blood red on all parts, fluorescence in UV-light none. Odour none, taste weakly raphanoid.
Spores (6.8−)7.1−7.5−8(−8.4) × (5.5−)5.7−6.1−6.4(−6.5) µm, Q = 1.25 ± 0.07 (n = 26), subglobose, moderately verrucose. Marginal elements scattered, clavate to vesiculose, hyaline, 18–28 × 7–9 µm. Basidia 25–30 × 7–8 µm, some ± capitate with a neck, many with yellow contents, four-spored. Pileipellis thin of repent hyphae, 4–5 µm wide. Hypocutis of oblong to fusoid elements, 50–100 × 15–30 µm, many with a yellow pigmentation. Clamp connections present.
Habitat. Gregarious, very rare, associated with Nothofagus spp.
Etymology. From Māori ārani ‘orange’ and iti ‘small’, due to the general aspect.
Comments. A small but spectacular dermocyboid species, recognised from its intense orange colours all over. Cortinarius aurantiellus (E. Horak) G. Garnier, rare in the same habitat, is more fibrillose, has larger spores, and exhibits an olive-coloured alkaline reaction. Cortinarius papaver Soop is larger, with more red coloration, and its spores are subglobose. Available molecular data (cf. above) indicate that C. araniiti, unlike the two preceding species, is not closely related to section Icterinula but nests in or near section Limonii Kühner & Romagn.
Acknowledgements
I wish to extend my gratitude to Peter Johnston and his team in Landcare Research, Auckland, for sequencing most of my material, and to Bruno Gasparini, Trieste for reviewing the Latin text.
Related Research Data
References
- Bougher N, Hilton R 1989. Three Cortinarius species from Western Australia. Mycological Research 93: 424–428. 10.1016/S0953-7562(89)80035-8
- Brandrud TE, Lindström H, Marklund H, Melot J, Muskos S 1989–1997. Cortinarius Flora Photographica. Matfors, Sweden, Cortinarius HB.
- Cleland J, Harris R 1948. Illustration and description of Australian fungi. 1. Agaricus and Cortinarius with special reference to antibiotic species. Records of the South Australia Museum 9: 43–56.
- Crosby TK, Dugdale JS, Watt JC 1998. Area codes for recording specimen localities in the New Zealand subregion. New Zealand Journal of Zoology 25: 175–183. 10.1080/03014223.1998.9518148
- Garnica S, Weiß M, Oberwinkler F 2002. New Cortinarius species from Nothofagus forests in South Chile. Mycologia 94: 136–145. 10.2307/3761853
- Garnica S, Weiß M, Oertel B, Oberwinkler F 2005. A framework for a phylogenetic classification in the genus Cortinarius (Basidiomycota, Agaricales) derived from morphological and molecular data. Canadian Journal of Botany 83: 1457–1477. 10.1139/b05-107
- Gasparini B 2001a. A contribution to the knowledge of Cortinarius and allied genera of Southern Tasmania, Australia 1. Australasian Mycologist 20: 49–54.
- Gasparini B 2001b. Cortinarius vinosipes Gasparini, sp. nov. Australasian Mycologist 20: 87–91.
- Gasparini B 2007. Genus Cortinarius, subgenus Phlegmacium in Tasmania. New Zealand Journal of Botany 45: 155–236. 10.1080/00288250709509711
- Gasparini B, Soop K 2008. Contribution to the knowledge of Cortinarius [Agaricales, Cortinariaceae] of Tasmania (Australia) and New Zealand. Australasian Mycologist 27: 173–203.
- Heim R 1951. Notes sur la flore mycologique des terres du Pacifique Sud. II. Cuphocybe, nouveau genre néozélandais d'Agarics ochrosporés. Revue Mycologique 16: 3–1.
- Horak E 1973. Fungi Agaricini Novazelandiae V Cuphocybe Heim (1951). Beiheft zur Nova Hedwigia 43: 193–200.
- Horak E 1979. Fungi, Basidiomycetes, Agaricales y Gasteromycetes secotioides. Flora Criptogámica de Tierra del Fuego 11 : 346.
- Horak E 1987. New species of Dermocybe (Agaricales) from New Zealand. Sydowia 40: 81–112.
- Horak E, Taylor M 1981. Fungi Agaricini Novazelandiae XI. Rozites Karsten. New Zealand Journal of Botany 19: 353–360. 10.1080/0028825X.1981.10426392
- Moser M 1983. Kleine Kryptogamenflora, Band II 2/b. Stuttgart, Gustav Fischer Verlag.
- Moser M, Horak E 1975. Cortinarius Fr. und nahe verwandte Gattungen in Südamerika. Beiheft zur Nova Hedwigia 52: 1–607.
- Peintner U, Horak E, Moser M, Vilgalys R 2002. Phylogeny of Rozites, Cuphocybe and Rapacea inferred from ITS and LSU rDNA sequences. Mycologia 94: 620–629. 10.2307/3761713
- Peintner U, Moncalvo JM, Vilgalys R 2004. Toward a better understanding of the infrageneric relationships in Cortinarius. Mycologia 96: 1042–1058. 10.2307/3762088
- Soop K 1998. Notes et observations sur les champignons cortinarioïdes de Nouvelle-Zélande. Documents Mycologiques 112: 13–26.
- Soop K 2001. Contribution à l’étude de la mycoflore cortinarioïde de Nouvelle-Zélande. Bulletin de la Société Mycologique de France 117: 91–132.
- Soop K 2002. Contribution à l’étude de la mycoflore cortinarioïde de Nouvelle-Zélande, II. Bulletin de la Société Mycologique de France 118: 173–194.
- Soop K 2005. A contribution to the study of the cortinarioid mycoflora of New Zealand, III. New Zealand Journal of Botany 43: 551–562. 10.1080/0028825X.2005.9512974
- Soop K 2010. A contribution to the study of the cortinarioid mycoflora of New Zealand, IV. Bresadoliana 1: 17–34.
- Soop K 2013. A contribution to the study of the cortinarioid mycoflora of New Zealand, V. Australasian Mycologist 31: 1–9.
- Soop K, Gasparini B 2011. Europe and the South Pacific: a comparison of two Cortinarius floras. Journal des JEC 13: 99–106.
- Stefani F, Jones RH, May TW 2013. Concordance of seven gene genealogies compared to phenotypic data reveals multiple cryptic species in Australian dermocyboid Cortinarius (Agaricales). Molecular Phylogenetics and Evolution 71: 249–260.