
ABSTRACT
Fourteen new species are described from New Zealand, Cortinarius amblyonis, Cortinarius chrysoconius, Cortinarius cremeorufus, Cortinarius cypripedii, Cortinarius dulcamarus, Cortinarius entheosus, Cortinarius lachanus, Cortinarius mycenarum, Cortinarius mysoides, Cortinarius psilomorphus, Cortinarius trichocarpus, Cortinarius urbiculus, Cortinarius vitreofulvus and Cortinarius xenosmatoides. Also two new sections, Cortinarius sect. Cremeolinae and Cortina-rius sect. Cycnei, are proposed, and a new form of Cortinarius lubricanescens is described. The Patagonian taxon Cortinarius rapaceus var. luridus is raised to species rank under the replacement name Cortinarius austrorapaceus.
Introduction
The agaric mycota of New Zealand contains many taxa that appear to form natural groups based on both morphological and genetic similarity. This is especially the case within the genus Cortinarius (Pers.) Gray. For instance, the southern species Cortinarius chalybaeus Soop (New Zealand) and Cortinarius australis Gasparini (Australia), are part of the bihemispherical section Purpurascentes M.M. Moser (Soop Citation2002; Gasparini Citation2007; Saar et al. Citation2014). Another interesting group, including Cortinarius cremeolina Soop (New Zealand), consists of fungi with a morphology similar to those of the boreal section Multiformes (R. Henry) Moënne-Locc. & Reumaux. One aim of this seventh instalment in a series on Cortinarius taxa from New Zealand is to investigate the affinities within the Cremeolina group and describe its new taxa. The group may be placed in subg. Phlegmacium (Fr.) Trog as morphologically defined by Brandrud et al. (Citation1989). A second aim is to investigate a group of cortinarii that may be placed in subg. Myxacium (Fr.) Trog. The group has a circum-Antarctic distribution and is centred around Cortinarius magellanicus Speg.
Other species treated in the present study may be placed in subgenera Dermocybe (Fr.) Trog and Telamonia (Fr.) Trog, in the above morphological sense. Species in these subgenera were partly considered in the previous instalments (Soop Citation2001CitationCitationCitation–Citation2014), and by Horak (Citation1987), Horak and Wood (Citation1990), Soop (Citation1998), and Gasparini and Soop (Citation2008), but have not been otherwise treated in publications aimed at the mycota of the country. The infrageneric geotaxonomy of Cortinarius, and in particular of the studied groups, was extensively discussed in Soop and Gasparini (Citation2011).
Methods
Phylogenetic reconstruction
A cladogram was produced to illustrate the new sections. Relevant sequences were downloaded from GenBank, while some were generated in this study (for methods, see Soop et al. Citation2016). When separated, internal transcribed spacer (ITS) and large subunit (LSU) sequences were manually catenated, whereas in a few cases only ITS was available. The small data set (33 samples) was then aligned with Mafft v. 7 (http://mafft.cbrc.jp/alignment/server/), using the E-INS-i option. The alignment was trimmed after the begin-ITS motif and before the end-LSU motif using Mega 5.2.1 (Tamura et al. Citation2011). The resulting alignment was 1380 positions long. A Newick tree was generated with Raxml (Stamatakis Citation2014) and converted to a cladogram in pdf format with Mega 5.2.1.
Studied taxa
All collections under Typification and Other collections examined were made in the native forests of New Zealand in the years 1997–2015, under the appropriate permits issued by the Department of Conservation.
Species mentioned for comparison under Comments are described from New Zealand unless otherwise specified. All holotype material has been deposited in the PDD fungarium, Auckland, New Zealand. The GenBank accession numbers and fungarium numbers are given under Typification and Other collections examined. The descriptor ‘CO’ indicates the author’s collection number. Collection localities are referred to the districts defined by Crosby et al. (Citation1998).
Morphology
In the descriptions, measurements of the basidiome pertain to adult specimens; these are the diameter of the pileus, the length of the stipe, and the diameter of the upper part of the stipe. Lamella colour pertains to immature individuals unless specified otherwise, the designation ‘L=’ means the number of lamellae reaching the stipe, and ‘l=’ the number of lamellulae between two lamellae. The universal and partial veils of the basidiome are referred to as ‘veil’ and ‘cortina’, respectively. The alkaline reaction was made with a 30% NaOH solution.
Microscopic observations were made principally on material mounted in 5% ammoniacal solution, with or without Congo red, and examined with an oil-immersion objective (1000 ×). The term ‘marginal elements’ refers to sterile cells that are found on the lamellar edge. The spore measurements are expressed as the mean value with one-sigma limits and, in parentheses, the observed maxima and minima. The Q value is the length/width quotient; its average and standard deviation are reported. The n value is the number of measurements.
Taxonomy
Cortinarius sectio Cremeolinae Soop sect. nov.
MycoBank 814847
Basidiomata
Agaricoid or secotioid. Pileus viscid, ± glabrous with cream to brown, often pale hues. Lamellae/gleba white or pale grey when young. Stipe/columella dry, whitish, with ± distinctive, sometimes wide and marginate bulb. Universal veil white. Odour usually melleous. Spores elliptic to amygdaloid, moderately to weakly verrucose. In native forests, associated with Nothofagaceae or Myrtaceae plants. Australasia and Patagonia.
Typification
Cortinarius cremeolina Soop Citation2001, typus sectionis. In Bulletin de la Société Mycologique de France 117(2) p. 103.
Currently known taxa in the section (with area of provenance):
Comments
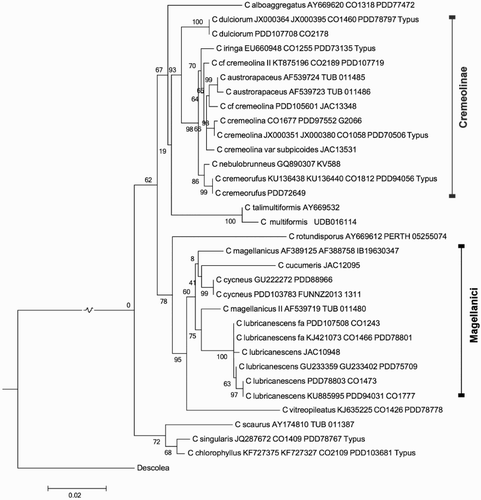
This is an austral section of phlegmacioid fungi, morphologically similar to the boreal sect. Multiformes. The section is monophyletic, forming a sister clade to Multiformes (; cf. Brandrud et al. Citation2014; Liimatainen et al. Citation2014). The Australian taxon is the only secotioid member.
The bootstrap support for the section is 93%. Several New Zealand taxa exhibit small incremental genetic distances (), suggesting an ongoing sympatric speciation. For example, the type of C. iringa differs from that of C. cremeolina by nine nuclear substitutions (ITS + LSU), whereas the variety subpicoides differs by four (ITS).
Key to sect. Cremeolinae.
Cortinarius austrorapaceus Soop nom. et stat. nov.
MycoBank 814848
Basionym
Cortinarius rapaceus var. luridus M.M. Moser in Horak & Moser Citation1975. Beiheft zur Nova Hedwigia 52, p. 92.
Comments
The name C. luridus is in use (Henry 1969). The Patagonian taxon was described as a variety of the European C. rapaceus (syn. C. foetens M.M. Moser), which belongs to sect. Caerulescentes.
Cortinarius cremeorufus Soop, sp. nov.
(C and C)MycoBank 814846
Figure 1. Basidiomata. A, Cortinarius amblyonis; B, Cortinarius chrysoconius; C, Cortinarius cremeorufus; D, Cortinarius cypripedii; E, Cortinarius dulcamarus; F, Cortinarius entheosus.

Typification
New Zealand. Otago, Waipori Falls, in Kunzea forest, 15 May 2008, P. Leonard KS-CO1812, holotype PDD 94056, isotype S F93253. GenBank KU136438(ITS), KU136440 (LSU).
Etymology
From C. cremeolina and Latin rufus ‘red-brown’, due to the general appearance.
Description
Pileus 30–40 mm diameter, rounded, later convex, weakly viscid, not hygrophanous; amber to pale red-brown, later greyish yellow, sometimes with an orange tinge and sparse white tufts at disc, finely innate fibrillose; margin somewhat paler, not striate. Lamellae pale grey when young, crowded. Stipe 50 × 8 mm, with a marginate bulb; pale tan, sometimes with a rudimentary white volva. Veil white, sparse, as a coating on stipe bulb. Context pale tan to almost white. Odour none or faintly sweet; taste none. Macrochemical reactions: NaOH weakly red-brown on cutis, else none.
Spores (8.4–)8.6–9.2–9.8(–10.6) × (4.9–)5.3–5.7–6.1(–6.5) µm, Q = 1.61 ± 0.11 (n = 25), obtusely elliptic, moderately verrucose. Marginal elements scattered, clavate to vesiculose, of varying sizes 12–17 × 7–8 and 22–28 × 7–8 µm. Basidia 25–30 × 7–9 µm, four-spored. Pileipellis with an upper layer of tangled, gelatinous hyphae, 3–5 μm wide, lower layer of repent hyphae 3–5 μm wide. Hypocutis of oblong elliptic elements, 25–60 × 8–15 μm. Clamp connections present.
Habitat
Gregarious, in myrtaceous forest, uncommon.
Other collections examined
New Zealand. Taupo, 29 Apr 2001, E. & A. Horak, PDD 72649, GenBank KT833622 (ITS + LSU).
Comments
Cortinarius cremeorufus resembles C. cremeolina, but presents an amber pileus and is the only agaricoid taxon in the section that grows with myrtaceous trees, characters that make it easy to determine.
Cortinarius lachanus Soop & M. Wallace sp. nov.
(A and G)MycoBank 814849
Figure 2. Basidiomata. A, Cortinarius lachanus; B, Cortinarius mycenarum; C, Cortinarius mysoides; D, Cortinarius psilomorphus; E, Cortinarius trichocarpus; F Cortinarius urbiculus.

Typification
New Zealand. Buller, Reefton, Murray Creek Track, in Fuscospora fusca forest, 11 May 2006, K. Soop CO1699, holotype PDD 103887, GenBank KF727361 (ITS), KF727305 (LSU).
Etymology
From Greek λαχανος ‘vegetable’, due to the general greenish coloration.
Description
Pileus 9–12 mm diameter, obtusely conical, dry, weakly hygrophanous, green to yellow-green or olive green, yellowing towards the margin when older, innate-fibrillose; margin ± plicate, later striate. Lamellae greenish yellow when young, distant, free, edge concolorous. Stipe 30–50 × 2–3 mm, cylindrical, slender, citrinous, pale turquoise at apex, brown-yellow towards base. Veil greenish, very thin; cortina not noted. Context pale green to olive green. Odour and taste insignificant. Macrochemical reactions: NaOH blood red on cutis, elsewhere brownish red; fluorescence in UV light none.
Spores (8.2–)8.5–9.0–9.5(–10.4) × (4.6–)4.9–5.2–5.5(–5.7) µm, Q = 1.75 ± 0.13 (n = 27), amygdaloid, weakly verrucose. Marginal elements crowded, cylindrical, some subcapitate, 18–25 × 6–8 µm. Basidia 25–35 × 6–8 µm, four-spored. Pileipellis of repent hyphae 3–6 μm wide. Hypocutis of oblong, ± rectangular elements, 25–45 × 8–12 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Taupo, Kaimanawa Rd, 19 May 2015, M. Wallace DSC 5337, PDD 107498.
Comments
This is a small dermocyboid fungus with a remarkably green coloration and a strong alkaline reaction, consistent with the presence of anthraquinonic pigments. Superficially it evokes species of Hygrocybe or Entoloma. According to available sequences, Cortinarius lachanus belongs to a small clade that includes the Australian species C. walkeri Cooke & Mass. (syn. C. austrovenetus Cleland) and the Patagonian species C. elaphinus M.M. Moser (data not shown).
Cortinarius mycenarum Soop sp. nov
(B and H)MycoBank 814850
Typification
New Zealand. Nelson, Abel Tasman NP, Harwoods Hole, in Lophozonia menziesii forest, 10 May 2015, K. Soop CO2185, holotype PDD 107715, isotype S F267771, GenBank KT875188 (ITS + LSU).
Etymology
From its resemblance to certain species of Mycena (Latin plur. genitive).
Description
Pileus 10 × 9 mm diameter, acutely conical, later bonnet-shaped, dry, weakly hygrophanous; dark red without a brown tinge, even when older, radially red innate-fibrillose. disc darker; margin not striate. Lamellae brownish red when young, free, distant, (L = 32, l = 1–2). Stipe 30–40 × 1–3 mm, cylindrical, pale red-brown but ± covered by reddish tufts and fibrils. Veil red, fairly copious; cortina very sparse. Context brownish red. Odour and taste insignificant. Macrochemical reactions: NaOH cherry red on cutis, red-brown on stipital veil, elsewhere red.
Spores (5.4–)5.6–6.1–6.6(–7.3) × (4.6–)4.7–5.0–5.3(–5.7) µm, Q = 1.22 ± 0.08 (n = 28), subglobose, moderately to fairly coarsely verrucose. Marginal elements crowded, cylindrical, some subcapitate or sinuous, 16–27 × 5–8 µm. Basidia 23–30 × 6–8 µm, four-spored. Pileipellis of repent, hyaline to pale orange hyphae 3–5 μm wide. Hypocutis of oblong, ± rectangular, pale yellow-brown elements, 35–65 × 8–15 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Otago Lakes, Haast Pass, Bridle Track, 28 Apr 2004, K. Soop CO1442, PDD 78785.
Comments
This small, dermocyboid fungus is easily taken for a Mycena in the field, due to its bonnet-shaped pileus and thin stipe. Cortinarius mycenarum is dark red all over (cf. M. haematopus) like a few other small cortinars in New Zealand, which all produce narrower spores. Of these, Cortinarius vinicolor (E. Horak) G. Garnier in the same habitat may be closest in habit, whereas Cortinarius cruentoides Soop grows in association with myrtaceous trees. A BLAST search reveals no close genetic relative, but available sequences indicate a position basal to several dermocyboid clades.
Cortinarius xenosmatoides Soop sp. nov.
(B and F)MycoBank 814851
Figure 3. Basidiomata. A, Cortinarius vitreofulvus; B, Cortinarius xenosmatoides; C, Cortinarius lubricanescens (violet form).

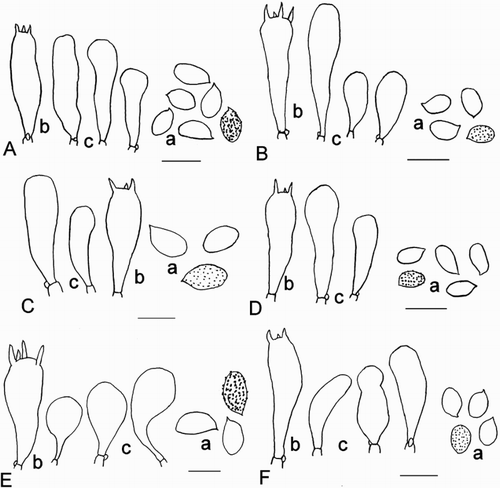
Figure 4. Microscopic details. a: spores, b: basidia, c: sterile marginal elements, scale = 10 µm. A, Cortinarius amblyonis; B, Cortinarius chrysoconius; C, Cortinarius cremeorufus; D, Cortinarius cypripedii; E, Cortinarius dulcamarus; F, Cortinarius entheosus; G, Cortinarius lachanus; H, Cortinarius mycenarum.
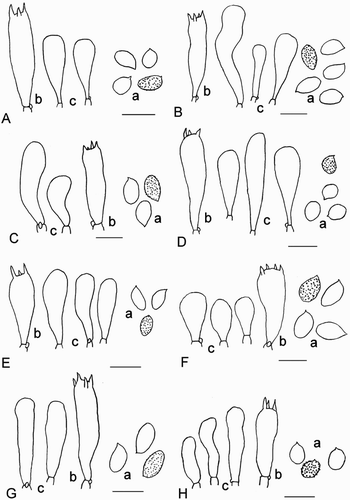
Figure 5. Microscopic details. a: spores, b: basidia, c: sterile marginal elements, scale = 10 µm. A, Cortinarius mysoides; B, Cortinarius psilomorphus; C, Cortinarius trichocarpus; D, Cortinarius urbiculus; E, Cortinarius vitreofulvus; F, Cortinarius xenosmatoides.
Typification
New Zealand. Buller, St Arnaud Range, in Fuscospora solandri forest, 4 May 2013, K. Soop CO2096, holotype PDD 103668, GenBank KF727362 (ITS), KF727318 (LSU).
Etymology
From its resemblance to C. xenosma.
Description
Pileus 20–40 mm diameter, convex, later expanded, dry, weakly or not hygrophanous, dark mahogany brown to umber, finely innate-fibrillose with pale ochraceous patches and fibrils; margin concolorous, not striate. Lamellae red-brown when young, adnate, moderately crowded to fairly distant (L = 30–40, l = 2). Stipe 30–65 × 4–9 mm, cylindrical, white to pale yellow-brown with rather dense brown fibrils, coated pale ochraceous at base. Veil greyish ochre, darkening to red-brown, fairly copious; cortina not noted. Context greyish yellow, marbled yellow-brown with a pale horn-rim near stipital cortex. Odour faint, raphanoid or like wax candles; taste none. Macrochemical reactions: NaOH none.
Spores (6.8–)7.2–7.6–8.0(–8.2) × (4.6–)4.9–5.2–5.5(–5.7) µm, Q = 1.47 ± 0.10 (n = 23), elliptical to obtusely elliptical, weakly verrucose. Marginal elements crowded, clavate, some subcapitate hyaline, 20–25 × 7–9 µm. Basidia 25–32 × 7–9 µm, four-spored. Pileipellis of repent, pale yellow-brown hyphae 4–6 μm wide. Hypocutis of oblong, pale yellow-brown elements, 40–50 × 8–16 μm, maculated by brownish encrustations. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, uncommon.
Other collections examined
New Zealand. Buller, St Arnaud Range, 4 May 2013, K. Soop CO2097, PDD 103669, S F244779; Nelson, Abel Tasman NP, Harwoods Hole, 10 May 2015, K. Soop CO2187, PDD 107717, S F267763, GenBank KT875183 (ITS + LSU).
Comments
This telamonioid fungus looks like Cortinarius xenosma Soop, but without the typical reddish tinge on the stipe. It also deviates by being more slender in average habit, and by the brick-coloured lamellae. Available molecular data show that the two species are genetically remote, placing C. xenosmatoides near several dermocyboid species, such as Cortinarius cardinalis (E. Horak) G. Garnier and Cortinarius canarius (E. Horak) Gasparini. This is remarkable, since it is a definitely telamonioid species, apparently lacking in anthraquinonic pigments as witnessed by the absent alkaline reaction.
Cortinarius trichocarpus Soop sp. nov.
(E and C)MycoBank 814852
Typification
New Zealand. North Canterbury, Arthurs Pass, Coral Track, in Fuscospora solandri forest, 20 Apr 1999, K. Soop CO1029, holotype PDD 103637, GenBank KF727354 (ITS), KF727335 (LSU).
Etymology
From Greek τριχος ‘hair’ and καρπος ‘fruit’, due to its hirsute appearance.
Description
Pileus 45–80 mm diameter, rounded, later convex-plane, dry, not hygrophanous; dark purple-brown, young ± covered by grey-brown to grey-yellow veil breaking into thick squames and patches; margin with similar tufts, not striate. Lamellae pale purple to greyish blue when young, thick, broadly adnate, fairly crowded (L = 62, l = 2). Stipe 45–65 × 10–19 mm, clavate, robust, pale grey to yellowish grey, ± covered by brownish girdles. Veil yellow-grey to brown-grey, very copious; cortina yellow-grey. Context greyish white, yellowish in pileus, marbled weakly violet. Odour and taste insignificant. Macrochemical reactions: NaOH trivial.
Spores (8.9–)9.5–10.3–11.1(–12.0) × (5.5–)5.7–6.2–6.7(–7.1) µm, Q = 1.68 ± 0.15 (n = 27), elliptical, weakly verrucose. Marginal elements crowded, subcapitate to clavate, hyaline, a few densely dark incrusted, 15–26 × 7–12 µm. Basidia 22–30 × 8–10 µm, four-spored. Pileipellis of repent hyphae, 4–7 μm wide, many with a brown vacuolar or cytoplasmic pigment. Hypocutis of ellipsoid to irregular hyaline elements, 40–70 × 20–35 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, very rare.
Comments
This is a robust species with a violaceous tint in lamellae and context and a copious, yellowish veil. It recalls Cortinarius xenosma Soop and Cortinarius cupreonatus Soop, but is generally larger with larger spores. Available molecular data place C. trichocarpus near the Rozites-like Cortinarius wallacei Soop. Moreover, the species may be regarded as an agaricoid form of a morphospecies that includes the secotioid Cortinarius ohauensis (Soop) Peintner & M. Moser, since their sequences (ITS + LSU) deviate by only four nuclear substitutions. The species has not been recorded again in New Zealand, but it was deemed worth publishing it in this study in view of its importance for the taxonomy of the genus.
Cortinarius entheosus Soop sp. nov.
(F and F)MycoBank 814853
Typification
New Zealand. Fiordland, Milford Rd, Totara Rest Area, in Lophozonia menziesii forest, 4 May 2001, K. Soop CO1210, holotype PDD 103639, GenBank KF727381 (ITS), KF727301 (LSU).
Etymology
From Greek ενθεος ‘inspired’, due to its bright and warm colours.
Description
Pileus 25–60 mm diameter, rounded, later convex to ± plane, viscid, weakly hygrophanous; tan to yellowish with a brownish orange disc, glabrous to finely innate-fibrillose; margin more greyish, flushing violet, not striate. Lamellae saturated lavender violet when young; thick, adnate, medium distant. Stipe 50–80 × 5–8 mm, cylindrical to ± fusoid, dry, white fibrillose with a faint violet sheen when young, later dirty greyish yellow; apex violet. Veil white to pale violet, fairly copious; cortina white to pale violet. Context greyish buff, marbled violet. Odour faint of Syringa flowers; taste strong but hardly unpleasant. Macrochemical reactions: NaOH none.
Spores (8.2–)8.7–9.3–9.9(–10.9) × (5.1–)5.5–5.9–6.4(–6.8) µm, Q = 1.58 ± 0.12 (n = 32), elliptic to subamygdaloid, moderately verrucose. Marginal elements crowded, vesiculose, hyaline, 18–26 × 7–9 µm. Basidia 20–24 × 7–9 µm, four-spored. Pileipellis of repent hyaline hyphae, 2–4 wide, deeper strata 5–7 µm. Hypocutis of irregular hyaline elements, 40–65 × 20–26 μm. Clamp connections present but sparse.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Otago Lakes, Kepler Track, 28 Apr 2009, K. Soop CO1880, PDD97508, GenBank KJ635234 (ITS + LSU).
Comments
This attractive but rare phlegmacioid fungus presents bright and warm colours with an apricot tinge on the pileus and violet lamellae. It recalls several other species described from the same habitat in New Zealand: Cortinarius bellus E. Horak has a viscid stipe and longer spores, Cortinarius ionomataius Soop exhibits similar colours but is dry, and Cortinarius artosus Soop has a bulbous stipe. A BLAST search has revealed no close genetic relative.
Cortinarius dulcamarus Soop sp. nov.
(E and E)MycoBank 814854
Typification
New Zealand. North Canterbury, Craigieburn, in Fuscospora solandri forest, 5 May 2009, K. Soop CO1909, holotype PDD 97534, isotype S F140318, GenBank KJ635238 (ITS + LSU).
Etymology
From Latin dulcis ‘sweet’ and amarus ‘bitter’, due to the combination of characters.
Description
Pileus 40–50 mm diameter, convex to expanded or ± plane; strongly viscid, not hygrophanous; tan to orange-brown with flammeous zones, glabrous; margin not striate. Lamellae pale cinnamon, moderately crowded. Stipe 35–45 × 7 mm, cylindrical to clavate, dry; white, flushing yellow-brown, base with a thin white coating. Veil white, sparse; cortina not noted. Context yellowish white, marbled yellow-brown. Odour strong, melleous; taste strongly bitter. Macrochemical reactions: NaOH red on brown parts of context and stipital veil, elsewhere none.
Spores (6.5–)6.7–7.3–7.9(–8.2) × (3.3–)3.6–3.9–4.2(–4.6) µm, Q = 1.88 ± 0.19 (n = 28), fusoid, pale, weakly verrucose. Marginal elements crowded, clavate, hyaline, 18–28 × 5–7 µm. Basidia 18–22 × 5–7 µm, four-spored. Pileipellis of a thin gelified layer of hyaline, repent to slightly entangled hyphae, 1.5–2.5 µm wide, deeper hyphae repent 3–4 µm. Hypocutis of rounded rectangular, hyaline elements, 40–60 × 10–15 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Nelson, Wangapeka Track, 13 May 2004, K. Soop DAO2069, OTA 60285, GenBank JX178615 (ITS).
Comments
A rare phlegmacioid species presenting generally ochraceous hues, and characterised by its remarkably narrow and fuse-shaped spores. Cortinarius dulcamarus recalls certain forms of Cortinarius orixanthus Soop, described from the same habitat in New Zealand; however, the latter produces a yellow veil and wider spores. A BLAST search reveals its close affinity to Cortinarius verniciorum Soop, another viscid, yellow-brown species with fusoid spores, but smaller and telamonioid in habit.
Cortinarius vitreofulvus Soop sp. nov.
(A and E)MycoBank 814855
Typification
New Zealand. Taupo, Cascade Hut Track, in Fuscospora fusca forest, 9 May 2009, K. Soop CO1920, holotype PDD 97545, isotype S F140326, GenBank KJ635243 (ITS + LSU).
Etymology
From the similar Cortinarius vitreopileatus and Latin fulvus ‘yellow-brown’, due to its general appearance.
Description
Pileus 35–50 mm diameter, obtusely rounded, later expanded, with a 2 mm thick, glutinous layer, hygrophanous; tan with greyish white zones when young, later darker yellow-brown, glabrous; margin paler when young, later striate. Lamellae white to grey-white when young, free, moderately crowded (L = 48, l = 3). Stipe 35–60 × 7–12 mm, cylindrical to slightly clavate, viscid, white, flavescent below apex. Veil hyaline, ± brunnescent, copious as gluten on stipe; cortina rudimentary. Context white. Odour and taste insignificant. Macrochemical reactions: NaOH reddish on brown zones of cutis, elsewhere trivial.
Spores (10.0–)10.7–11.3–12.0(–12.8) × (6.0–)6.4–6.8–7.2(–7.6) µm, Q = 1.66 ± 0.12 (n = 31), amygdaloid, strongly verrucose. Marginal elements numerous, mostly vesiculose, some clavate, often finely greyish encrusted, 18–27 × 9–12 µm. Basidia mostly finely greyish encrusted, 25–36 × 10–12 µm, four-spored. Pileipellis a thick gelified layer of entangled, hyaline hyphae, 2–3 µm wide, deeper hyphae repent 3–4 µm wide. Hypocutis of irregular, hyaline elements, 12–27 × 9–18 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, uncommon but possibly overlooked.
Other collections examined
New Zealand. Otago Lakes, Kepler Track, 28 Apr 2009, K. Soop CO1886; Taupo, 27 Apr 2001, E. & A. Horak, PDD 72615, GenBank KT833621 (ITS + LSU).
Comments
A typical cortinar in subgenus Myxacium with a thick gluten on most of the basidiome. Cortinarius vitreofulvus resembles the very common C. vitreopileatus E. Horak, which may be separated by its pale pileus colour and moderately verrucose spores. From available molecular data the genetic affinity to the latter is also evident, although its closest relative appears to be Cortinarius marmoratus E. Horak, characterised by a dark, chocolate-coloured pileus and violet lamellae. According to the protologue, Cortinarius viscostriatus E. Horak (Horak & Wood Citation1990) can be separated by its pale colours and minutely warty spores.
Cortinarius psilomorphus Soop sp. nov.
(D and B)MycoBank 814856
Typification
New Zealand. Nelson, Wangapeka Valley, in grassland close to Kunzea ericoides trees, 13 May 2004, K. Soop CO1492, holotype PDD 103885, GenBank KF727393 (ITS), KF727347 (LSU).
Etymology
From Greek ψιλος ‘smooth’ and μορφη ‘shape’, due to its general appearance.
Description
Pileus 15–40 mm diameter broadly conical with a small umbo, viscid, sticky with a ± gelatinous cutis, weakly hygrophanous; pale tan, young finely granulose to innate fibrillose; margin slightly paler, striate. Lamellae almost white when young, later pale grey-brown, somewhat decurrent. Stipe 25–50 × 2–4 mm, slender, cylindrical, waxy, pale brownish grey, base bristling from white rhizoids. Veil thin, yellow-brown; cortina pale. Context greyish yellow. Odour insignificant. Macrochemical reactions: NaOH orange to red-brown on cutis, yellow on context and stipital veil, ± red on lamellae.
Spores (5.5–)6.0–6.5–7.0(–7.6) × (4.0–)4.4–4.6–4.9(–5.1) µm, Q = 1.41 ± 0.13 (n = 30), elliptical, often obtusely, rather pale, finely verrucose. Marginal elements crowded, clavate, of varying sizes 15–32 × 7–8 µm. Basidia 22–28 × 7–8 µm, four-spored. Pileipellis thick with a gelified layer of entangled, hyaline hyphae, 1.5–3 µm wide, deeper hyphae repent 3–4 µm wide. Hypocutis of oblong irregular, hyaline elements, 20–35 × 6–10 μm. Clamp connections present.
Habitat
Gregarious, probably with myrtaceous trees, very rare.
Comments
From several phenotypic traits (growth in a lawn without evident associated host, sticky basidiomata with decurrent lamellae) this fungus was originally taken for a Psilocybe, though the spore characters readily dispelled the error. One notes that the surprising lamellae attachment may, of course, be an occasional character. The species may be placed in subgenus Paramyxacium E. Horak, where the closest taxon appears to be Cortinarius gemmeus E. Horak. But the latter deviates by an absent alkaline reaction and a strongly glutinous stipe, and the colour photo of collection PDD 27268 (E. Horak), available at the New Zealand Fungi website, clearly shows a different species. No genetic sequences have been published for the holotype of C. gemmeus, but available molecular markers place C. psilomorphus in the bihemispherical section Vibratiles Melot together with the South Pacific taxon Cortinarius melleomitis E. Horak, which may be separated by its white stipe and longer spores.
Cortinarius sectio Cycnei Soop sect. nov.
MycoBank 814857Basidiomata agaricoid or secotioid. Pileus and stipe viscid to glutinous, rarely hygrophanous, ± glabrous with white, brownish, or violet hues. Lamellae/gleba whitish or violaceous when young. Stipe cylindrical to slightly clavate, viscid. Universal veil hyaline, often with a pale violet tinge. Alkaline reaction weak or absent. Spores elliptic to amygdaloid, 9–14 µm long, moderately to rather coarsely verrucose. In native forests, mostly associated with Nothofagaceae. Australasia and Patagonia.
Typification
Cortinarius cycneus E. Horak, 1990, typus sectionis. In Horak & Wood. Sydowia 42, p. 94.
Currently known taxa in the section (with area of provenance):
Comments
This is a section of myxacioid fungi confined to the southern hemisphere. The section is monophyletic (). Even if the genetic support is modest (60%), the section is morphologically rather homogeneous. The Australian taxon is the only secotioid member, and the only member currently associated with myrtaceous plants.
Figure 6. Maximum likelihood tree of Cortinarius sect. Cremeolinae and Cycnei.
Cortinarius lubricanescens Soop Citation2001
Misapplied name
C. magellanicus Speg. ss auct. plur.
Comments
The South American species Cortinarius magellanicus has been reported repeatedly from New Zealand (Segedin & Pennycook Citation2001), but available molecular data show that it is a question of a violet form of C. lubricanescens. This form is rare, but the normal form, presenting white to greyish white colours, is fairly common. The normal form may also occasionally exhibit pale violet parts of the basidiome. According to Spegazzini’s protologue (Citation1887), the only morphological difference seems to be the lamellar colour: white with C. magellanicus, violaceous with the violet form of C. lubricanescens. Possibly, the latter form also presents a more red-lilac shade of violet.
Apparently there exists a third species, collected in South America, which has been named C. ‘magellanicus’. This taxon (labelled C. cf. magellanicus in the list) is part of sect. Cycnei, but not co-taxic with any of the other members. Moreover, a fourth taxon, collected in New Caledonia, has also been so named (F. Carriconde pers. comm.).
Collections examined
New Zealand. Normal form: Buller, St Arnaud Range, 3 May 2008, K. Soop CO1777, PDD 94031, S F93231, GenBank KU885995 (ITS); Nelson, Brightwater, Eves Bush, 11 May 2004, K. Soop CO1473, PDD78803, S F44443. Violet form: Nelson, Floral Saddle, 10 May 2004, K. Soop CO1466, PDD 78801, GenBank KJ421073 (ITS + LSU); Rangitikei, Lake Waikareiti Track, 9 May 2001, K. Soop CO1243, PDD107508.
Summary description
C. lubricanescens (violet form) (C).
Pileus 18–40 mm diameter, rounded, later convex, glutinous, not or weakly hygrophanous; violet, mostly with a red-lilac shade, disc darker, glabrous; margin often sinuate, not or weakly striate. Lamellae pale violet when young. Stipe 25–50 × 2–4 mm, cylindrical to tapering downwards, slender, tough, pale blue to violet. Veil saturated reddish violet, viscid, fairly copious, often peronate on stipe; cortina rudimentary. Context violet, sometimes paler, fragile. Odour faint, pleasant like ‘pastry’, taste insignificant. Macrochemical reactions: NaOH none or trivial. Spores amygdaloid to subelliptic, 9–11 × 5.5–6.3 µm, moderately verrucose.
Habitat
As the type, this form is found in Nothofagaceae forest.
Key to sect. Cycnei
Cortinarius mysoides Soop sp. nov.
(C and A)MycoBank 814858
Typification
New Zealand. Otago Lakes, Haast Pass, Bridle Track, in Fuscospora fusca forest, 1 May 2008, K. Soop CO1771, holotype PDD 94027, isotype S F93227, GenBank KJ635232 (ITS + LSU).
Etymology
From Greek μυς ‘mouse’ and ειδος ‘similar’, due to its greyish hues and modest size.
Description
Pileus 20–40 mm diameter, obtusely conical, later broadly conical with a shallow umbo, dry, hygrophanous; light red-brown to yellow-brown with a greyish smoky shade, rather coarsely radially innate-fibrillose with a glabrous disc; margin paler with sparse brownish fibrils, weakly striate by translucent darker streaks. Lamellae saturated brick red when young, distant to medium crowded (L = 40, l = 2). free. Stipe 60–90 × 2–5 mm, cylindrical, slender, tough, often hollow, greyish yellow with red-brown to brown tufts and girdles. Veil red-brown, fairly copious; cortina yellowish grey. Context yellow-brown marbled orange-brown. Odour raphanoid; taste nil. Macrochemical reactions: NaOH dark red to red-brown on cutis, lamellae, and stipital veil.
Spores (7.1–)7.6–8.1–8.7(–9.3) × (4.6–)4.7–5.0–5.3(–5.5) µm, Q = 1.62 ± 0.15 (n = 30), elliptical to subamygdaloid, moderately to rather strongly verrucose. Marginal elements crowded, cylindrical to clavate, hyaline, 24–30 × 6–8 µm. Basidia 24–28 × 7 µm, four-spored. Pileipellis thin with hyaline, repent hyphae, 3–4 µm thick. Hypocutis of oblong, rectangular elements, 20–35 × 6–10 μm. Clamp connections present.
Habitat
Caespitose to gregarious in Nothofagaceae forest, uncommon.
Other collections examined
New Zealand. Otago Lakes, Haast Pass, Bridle Track, 14 May 2015, K. Soop CO2191, PDD 107721, GenBank KT875189 (ITS + LSU); idem, Cameron Creek, 15 May 2015, K. Soop CO2194, PDD 107724, S F267764, GenBank KT875190 (ITS + LSU).
Comments
A small grey fungus characterised by the remarkable smoky or cloudy appearance of the cutis, especially when young, and by the reddish lamellae and reddish veil remnants on the stipe. A BLAST search reveals no close genetic relative. Cortinarius mysoides is rather similar to Cortinarius rattinus Soop and to Cortinarius rattinoides Soop, which however, both display bluish lamellae. They also exhibit different molecular markers, placing the latter in the bihemispherical section Anomali Konrad & Maubl.
Cortinarius chrysoconius Soop sp. nov.
(B and B)MycoBank 814859
Typification
New Zealand. Fiordland, Borland Lodge Track, in Fuscospora solandri forest, 25 Apr 2004, K. Soop CO1428, holotype PDD 107635, GenBank KU136438 (ITS), KU136440 (LSU).
Etymology
From Greek χρυσος ‘golden’ and κονια ‘powder’, due to the aspect of the cutis.
Description
Pileus 25–30 mm diameter, convex, dry, weakly hygrophanous; bright yellow, more yellow-brown at disc, golden yellow towards margin, finely innate fibrillose or slightly granulose, ± covered by disparate white squamules, margin with white fibrillose tufts. Lamellae golden brown, medium crowded. Stipe 50–100 × 3–4 mm, cylindrical with a small rounded bulb, coated white, partly flushing yellow-brown. Veil white, fairly copious; cortina not noted. Context pale ochre to yellow. Odour distinctly raphanoid, taste nil. Macrochemical reactions: NaOH trivial.
Spores (7.6–)8.3–9.1–9.9(–10.9) × (4.9–)5.2–5.5–5.8(–6.2) µm, Q = 1.66 ± 0.12 (n = 28), elliptical, moderately to finely verrucose. Marginal elements crowded, clavate, hyaline, 18–26 × 5–8 µm, a few differentiated (cheilocystidia) 30–38 × 7–9 µm. Basidia 25–32 × 7–9 µm, four-spored. Pileipellis thin with pale yellow, repent hyphae, 4–6 µm thick. Hypocutis of oblong, ellipsoid to rectangular, pale yellow elements, 35–70 × 10–25 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Otago Lakes, Kepler Track, 26 Apr 2004, K. Soop CO1489; Nelson, Mount Arthur, 13 May 2004, D. Orlovich DAO2002, OTA 60292, GenBank JX178607 (ITS).
Comments
This fungus, characterised by the bright golden pileus, punctuated by white granules, may evoke a Pholiota in sect. Flammans. Cortinarius peraureus Soop is somewhat similar, but lacks the granulose pileipellis structure and possesses a more robust habit. Available molecular data place C. chrysoconius in an isolated position.
Cortinarius urbiculus Soop sp. nov.
(F and D)MycoBank 814863
Typification
New Zealand. Fiordland, Kepler Track, in Fuscospora solandri forest, 24 Apr 2006, K. Soop CO1627, holotype PDD 88259, GenBank KJ547668 (ITS + LSU).
Etymology
From evoking a small form of the boreal species Cortinarius urbicus.
Description
Pileus 20–35 mm diameter, obtusely rounded to subconical, later convex-expanded, dry, weakly or not hygrophanous; young with a white, felty or pruinose coating that soon absorbs to grey-brown; margin paler with a silky white rim, not striate. Lamellae violet when young, sometimes greyish with a violet shade, narrowly adnexed, medium crowded (L = 40–60, l = 1–2). Stipe 20–60 × 4–9 mm, clavate to ± cylindrical, white, with a thin white pruinose coating, sometimes with a violet tinge. Veil white, sparse; cortina white to greyish white. Context white, often marbled violet, rather soft. Odour insignificant; taste ± bitter. Macrochemical reactions: NaOH and guayac trivial.
Spores (5.5–)6.3–6.9–7.5(–8.2) × (3.8–)4.0–4.3–4.6(–4.9) µm, Q = 1.61 ± 0.15 (n = 30), elliptical, moderately to finely verrucose. Marginal elements crowded, clavate, hyaline, 15–24 × 6–8 µm. Basidia 20–25 × 6–7 µm, four-spored. Pileipellis thin of repent hyphae, 4–6 µm thick. Hypocutis of oblong rectangular elements, 40–85 × 20–25 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Fiordland, Kepler Track, 16 May 2015, K. Soop CO2195, PDD 107725, S F267765, GenBank KT875180 (ITS + LSU).
Comments
A small greyish species, typically presenting a clavate stipe and a matt, slightly grainy coating. It evokes cortinars in sections Malachii (e.g. C. urbicus Fr.) and Anomali. Available molecular markers place C. urbiculus close to Cortinarius laquellus Soop, which deviates by possessing a viscid surface and a thin stipe. Both are closely related to the Lustrati clade (cf. Ammirati et al. Citation2007).
Cortinarius amblyonis Soop sp. nov.
(A and A)MycoBank 814866
Typification
New Zealand. Otago, Dunedin, Waipori Falls, in Leptospermum scoparium forest, 13 Apr 1997, K. Soop CO796, holotype PDD 103634, GenBank KF727364 (ITS), KF727300 (LSU).
Etymology
Fantasy name inspired by the similar Cortinarius obtusus (Greek αμβλυς ‘obtuse’).
Description
Pileus 15–30 mm diameter, obtusely conical, later conical-expanded, dry, hygrophanous; dark red-brown to orange-brown, thinly white frosty when young, finely innate-fibrillose; margin striate. Lamellae red-brown to cinnamon when young, free, distant (L = 30, l = 2–3). Stipe 30–40 × 3–5 mm, cylindrical to somewhat tapering downwards with a thin white coating that absorbs to pale brown. Veil white, sparse; cortina white. Context yellow-brown, paler in stipe base, often with a green-blue tinge when young. Odour faintly raphanoid or of iodine; taste insignificant. Macrochemical reactions: NaOH dark saturated red-brown on cutis and lamellae, elsewhere nil.
Spores (6.5–)6.8–7.3–7.8(–8.2) × (4.3–)4.4–4,7–5.0(–5.5) µm, Q = 1.56 ± 0.10 (n = 26), elliptical, moderately verrucose. Marginal elements crowded, clavate, hyaline, 16–28 × 6–8 µm. Basidia 25–30 × 7–8 µm, four-spored. Pileipellis thin of repent hyphae, 3–6 µm thick. Hypocutis of oblong elliptical, pale yellow-brown elements, 40–55 × 14–20 μm. Clamp connections present.
Habitat
Gregarious in myrtaceous forest, uncommon.
Other collections examined
New Zealand. Otago, Dunedin, Waipori Falls, 12 May 2008, K. Soop CO1801, PDD 94049, S F93246.
Comments
Available molecular data indicate that the species belongs to the bihemispherical section Obtusi Melot, where it is a typical member: small with a dry red-brown pileus, a whitish stipe, and a faint odour of iodoform. However, the lamellae do not exhibit the differentiated cheilocystidia that are prominent on several taxa in the section. Cortinarius amblyonis also recalls the New Zealand species Cortinarius paraoniti Soop and Cortinarius saturniorum Soop, which both grow in Nothofagaceae forest.
Cortinarius cypripedii Soop sp. nov.
(D and D)MycoBank 814867
Typification
New Zealand. Otago Lakes, Haast, Cameron Creek Track, in Fuscospora fusca forest, 15 May 2015, K. Soop CO2193, holotype PDD 107723, GenBank KT875199 (ITS + LSU).
Etymology
Due the colours recalling the orchid Cypripedium calceolus.
Description
Pileus 20–35 mm diameter, obtusely conical, later campanulate to bonnet-shaped with a wide umbo, dry, hygrophanous; warmly brown-red to dark orange, disc very dark brownish, almost black, glabrous to very finely innate-fibrillose; margin paler with fine reddish fibrils, striate. Lamellae brownish red to cinnamon when young, narrowly adnexed, distant (L = 30–32, l = 1–2). Stipe 20–50 × 3–6 mm, cylindrical, ± pointed at base, pale brownish yellow with rather dense red-brown fibrils. Veil orange brown to brick red, fairly copious; cortina white. Context dark yellow-brown to saturated brownish red. Odour raphanoid; taste insignificant. Macrochemical reactions: NaOH trivial.
Spores (5.5–)5.9–6.3–6.7(–7.1) × (4.9–)5.1–5.5–5.8(–6.2) µm, Q = 1.16 ± 0.09 (n = 26), subglobose, moderately verrucose. Marginal elements crowded, clavate, hyaline, of various sizes 12–17 × 6–7 µm or 20–32 × 4–9 µm. Basidia 24–32 × 7–8 µm, four-spored. Pileipellis of repent, pale yellow-brown hyphae, 6–8 µm. Hypocutis of rectangular elements, 45–70 × 12–20 μm. Clamp connections present.
Habitat
Gregarious in Nothofagaceae forest, rare.
Other collections examined
New Zealand. Otago Lakes, Haast, Cameron Creek Track, 2 May 2009, K. Soop CO1902, PDD 97527, S F140311.
Comments
The interesting colour combination of the pileus of this small but decorative fungus recalls certain flowers. It is further characterised by reddish lamellae and veil, and subglobose spores. Cortinarius cypripedii superficially resembles several other small, vividly coloured cortinars in the forests of New Zealand: Cortinarius viscilaetus Soop is viscid; Cortinarius palissandrinus Soop is more evenly coloured without an orange tinge; and Cortinarius ignellus presents paler pileal hues and a strong alkaline reaction. Available molecular data also show these taxa to be genetically disparate, while C. cypripedii is placed in the bihemispherical section Illumini Liimat. et al., which mainly contains red-brown taxa with subglobose spores.
Discussion
Cortinarius is the most diverse of macrofungal genera with an estimated 2700 species worldwide, present in all regions with a temperate climate (cf. Liimatainen et al. Citation2014). Since most Cortinarius species (c. 1900) have been described from Europe, the northern hemisphere tends to dominate in extant studies (cf. Peintner et al. Citation2004). It is nevertheless evident that the genus is well represented in the southern hemisphere, where it often forms a dominant element in the ectomycorrhizal forests of New Zealand during the fruiting season, and its diversity in this habitat appears to be at least as high as in the ectomycorrhizal forests of Europe (pers. obs.). The aim of the present study, as well as of the six previous instalments cited in the Introduction, has been to uncover and describe many formerly anonymous taxa, in an attempt to bridge an important taxonomical gap in the mycota of the country.
In their barcoding study, Garnica et al. (Citation2016) addressed the genus on a global scale and revealed a clade structure of c. 900 species based on the internal transcribed spacer regions of the rDNA genome. Many southern species were included, among others most species described in the instalments mentioned. The present study recovers two of the clades as new sections, and corroborates the diversity demonstrated by the barcoding study.
Acknowledgements
I wish to extend my gratitude to my colleagues Jerry Cooper (Christchurch), László Nagy, Bálint Dima (Budapest), and David Orlovich (Dunedin) for contributing material and performing part of the molecular sequencing. Also thanks to Peter Johnston and his team in Landcare Research, Auckland for sequencing the major part of my material.
Associate Editor: Dr Eric McKenzie.
Disclosure statement
No potential conflict of interest was reported by the author.
References
- Ammirati J, Garnica S, Halling RE, Mata M, Mueller GM, Carranza J. 2007. New Cortinarius species associated with Quercus and Comarostaphylis in Costa Rica. Can J Bot. 85:794–812. doi: 10.1139/B07-067
- Brandrud TE, Dima B, Schmidt-Stohn G, Bellù F, Frøslev T, Oertel B, Saar G, Soop K. 2014. Cortinarius subgenus Phlegmacium section Multiformes in Europe. J des JEC. 16:162–199.
- Brandrud TE, Lindström H, Marklund H, Melot J, Muskos S. 1989. Cortinarius Flora Photographica, vol. 1. Cortinarius HB, Matfors, Sweden.
- Crosby TK, Dugdale JS, Watt JC. 1998. Area codes for recording specimen localities in the New Zealand subregion. New Zeal J Zool. 25:175–183. doi:10.1080/03014223.1998.9518148
- Garnica S, Schön ME, Abarenkov K, Riess K, Liimatainen K, Niskanen T, Balint D, Soop K, Froeslev T, Jeppesen T, Peintner U, Brandrud TE, Saar G, Oertel B, Ammirati J. 2016. Determining threshold values for barcoding fungi: lessons from Cortinarius (Basidiomycota), a highly diverse and widespread ectomycorrhizal genus. FEMS Microbiol Ecol. in press.
- Gasparini B. 2007. Genus Cortinarius, subgenus Phlegmacium in Tasmania. New Zeal J Bot. 45:155–236. doi: 10.1080/00288250709509711
- Gasparini B, Soop K. 2008. Contribution to the knowledge of Cortinarius [Agaricales, Cortinariaceae] of Tasmania (Australia) and New Zealand. Austral Mycol. 27:173–203.
- Horak E. 1987. New species of Dermocybe (Agaricales) from New Zealand. Sydowia. 40:81–112.
- Horak E, Moser M. 1975. Cortinarius Fr. und nahe verwandte Gattungen in Südamerika. Beiheft zur Nova Hedwigia. 52:1–607.
- Horak E, Wood AE. 1990. Cortinarius Fr. (Agaricales) in Australasia. 1. Subgen. Myxacium and subgen. Paramyxacium. Sydowia. 42:88–168.
- Liimatainen K, Niskanen T, Dima B, Kytövuori I, Ammirati J, Frøslev T. 2014. The largest type study of Agaricales species to date: bringing identification and nomenclature of Phlegmacium (Cortinarius) into the DNA era. Persoonia – Mol Phylog Evolut Fungi. 33:98–140. doi: 10.3767/003158514X684681
- Peintner U, Moncalvo JM, Vilgalys R. 2004. Toward a better understanding of the infrageneric relationships in Cortinarius. Mycologia. 96(5):1042–1058. doi: 10.2307/3762088
- Saar G, Brandrud TE, Dima B, Schmidt-Stohn G, Bellù F, Frøslev T, Oertel B, Soop K. 2014. Cortinarius Untergattung Phlegmacium Sektion Purpurascentes in Europa. J des JEC. 16:140–161.
- Segedin B, Pennycook S. 2001. A nomenclatural checklist of agarics, boletes, and related secotioid and gasteromycetous fungi recorded from New Zealand. New Zeal J Bot. 39:285–348. doi:10.1080/0028825X.2001.9512739
- Soop K. 1998. Notes et observations sur les champignons cortinarioïdes de Nouvelle-Zélande. Documents Mycologiques. 112:13–26.
- Soop K. 2001. Contribution à l'étude de la mycoflore cortinarioïde de Nouvelle-Zélande. Bulletin de la Société Mycologique de France. 117:91–132.
- Soop K. 2002. Contribution à l'étude de la mycoflore cortinarioïde de Nouvelle-Zélande, II. Bulletin de la Société Mycologique de France. 118:173–194.
- Soop K. 2005. A contribution to the study of the cortinarioid mycoflora of New Zealand, III. New Zeal J Bot. 43:551–562. doi:10.1080/0028825X.2005.9512974
- Soop K. 2010. A contribution to the study of the cortinarioid mycoflora of New Zealand, IV. Bresadoliana. 1(2):17–34.
- Soop K. 2013. A contribution to the study of the cortinarioid mycoflora of New Zealand, V. Austral Mycol. 31:1–9.
- Soop K. 2014. A contribution to the study of the cortinarioid mycoflora of New Zealand, VI. New Zeal J Bot. 52(3):328–342. doi:10.1080/0028825X.2014.915223
- Soop K, Dima B, Szarkàndi JG, Cooper J, Papp T, Vágvölgyi Cs, Nagy LG. 2016. Psathyloma, a new genus in Hymenogastraceae described from New Zealand. Mycologia, in press.
- Soop K, Gasparini B. 2011. Europe and the South Pacific: A comparison of two Cortinarius floras. J des JEC. 13:99–106.
- Spegazzini C. 1887. Fungi Patagonici, Boletín de la Academia Nacional de Ciencias en Córdoba.
- Stamatakis A. 2014. RAxML version 8: a tool phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 30:1312–1313. doi:10.193/bioinformatics/btu033
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 28(10):2731–2739. doi:10.1093/molbev/msr121