
Abstract
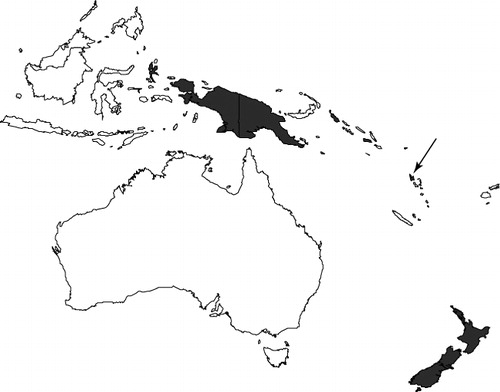
A plant specimen collected in Vanuatu (southwest Pacific), which was initially identified as Alphitonia (Rhamnaceae) because of its fruit morphology, was found through molecular phylogenetic analysis to in fact belong to Carpodetus (Rousseaceae), a genus previously reported only from New Zealand, New Guinea and the Solomon Islands. The plant appears relatively distinct genetically from both species currently accepted in this genus, C. serratus and C. arboreus. However, we cannot exclude the possibility that it belongs to one of the taxa currently placed in the synonymy of the polymorphic C. arboreus. Beyond the range extension of the genus, this observation also stresses the need for a revision of Carpodetus, particularly in New Guinea.
Introduction
Rousseaceae sensu APG (Citation2009) is an assemblage of four poorly known genera: the Mauritian endemic Roussea Sm. (monotypic), two Australian endemics Abrophyllum Hook f. (two species) and Cuttsia F. Muell. (monotypic), and Carpodetus J.R. Forst. & G. Forst. (two or more species). The latter genus has been reported from New Guinea, the Solomon Islands and New Zealand (Gustafsson & Bremer Citation1997; Gustafsson Citation2007) and has also been collected from the islands of Bougainville and Halmahera (Moluccas). The affinities of these four genera have mostly been uncovered using molecular phylogenetics (Gustafsson & Bremer Citation1997; Lundberg Citation2001). Alternative placements for Carpodetus in the past included Grossulariaceae, Escalloniaceae and Carpodetaceae.
The systematics of Carpodetus (syn. Argyrocalymma K. Schum. & Lauterb.) is somewhat obscure. International Plant names Index (IPNI Citation2013) lists a total of 13 specific epithets for the genus Carpodetus. Reeder (Citation1946) recognised at least six species in New Guinea in addition to C. arboreus (K. Schum. & Lauterb.) Schltr. and also described C. amplus Reeder, apparently known only from the type, collected on Guadalcanal, Solomon Islands. By contrast, van Royen (Citation1983) recognised only two species, C. serratus J.R. Forst. & G. Forst. from New Zealand and C. arboreus. In the latter, he included all described species based on material from New Guinea, as well as C. amplus. Van Royen did not discuss the taxonomy of C. arboreus except for noting that it is a ‘very variable species in almost all details’ (p. 2512). Carpodetus arboreus sensu van Royen is particularly variable in the size and shape of its leaves and in its indumentum.
During ‘Santo 2006’, a multidisciplinary expedition to Vanuatu lead by Institut de Recherche pour le Développement (IRD), Muséum National d'Histoire Naturelle, Paris (MNHN) and Pro-Natura (a non-governmental organisation), new plant material was collected from Santo, also known as Espiritu Santo, the largest and highest island of this archipelago. Several specimens received preliminary identifications as Alphitonia sp. (Rhamnaceae), because they had simple leaves and dark, globular, leathery fruits, c. 6–6.5 mm in diameter, with an annular scar near their widest point and the lower part of the fruit suggesting a leathery cupule. The molecular evidence presented here, however, suggests an affinity with Carpodetus.
Materials and methods
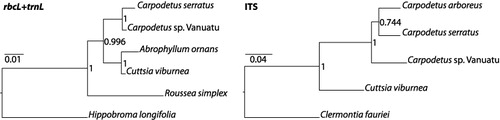
For one specimen, Pillon et al. 571, we sequenced three loci using lab protocols from Richardson et al. (Citation2000) and Pillon et al. (Citation2007): (1) the chloroplast-coding rbcL, (2) the chloroplast trnL region (trnL intron and trnL–trnF intergenic spacer) and (3) the nuclear ribosomal transcribed spacer ITS (Genbank accession numbers: KF780482, KF780483 and KF780481). Alignment of these DNA sequences with other accessions of Alphitonia was problematic, and subsequent BLAST searches in Genbank indicated affinities with Carpodetus. We aligned the sequences from this plant with those available for Rousseaceae in Genbank (see Appendix) and one species of the sister family Campanulaceae as an outgroup. The best-fit evolution models for rbcL, trnL and ITS were GTR + I, GTR + γ and GTR + γ, respectively, according to jModeltest analyses (Posada Citation2008). We conducted two Bayesian phylogenetic analyses using MrBayes (Ronquist & Huelsenbeck Citation2003), one combining rbcL and trnL, and one using the single ITS locus, and ran 1,000,000 generations of Markov Chain Monte Carlo (MCMC), sampling one tree every 1000 generations, using the gene-specific substitution model and default settings otherwise. Loci were unlinked in the combined rbcL + trnL analyses. Convergence of runs and adequate MCMC sampling were checked using Tracer (Rambaut & Drummond Citation2007). The burn-in period per run was then set to 250,000 generations for both datasets. The remaining trees were used to construct a half-compatible consensus tree and its associated estimated Bayesian posterior probabilities ().
Results
The phylogenetic analysis based on rbcL + trnL DNA sequences () strongly supported the placement of the Vanuatu plant within Rousseaceae as sister to C. serratus (posterior probability of 1). In the ITS analysis (), where sequences were available for both the currently recognised species of Carpodetus, the plant was recovered as sister to C. arboreus (New Guinea) + C. serratus (New Zealand). This group of three terminals is strongly supported (posterior probability of 1), although their exact relationships are not confidently resolved.
Discussion and conclusions
The phylogenetic analysis of the plant from Vanuatu places this specimen unambiguously in the genus Carpodetus, and this represents an important range extension for the genus and the family Rousseaceae (). Re-examination of the herbarium specimen, Pillon et al. 571, agreed with its placement in Carpodetus. Externally, the fruits of Alphitonia and Carpodetus are quite similar; both are globose with a ring-scar at or just below the widest point. In most species of Alphitonia, however, the black pericarp contains a reddish spongy to powdery mesocarp surrounding three woody endocarpids, each of which dehisces to reveal a single, erect, often persistent, arillate seed (Bean Citation2010). The fruits of Carpodetus, by contrast, are indehiscent berries with numerous small angular seeds (van Royen Citation1983; Gustafsson Citation2007). Two other specimens collected during the same expedition, Munzinger 3720 and Munzinger 3896, also initially identified as Alphitonia, are conspecific with Pillon et al. 571. A further specimen, from the Royal Society New Hebrides expedition in 1971, Gillison & Beveridge RSNH 3524 (K and P), is also from Santo and belongs to the same taxon. This specimen has ‘?Carpodetus’ typed on the collectors' label, although P.S. Green later annotated the sheet as ‘unknown’. A scanned-image of the duplicate at P is available online (http://sonneratphoto.mnhn.fr/2011/09/29/10/P03616858.jpg).
The morphological variation within these four specimens is within the accepted range of the variable C. arboreus. It is not possible to ascertain the taxonomic status of Carpodetus sp. Vanuatu any further because of the taxonomic uncertainty within C. arboreus. There are more than 40 base differences between the ITS sequence from the plant from Vanuatu and the sequences available in Genbank for the two species currently recognised, C. arboreus and C. serratus, suggesting that the former may represent a third species. However, we cannot exclude the possibility that it could be identical to one of the taxa described from New Guinea or the Solomon Islands currently placed in synonymy with C. arboreus. The genus Carpodetus therefore seems in need of a taxonomic revision that better reflects the morphological diversity present in New Guinea and the Pacific Islands.
Material seen: Vanuatu, Santo
Gillison & Beveridge RSNH 3524 (K!, P[image]! [http://sonneratphoto.mnhn.fr/2011/09/29/10/P03616858.jpg, accessed 1 December 2013]) Mt Tabwemasana, 5800 ft (c. 1770 m), wind damaged and slightly stunted forest. 27 August 1971 (fr.).
Munzinger 3720 (MO, NOU!, PVNH, SUVA) Penaorou Valley, 166°39′21.69″E 14°58′0.17″S, 900 m, montane forest, volcanic rocks. Tree 15 m. 7 November 2006 (fr.).
Munzinger 3896 (CANB, MO, NOU!, PVNH, SUVA) Penaorou Valley, 166°39′21.69″E 14°58′0.17″S, 900 m, montane forest. Tree 5 m. 12 November 2006 (fr.)
Pillon, Sam, Plunkett & Rouhan 571 (NOU!, P, PVNH) Mt Tabwemasana 166°43′40″E 15°20′10″S, 1000 m, grassy ridge (‘white grass’). Tree 15 m. 6 November 2006 (fr.)
Acknowledgements
The sample from Vanuatu was collected during the expedition Santo ‘2006’ (IRD/MNHN/Pro-Natura). We thank the curator of NOU for access to the collection; the core genetics facility at the University of Hawai‘i at Hilo; two anonymous reviewers and the associate editor for comments on an earlier version of the manuscript.
References
- Angiosperm Phylogeny Group. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161: 105–121. 10.1111/j.1095-8339.2009.00996.x
- Bean AR 2010. A revision of Alphitonia (Rhamnaceae) for Australia. Muelleria 28: 3–17.
- Gustafsson MHG 2007. Carpodetaceae. In: Kadereit JW, Jeffrey C eds. Flowering plants. Eudicots. Asterales. Berlin-Heidelberg, Springer-Verlag. Pp. 57–60.
- Gustafsson MHG, Bremer K 1997. The circumscription and systematic position of Carpodetaceae. Australian Systematic Botany 10: 855–862. 10.1071/SB96030
- International Plant Names Index 2013. http://www.ipni.org (accessed 1 July 2013).
- Lundberg J 2001. The asteralean affinity of the Mauritian Roussea (Rousseaceae). Botanical Journal of the Linnean Society 137: 267–276. 10.1111/j.1095-8339.2001.tb01122.x
- Pillon Y, Fay MF, Hedren M, Bateman RM, Devey DS, Shipunov AB et al. 2007. Evolution and temporal diversification of western European polyploid species complexes in Dactylorhiza (Orchidaceae). Taxon 56: 1185–1208. 10.2307/25065911
- Posada D 2008. jModelTest: phylogenetic model averaging. Molecular Biology and Evolution 25: 1253–1256. 10.1093/molbev/msn083
- Rambaut A, Drummond AJ 2007. Tracer v.1.4 [Computer program].
- Reeder JR 1946. Notes on Papuasian Saxifragaceae. Journal of the Arnold Arboretum 27: 275–288.
- Richardson JE, Fay MF, Cronk QCB, Bowman D, Chase MW 2000. A phylogenetic analysis of Rhamnaceae using rbcL and trnL-F plastid DNA sequences. American Journal of Botany 87: 1309–1324. 10.2307/2656724
- Ronquist F, Huelsenbeck JP 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574. 10.1093/bioinformatics/btg180
- van Royen P 1983. The alpine flora of New Guinea. Vol. 4. Cramer, Vaduz.
Appendix
Genbank accession numbers for previously published DNA sequences of rbcL, trnL and ITS used in this study:
Abrophyllum ornans: AF299090, GQ984068, –; Carpodetus arboreus: –, –, DQ499091; Carpodetus serratus: Y08461, AJ430971, DQ499090; Clermontia fauriei: –, –, JX500338; Cuttsia viburnea: Y10676, GQ984071, DQ499092; Hippobroma longiflora: DQ356140, DQ356206, –; Roussea simplex: AF084477, AJ430977, –.