
ABSTRACT
Ascomycetes specialised to live on hardened plant exudates occur worldwide, but the number of species so far described is relatively small (c.30). Particularly within the genus Chaenothecopsis (Ascomycota: Mycocaliciales), many species produce their ascomata on hardened but still relatively fresh outpourings of conifer resin or angiosperm exudate. Temperate rainforests of New Zealand provide habitat for several endemic Chaenothecopsis species, including Chaenothecopsis schefflerae, which was previously known from a single sample collected from the exudate of Schefflera digitata (Araliaceae) in the early 1980s. Here we show that C. schefflerae is neither lost nor very rare, but occurs sporadically throughout New Zealand. The fungus does not primarily grow on Schefflera but on exudate of several species of Pseudopanax (Araliaceae), also endemic to the region. We compare the morphology of the new specimens to the type specimen of C. schefflerae and provide a detailed description of the new material. Phylogenetic analyses based on nuclear ITS and LSU rDNA place C. schefflerae together with other morphologically similar Chaenothecopsis species growing on angiosperm exudates.
Introduction
The genus Chaenothecopsis Vainio belongs to the ascomycete order Mycocaliciales (Tibell and Wedin Citation2000). It is ecologically diverse and includes many highly specialised species that only produce ascomata on specific substrates. Many species grow as parasites or commensals on lichens while others are parasitic on green algae (e.g. Titov Citation2006). A number of Chaenothecopsis species occur exclusively on conifer resins or exudates of other vascular plants, and are sometimes restricted to the exudates of a single tree genus or even species (e.g. Tibell and Titov Citation1995; Tuovila et al. Citation2011a, Citation2011b; Tuovila Citation2013; Rikkinen et al. Citation2014; Selva and Tuovila Citation2016).
The genus Chaenothecopsis is well represented in cool temperate forest ecosystems of both Hemispheres, but it also occurs in the tropics (e.g. Tuovila et al. Citation2011a, Citation2014; Rikkinen et al. Citation2014). New Zealand forests support a diverse Chaenothecopsis mycota as described by Tibell (Citation1987) in his monograph Australasian Caliciales. Although some local species grow on lichen thalli, others occur on lignum of local conifers (e.g. Agathis, Dacrydium, Libocedrus, Podocarpus) or angiosperms (e.g. Nothofagus, Griselina, Laurelia, Metrosideros). However, only one Chaenothecopsis species in New Zealand has so far been found growing on plant exudates. Samuels and Buchanan (Citation1983) described the species from Schefflera digitata, a widespread small endemic tree (< 8 m tall) in the family Araliaceae. The authors named the fungus Mycocalicium schefflerae and Tibell (Citation1987) later transferred the species to the genus Chaenothecopsis on the basis of ascus and ascospore characteristics. At that time Chaenothecopsis schefflerae was still only known from the type location, namely in Whangarei, Northland, and no subsequent collections from Schefflera have been reported.
Here we show that C. schefflerae is widely distributed on the exudates of several Pseudopanax species in New Zealand. Although there are some differences in the anatomy between fungal specimens from the exudates of different host species, the presently available material does not permit the unambiguous circumscription of several discrete species. DNA sequence data obtained from the type specimen and from the fresh material also indicate that all the new specimens from different Pseudopanax species and S. digitata represent the same fungus.
Material and methods
Biological material
Chaenothecopsis specimens were collected from Pseudopanax colensoi var. ternatus Wardle (three-finger tree), Pseudopanax arboreus (Murray) Philipson (five-finger tree) and Pseudopanax crassifolius (Sol. ex A.Cunn.) K. Koch from 16 different localities in temperate broadleaf mixed forests of North and South Islands, New Zealand (; ). The New Zealand Fungarium (PDD), Landcare Research in Auckland, kindly supplied the type specimen of C. schefflerae collected from S. digitata J.R. Forst. et G. Forst. (seven-finger tree) for study. A culture of C. schefflerae (CBS 113957) was obtained from the Centraalbureau voor Schimmelcultures (CBS) in Utrecht.
Figure 1. Collection localities of Chaenothecopsis schefflerae in New Zealand. The different host species are indicated by colour.
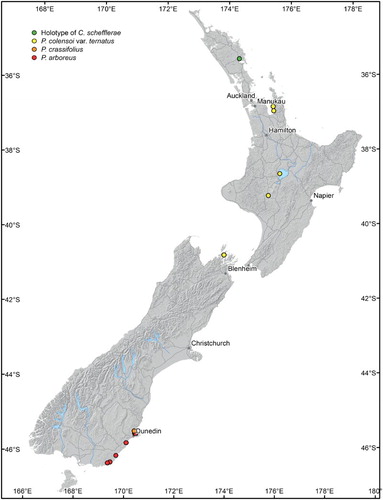
Table 1. Voucher information for the new specimens studied. All specimens are deposited in the New Zealand Fungarium (PDD) Collection in Auckland.
Light microscopy and scanning electron microscopy
Fungal specimens were studied and imaged under Carl Zeiss AxioScope A1 and Leica DMLS compound microscopes equipped with Canon EOS 5D digital cameras. In some cases, incident and transmitted light were used simultaneously (, ). Overview images of ascomata represent digitally stacked photomicrographs obtained from up to 100 focal layers merged using the software package HeliconFocus 5.0.
Morphological details of the fungal specimens were studied under a 100 × oil-immersion objective. Some diagnostic structures, like paraphyses and stipe hyphae, were observed by using potassium hydroxide (KOH), Lugol’s reagent (IKI), Melzer’s reagent (MLZ), Congo Red (CR; CR+ congophilous, colouring strongly red in CR), and nitric acid (N) as described in Tuovila (Citation2013).
For scanning electron microscopy, dry Chaenothecopsis ascomata were placed on a carbon-covered scanning electron microscope mount, sputtered with gold/palladium and examined under a Carl Zeiss LEO 1530 Gemini field emission scanning electron microscope.
Spore isolation and cultivation
Cultures of Chaenothecopsis were established using fresh field specimens collected from P. colensoi var. ternatus and P. arboreus (). For this purpose, a single ascoma was picked from the substrate and transferred to a double-cavity glass slide containing a drop of autoclaved water. All remaining substrate particles were removed in the first cavity and only the capitulum of the ascoma was transferred to the edge of the second cavity and gently crushed with a flamed needle to liberate ascospores. The available ascospores were carefully removed, without soaking the remains of the capitulum, by using a pipette, further diluted in 200–300 µL sterile 0.9% NaCl and transferred to malt yeast extract agar (MYA; 20 g malt extract, 2 g yeast extract, 20 g agar to 1000 mL distilled water, pH 6.5–7, 50 µg/mL streptomycin and penicillin G), malt extract agar (MEA, 20 g malt extract, 1 g peptone, 20 g glucose, 20 g, agar to 1000 mL distilled water, pH 5–5.5, 50 µg/mL streptomycin and penicillin G) and potato dextrose agar (PDA, Carl Roth, Karlsruhe, Germany). A spore gradient was generated by dipping a flamed loop into the spore suspension and lightly touching the media over two to three dots, from the centre of the Petri dish to the edges. The cultures are deposited at CBS of the Westerdijk Fungal Biodiversity Institute in Utrecht (reference no. CBS 113957) and in the International Collection of Microorganisms from Plants (ICMP), Landcare Research in Auckland, New Zealand (reference no ICMP 21682 (type)/ICMP 21683).
DNA extraction, PCR amplification and sequencing
DNA was successfully extracted from Chaenothecopsis ascomata growing on Pseudopanax exudates from 15 of the 17 localities () and from cultures of C. schefflerae on P. colensoi var. ternatus (ICMP 21683) and the type material (CBS 113957/ ICMP 21682). From each locality five to ten dry ascomata were gently crushed with a micro glass mortar (Carl Roth, Karlsruhe, Germany) before DNA extraction. Enzymatic DNA extraction was performed with the DNA Micro Kit from Qiagen (Hilden, Germany) by following the manufacturer’s protocol, but modifying the lysis time to 24 hours to guarantee the lysis of the fungal cells. For phylogenetic analysis we used the internal transcribed spacer (ITS) region and a partial gene of the nuclear large subunit (LSU) ribosomal RNA. The ITS region was amplified using the primers ITS4 and ITS5 (White et al. Citation1990) or ITS4 and ITS1F (Gardes and Bruns Citation1993) and the partial LSU gene was amplified with the primers LR0R (Rehner and Samuels Citation1994), LR3R (Moncalvo et al. Citation2000), LR5 and LR7 (Vilgalys and Hester Citation1990).
Polymerase chain reaction (PCR) was conducted using Taq DNA polymerase (Promega, Madison, WI) by following the manufacturer’s recommendations. The PCR conditions included the following steps: (1) hot start with 95 °C for 2 min; (2) 35 cycles of 45–60 s at 95 °C, 60 s at 52–55 °C and 45–60 s at 72 °C and (3) 10 min of final elongation at 72 °C. Subsequently, PCR products were purified using PCRapace (Invitek, Berlin, Germany) and sequenced in both directions with a MegaBACE 1000 automated sequencing machine and DYEnamic ET Primer DNA sequencing reagent (Amersham Biosciences, Little Chalfont, UK). Sequences were assembled and edited using Bioedit 5.0.9 (Hall Citation1999).
Phylogenetic analysis
The ITS and LSU rDNA was sequenced in both directions. Forward and backward sequences were assembled using Bioedit 5.0.9 (Hall Citation1999). ITS and LSU data sets were aligned separately using MAFFT version 6 (Katoh and Toh Citation2008) and subsequently combined in Bioedit 5.0.9 (Hall Citation1999). Gene regions for phylogenetic analyses were selected using Gblocks (Castresana Citation2000).
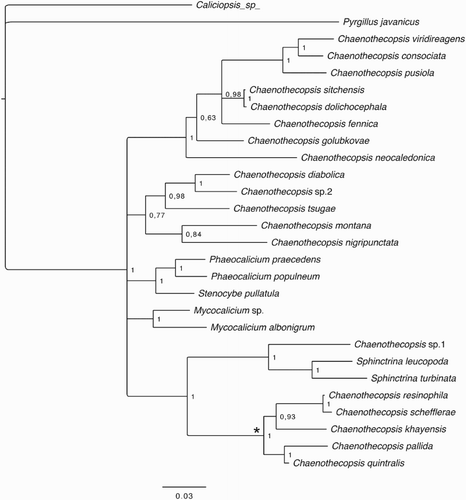
The best fitting substitution model for each gene was chosen separately from seven substitution schemes included in the software package jModeltest 2.1.1 (Darriba et al. Citation2012), and models were selected according to the Bayesian information criterion (Schwarz Citation1978). The Bayesian information criterion supported the GTR + I+G model as the best fit for the ITS region and the TIM2 + I + G model for the LSU gene. Both genes were combined in a data matrix using Bioedit 5.0.9 (Hall Citation1999) and Bayesian analyses were carried out using Markov chain Monte Carlo in MrBayes 3.1.2 (Ronquist and Huelsenbeck Citation2003). Analyses were run using four chains for 10 million generations each, sampling parameters every 1000th generation. All analyses were performed on the freely available computational resource CIPRES (www.cipres.org). Average standard deviations of split frequency < 0.01 were interpreted as indicative of independent Markov chain Monte Carlo convergence. A burn-in sample of 2500 trees was discarded for the run and the remaining trees were used to estimate branch lengths and posterior probabilities. Convergence and sufficient chain mixing (Effective sample sizes > 200) were controlled using Tracer 1.5 (Rambaut and Drummond Citation2009). GenBank accession numbers of all fungal specimens used for phylogenetic reconstruction are listed in the Supplementary material (Table S1). The combined data matrix, settings for the Bayesian analyses, and resulting phylogenetic tree () were uploaded to TreeBASE, direct access: http://purl.org/phylo/treebase/phylows/study/TB2:S20742
Results
Taxonomy
Chaenothecopsis schefflerae (Samuels & D.E. Buchanan) Tibell, Symbolae Botanicae Upsalienses 27(1): 158 (1987) [MB#131062]
Basionym: Mycocalicium schefflerae Samuels & D.E. Buchanan, N. Z. J. Bot. 21: 163 (1983)
Holotype (): New Zealand Fungarium (PDD), Landcare Research in Auckland, PDD 42005 (Isotypes: NY, ZT), communicated by I. Hood, 17 Mar 1980, Whangarei, Northland, on S. digitata.
Representative new material examined (): Chaenothecopsis specimen PDD110730, on exudate of P. colensoi var. ternatus, Beimforde, March 2015, South Island, Otago, Dunedin; Chaenothecopsis specimen PDD110728, on exudate of P. colensoi var. ternatus, Beimforde, February 2015, South Island, Otago, Dunedin; Chaenothecopsis specimen PDD110739, on exudate of P. arboreus, Rikkinen, February 2013, South Island, Catlins. These and all other specimens are deposited in the New Zealand Fungarium (PDD), Landcare Research in Auckland, with a duplicate specimen (PDD110739b) in Helsinki (H). The collection data and GenBank accession numbers are given in .
Figure 2. Chaenothecopsis schefflerae on Pseudopanax colensoi var. ternatus in New Zealand. A, Dead branch infested by several different beetle species. B, Hardened exudate with Chaenothecopsis (PDD110728). C, Semi-hardened exudate with Chaenothecopsis (PDD110739). D, Beetle pupae within larval cavities in host branch.

Figure 3. Light micrographs of Chaenothecopsis schefflerae (PDD110739) on Pseudopanax arboreus. A, Ascomata. B, Detail of ascoma. Scale bars 1 mm.

Figure 4. Scanning electron micrographs of Chaenothecopsis schefflerae (PDD110739) on Pseudopanax arboreus. A, Ascoma. B, Capitulum. C, Stipe. D, Ascospore. E–F, Semi-mature capitula. G, Cross-section of stipe. H, Stipe surface. Scale bars 100 µm (A, C), 30 µm (E, F), 20 µm (B, H), 10 µm (G) and 1 µm (D).
Description of Chaenothecopsis schefflerae from Pseudopanax exudates (): Apothecia on exudate of Pseudopanax, 680–2100 µm high. Stipe black, straight to slightly curved, 90–240 µm wide. Capitulum black, first lenticular to subspheric, later hemispheric to almost spheric, sometimes fully or partly divided into lobes, 220–650 µm wide. Asci cylindrical, 43–54 × 3.5–4.5 µm (n = 15), apex variously thickened, often penetrated by a minute or barely noticeable canal, mature asci usually without a thickening, formed without croziers. Ascospores uniseriate, obliquely to periclinally oriented in asci, aseptate, yellowish green to brown olive green, ellipsoid; released and fully mature spores with minute ornamentation, often seen as low, uneven ridges consisting of small warts, visible under light microscope at high magnification (1000×); (5.5–) 6–7.7 (–8.2) × (2.5–) 2.8–4 (–4.5) µm (n = 133, from 13 ascocarps, four populations), mean 6.6 × 3.2 µm, Q = (1.5–) 1.7–2.4 (–3), mean Q = 2.0. Paraphyses hyaline, filiform, 1.5–2.5 µm wide (n = 20), as long as or usually longer than asci, often branching several times, especially at the ascus apex level, branches intertwined and agglutinated with the hyphae of epithecium, variously covered with small crystals and/or amorphous material, both CR+; septate, septal intervals 3–14 µm. Stipe moss to olive green to dark brown, hyphae 4.5–7 µm wide, walls 1–2 µm wide, hyphae parallel and slightly intertwined (textura prismatica-intricata); inner part of the stipe hyphae light green, 5–7 µm wide, walls 1–2 µm wide, hyphae slightly intertwined (textura prismatica-intricata); whole stipe IKI+ very slowly greyish to violet red, reaction difficult to see at first because of the strong colour of the cell walls, hyphae swell in KOH and brown pigment leaks into the medium; stipe covered with dense net of arching hyphae, 2–3 µm wide. Excipulum moss to olive to brownish green, hyphae 3–6 µm wide, walls 1–2.5 µm wide, hyphae parallel (textura prismatica), covered with dense net of arching hyphae, 2–3 µm wide; IKI+ very slowly greyish to wine red, reaction difficult to see at first because of the strong colour of the cell walls. Hymenium IKI–. Hypothecium green, hyphae 5–7 µm wide, swelling in KOH. Epithecium light green to moss green, formed as a continuation of the hyphae covering the stipe and the excipulum, 3–4 µm wide, walls unevenly covered with amorphous material; IKI+ very slowly greyish red. All parts MLZ–.
The type specimen from S. digitata () differs from the new material from Pseudopanax in some anatomical details, but the taxonomic significance of these differences is impossible to evaluate because the only available material from Schefflera comes from only one population. Ascospores measured from the type seem to be on average slightly smaller (4.9–) 5.1–7.2 (–8.3) × (2.6–) 2.9–3.6 (–3.8), mean 6.2 × 3.3, Q = 1.5–2.4, mean 1.9 (n = 50, from three ascomata) than those measured from the new material. However, because of the limited material from S. digitata, a significantly smaller sample size was used for measuring the spores (50 measurements from three ascomata, whereas spore measurements from the new material are based on 133 measurements of 13 ascocarps). Another difference is seen in the excipulum, which in the type has an outer layer of relatively wide and intertwined hyphae with agglutinated walls (textura oblita-intricata). Its hymenium contains hyaline CR+ crystals, and light brown to hyaline amorphous, CR+ material, strongly gluing the asci and paraphyses together. Finally, the epithecium is composed of two layers, with an outer layer with 2–3 µm wide hyphae extending from outer excipulum, and an inner layer consisting of paraphysis tips covering the asci (compare H and D).
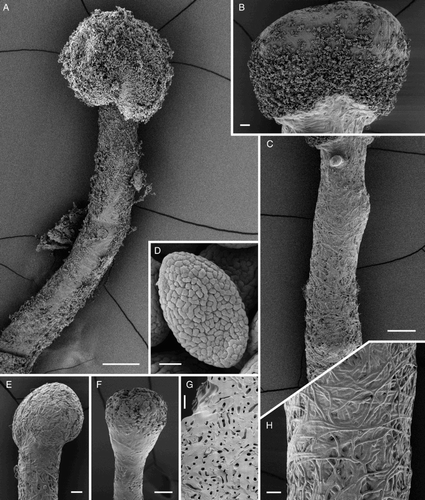
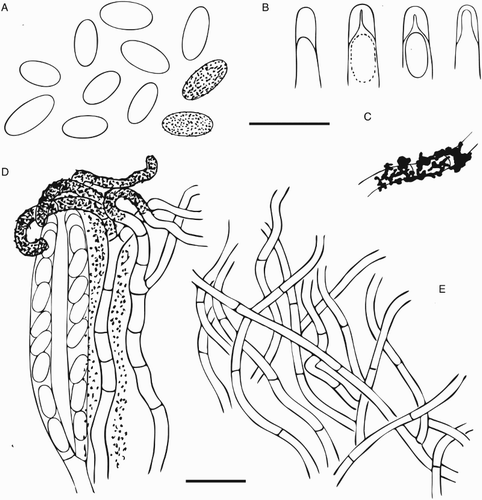
Figure 5. Ascomal structures of Chaenothecopsis schefflerae (PDD110739) on Pseudopanax arboreus. A, Ascospores. B, Ascus tips. C, Hyphae of epithecium. D, Excipulum structure. E, Epithecium structure, tips of paraphyses presented as small circles. F, Stipe hyphae. G, Paraphyses. H, Asci. All scale bars 10 µm. Drawings by Hanna Tuovila.
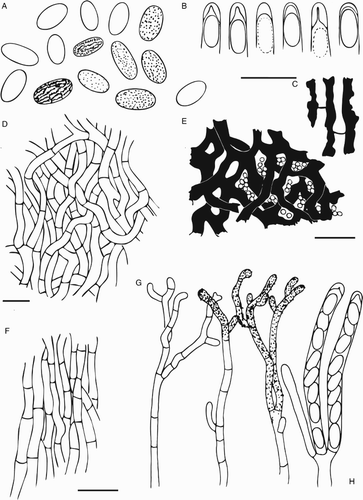
Figure 6. Light micrographs of Chaenothecopsis schefflerae (Holotype, PDD42005). A, Ascomata. B, Detail of ascoma. Scale bars 1 mm.

Figure 7. Scanning electron micrographs of Chaenothecopsis schefflerae (Holotype, PDD42005). A, Ascomata; B, Capitulum. C, Stipe surface. D, Ascospore. E, Semi-mature capitulum. Scale bars 200 µm (A), 50 µm (E), 20 µm (B,C), 1 µm (D).
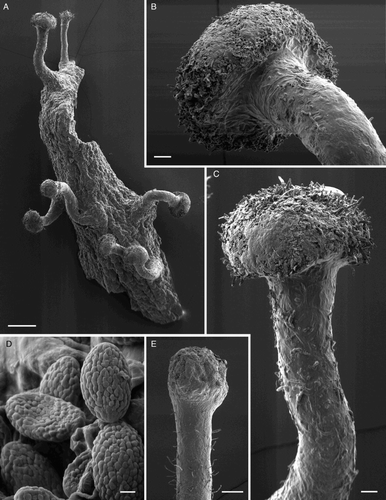
Figure 8. Ascomal structure of Chaenothecopsis schefflerae (Holotype, PDD42005). A, Ascospores. B, Ascus tips. C, Detail of paraphyse showing the ornamentation/amorphous crystals. D, Asci with spores, paraphyses and hymenial crystals. E, Excipulum structure. All scale bars 10 µm. Drawings by Hanna Tuovila.
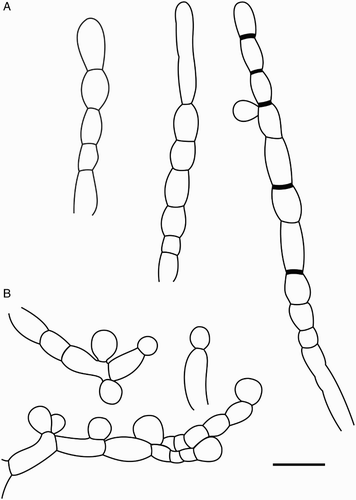
Characteristics in culture (): Germination of ascospores occurs after 2–3 days at 20 °C in the dark on MEA, MYA and PDA. Spores swelled and germinated with one or two unbranched germ tubes 2.3–3.4 µm wide and 7–20 µm long. Colonies on MEA, MYA and PDA brownish to velvety, opaque (A), appearing moist and shiny with a wrinkly surface, the surface turning undulate to notched in older colonies (B); after 2 weeks of incubation at 20 °C in the dark, colonies are 0.5–1 cm in diameter; after 3 months colonies are 1–1.8 cm; and after 1 year they are 2.5–3 cm in diameter, colonies become elevated with age. Clustered aerial hyphae arise from older colonies, often from the centre, irregularly septate, 3–3.3 µm wide, 5–15 µm long, whitish to opaque-colourless, often aggregated to form spine-like structures (C); immersed hyphae colourless 2–3.1 µm wide. No formation of apothecia on MEA, MYA or on PDA media. Conidiophores (F; B), single-celled, unbranched, 7–9.2 µm long and 4.2–5 µm wide, arising from aerial hyphae, blastic development of conidia, sometimes two conidia arise from single conidiophore. Conidia (I) translucent, globose to sub-globose to elliptic, sometimes slightly tapering at basal ends with a flattened abscission scar, size of conidia (3.6–) 4.2–9.8 (–10.7) µm × (3.6–) 4.2–7 (–7.7) µm (n = 60), mean = 4.6 × 4.2 µm, Q = 1–1.3 (–1.6) mean Q = 1.2. Chlamydospores (G–H; A) formed on aerial hyphae, often in chains, hyphal cells swell and form rounded cells with thickened cell walls, 5.8–11.1 µm long and 4.6–9 µm wide, septa sometimes thickened. Additional conidia (C–D) on aerial hyphae are sub-globose to elliptic to ovoid in shape, 12.5–28 µm long and 10–20 µm wide.
Figure 9. Chaenothecopsis schefflerae in culture (ICMP 21683) isolated from specimen PDD110730. A, Three-month-old colonies on malt extract agar. B, Detail of colony surface. C–D, Conidiophores and conidia. E, Bundles of aerial hyphae. F, Conidiophores. G–H, Chlamydospores. I, Conidia. Scale bars 2 mm (A), 1 mm (B), 50 µm (D) 20 µm, (E–F), 10 µm (C, G, H, I).
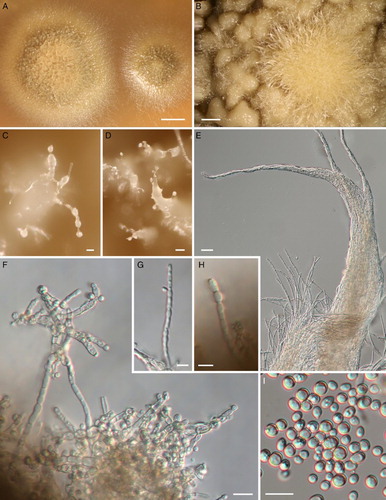
Figure 10. Reproductive structures of Chaenothecopsis schefflerae in culture (ICMP 21683). A, Production of chlamydospores. B, Conidiogenesis. Scale bar 10 µm.
Ecology and distribution: Chaenothecopsis schefflerae has to date only been found in New Zealand and is therefore most probably endemic. It has been collected from temperate broad-leaved rainforests, mainly from semi-hardened exudate and exudate-soaked exposed wood and bark of P. arboreus, P. colensoi var. ternatus and more rarely from P. crassifolius. As the combined distribution of these three species covers most of New Zealand (, ), one can expect that C. schefflerae may also occur on other exudate-producing Pseudopanax species across all forested bioclimatic zones represented in the region. The holotype was collected from exudate-soaked wood and bark of S. digitata from Whangarei, Northland, and it still remains the only record from trees other than Pseudopanax.
Discussion
Phylogenetic analysis of the ITS region indicates that all the new Chaenothecopsis specimens from Pseudopanax and cultures of the type specimen (CBS 113957/ ICMP 21682) represent a single species. In their general morphology the specimens on Pseudopanax are very similar to the type of C. schefflerae and also grow on exudates of Araliaceae species. Our microscopic analyses also demonstrate that the ascomatal anatomy of the specimens from Pseudopanax and the type material from Schefflera are very similar, and despite small differences in ascospore size and other features such as paraphyses and epithecium structure (; ), we presently assign all the new specimens from Pseudopanax to the previously described species.
Generic delimitation in Mycocaliciales is problematic and the assignment of a particular species to genera is not always possible on the basis of only one or two morphological characters (Schmidt Citation1970; Tibel Citation1984, Citation1987; Titov Citation2006; Tuovila Citation2013). Chaenothecopsis schefflerae was first described as Mycocalicium schefflerae Samules & D.E. Buchanan, but on the basis of ascospores and the ascus structure Tibell (Citation1987) transferred it to Chaenothecopsis. The ascospores of C. schefflera are ellipsoid (D; D), whereas those of Mycocalicium species are usually more or less allantoid. As previously noted by Samuels and Buchanan (Citation1983) and Tibell (Citation1987) the apices of immature asci are thickened and penetrated by a barely noticeable canal (B; B). This is a typical feature of Chaenothecopsis rather than Mycocalicium (Schmidt Citation1970; Tibell Citation1987). The stipe of C. schefflerae consists of intertwined hyphae (C, H; B, C), which is also typical of many Chaenothecopsis species, whereas the stipes of Mycocalicium species tend to consist of parallel hyphae.
Unresolved issues of generic delimitation of mycocalicioid fungi are also clearly seen in our present phylogenetic tree () in which the different genera, as traditionally circumscribed, do not form monophyletic groups. Wider taxon sampling and more sequence data from additional phylogenetic informative genes are obviously needed to resolve the correct generic relationships in this group (Hawksworth Citation1980; Tibell Citation1984; Tibell and Vinuesa Citation2005; Tuovila Citation2013). In any case, as already shown in previous studies (Tuovila et al. Citation2011a, Citation2011b, Citation2013, Citation2014; Rikkinen et al. Citation2014) the Chaenothecopsis species that grow on angiosperm exudates constitute a well-supported monophyletic group (). They also share significant morphological, anatomical and chemical traits, including the production of asci without croziers and similar, quite distinctive ascospores. Most species in this group can only be distinguished from each other by using a combination of characters (Tibell Citation1987; Tuovila Citation2013; Tuovila et al. Citation2014). On the basis of ribosomal DNA () the closest known relative of C. schefflerae is Chaenothecopsis resinophila from central China. The ascomata of these two species are indeed very similar; both have ornamented ascospores, CR+ crystals on the tips of paraphyses, and a similar type of epithecium. However, C. schefflerae differs from C. resinophila in having wider hyphae with thicker walls in the stipe and excipulum, more branched paraphyses, and slightly smaller ascospores and Q-value (Tuovila et al. Citation2014).
Figure 11. Phylogenetic relationships of mycocalicioid fungi (Mycocaliciales, Ascomycota). Bayesian tree inferred from ribosomal internal transcribed spacer region and 28S (partial) sequences. Numbers at branches indicate Bayesian posterior probabilities. Asterisk marks the monophyletic clade on angiosperm exudates.
Samuels and Buchanan (Citation1983) noted that C. schefflerae is easy to cultivate and we were able to germinate ascospores and grow mycelia on several different carbohydrate-based media (MEA, MYA, PDA; ). Unlike in the cultures obtained by Samuels and Buchanan, our cultures did not produce ascomata. This may have been due to different growth conditions; we, for example, kept our cultures in the dark whereas Samuels and Buchanan exposed their cultures to 12 h darkness/12 h near ultraviolet and fluorescent light. The conidiophores and chlamydospores produced in our cultures were about the same shape and size as those reported by Samuels and Buchanan. However, the larger conidia (C, D) were not mentioned in the original description.
Chaenothecopsis schefflerae is the only Chaenothecopsis species presently known from angiosperm exudates in New Zealand. Our new findings demonstrate that it is widely distributed on both main islands of New Zealand and can occur abundantly on exudate or exudate-soaked wood and bark of P. colensoi var. ternatus, P. arboreus and P. crassifolius. Those host trees belong to the Arboreus and Crassifolius clades, which represent two of the three major clades within New Zealand Pseudopanax (Perrie and Shepherd Citation2009). Inasmuch as the fungus has been collected only once from S. digitata, a species that rarely produces copious amounts of exudate, we suspect that the type specimen may have been collected from a relatively rare substrate, even though the type specimen is quite impressive in having numerous ascomata. Even with some effort we have not been able to find new material of C. schefflerae from Schefflera and on this basis we cannot dismiss the possibility that the type specimen might have been collected from Pseudopanax and mislabelled.
Based on our observations, C. schefflerae grows mainly in dense and humid rainforests, from the lowlands up to an altitude of 670 m, sometimes in more open secondary vegetation. We often observed beetle borings in branches with colonised exudate outpourings (A–B) and found several larvae and pupae, which specifically colonised the outer bark, the xylem, or the phloem of P. colensoi var. ternatus (D). It is likely that insect activity induces the production of exudates in Pseudopanax and that adult insects play a role in the spore dispersal of C. schefflerae. A range of weevils such as Pactola variabilis Pascone and Pactola fuscicornis Broun (Mazur et al. Citation2016), Eiratus parvulus Pascone (Kuschel Citation1982) and Ectopsis ferrugalis Broun (Gibbs Citation2009) exclusively colonise species of Pseudopanax and Schefflera digitata. These specialised and presumably ancient insect–plant associations can facilitate the dispersal of the fungal spores, guaranteeing that they reach the specific substrate essential for their germination and growth (Tuovila et al. Citation2011a). A similar situation was recently described for Resinogalea humboldtensis (Rikkinen et al. Citation2016), another resinicolous ascomycete from New Caledonia, which is closely associated with its endemic plant host but also with host-specific insects attacking the host tree and so providing resinous substrate for the fungus (Beimforde et al. Citation2017).
The close relationship of Chaenothecopsis species from angiosperm exudates, as represented by their well-supported monophyletic clade (), indicates a single origin of their unusual ecology. In contrast, there may have been multiple losses or gains of a similar ecology centreing on conifer resins, as such species are clearly polyphyletic within the Mycocalicaceae. In any case, fossilised Chaenothecopsis specimens from Palaeogene ambers provide solid evidence that the association between some Chaenothecopsis species and conifer resin is ancient and has remained essentially unchanged for at least 35 million years (Rikkinen and Poinar Citation2000; Tuovila et al. Citation2013; Beimforde et al. Citation2014). Nine mycocalicioid species are currently known to grow exclusively on angiosperm exudates (Tuovila Citation2013; Tuovila et al. Citation2014). These species share many similar morphological features, but have highly disjunct distributions in Africa, Asia, South America and New Zealand. This suggests that also the ecology of this lineage may have an ancient origin.
The closest relative of C. schefflerae is C. resinophila, a species that grows on exudate of Kalopanax septemlobus (Thunb.) Koidz, a species in the Araliaceae restricted to temperate forests of East Asia. The high consistency of the ITS barcoding genes of both species would suggest that C. resinophila and C. schefflerae may have diverged relatively recently. According to molecular age estimates (Nicolas and Plunkett Citation2014), Kalopanax appeared in the Eocene around 35 million years ago and species of Pseudopanax appeared later in the Miocene, around 17 million years ago. However, recently found fossil Pseudopanax from 23-million-year-old sediments of the Foulden Maarcrater in Otago, southern New Zealand, indicates a slightly older origin of the New Zealand Araliaceae (personal communication Jennifer Bannister). Given the fact that the vast majority of araliad species are distributed today in southern Asia, as well as the fact that Kalopanax is older than Pseudopanax, one might hypothesise that C. schefflerae and C. resinophila would have had a common ancestor that lived on exudates of Asian Araliaceae. Also, the very low intraspecific divergence within the ITS sequence of C. schefflerae would indicate that this particular species might not be ancient. From 497 base pairs of the ITS region came only one from 15 samples that differed in one base pair, which corresponds to a Kimura two-parameter distance (Kimura Citation1980) of only 0.002.
Table S1. GenBank accessions for the fungal ITS and LSU sequences used in this study for phylogenetic analysis (Figure 11).
Download MS Word (16.6 KB)Acknowledgements
We would like to thank Uwe Kaulfuß (Dunedin) for help in the preparation and execution of fieldwork in New Zealand and Phil Bishop (Dunedin) for providing access to Maud Island. We further thank Wanda Daley (Auckland) and Adrienn Stanton (Auckland) from the New Zealand Fungarium (PDD) Landcare Research for providing the type material of Chaenothecopsis schefflerae, and Melanie van der Merwe (Utrecht) and Gerard Verkley (Utrecht) from the Westerdijk Fungal Biodiversity Centre for the type strain material of C. schefflerae. We also thank Jennifer Bannister (Dunedin) and Daphne Lee (Dunedin) for information about fossil Araliaceae in New Zealand as well as Dorothea Hause-Reitner for scanning electron micrographs and Luke Easterbrook (Dunedin) for his help in GIS-mapping. We also thank two anonymous reviewers for their detailed review of the manuscript and their constructive comments.
Disclosure statement
No potential conflict of interest was reported by the authors.
Related Research Data
References
- Beimforde C, Feldberg K, Nylinder S, Rikkinen J, Tuovila H, Dörfelt H, Gube M, Jackson D, Reitner J, Seyfullah LJ, Schmidt AR. 2014. Estimating the Phanerozoic history of the Ascomycota lineages: combining fossil and molecular data. Molecular Phylogenetics and Evolution. 77:307–319.
- Beimforde C, Seyfullah LJ, Perrichot V, Schmidt K, Rikkinen J, Schmidt A. 2017. Resin exudation and resinicolous communities on Araucaria humboldtensis in New Caledonia. Arthropod-Plant Interactions. doi:10.1007/s11829-016-9475-3.
- Castresana J. 2000. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Molecular Biology and Evolution. 17:540–552. doi: 10.1093/oxfordjournals.molbev.a026334
- Darriba D, Taboada GL, Doallo R, Posada D. 2012. jModelTest 2: more models, new heuristics and parallel computing. Nature Methods. 9:772–772. doi: 10.1038/nmeth.2109
- Gardes M, Bruns TD. 1993. ITS primers with enhanced specificity for basidiomycetes - application to the identification of mycorrhizae and rusts. Molecular Ecology. 2:113–118. doi: 10.1111/j.1365-294X.1993.tb00005.x
- Gibbs GW. 2009. The end of an 80-million year experiment: a review of evidence describing the impact of introduced rodents on New Zealand’s ‘mammal-free’ invertebrate fauna. Biological Invasions. 11:1587–1593. doi: 10.1007/s10530-008-9408-x
- Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series. 41:95–98.
- Hawksworth DL. 1980. Two little-known members of the mycocaliciaceae on polypores. Transactions of the British Mycological Society. 74:650–651. doi: 10.1016/S0007-1536(80)80073-8
- Katoh K, Toh H. 2008. Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform. 92:86–98.
- Kimura M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution. 16:111–120. doi: 10.1007/BF01731581
- Kuschel G. 1982. Apionidae and Curculionidae (Coleoptera) from the Poor Knights Islands, New Zealand. Journal of the Royal Society of New Zealand. 12:273–282. doi: 10.1080/03036758.1982.10415349
- Mazur MA, Tron F, Mille C. 2016. Pactola kuscheli sp. nov. (Coleoptera: Curculionidae), a potential cause for the decline of the threatened New Caledonian conifer, Agathis montana de Laubenfels, 1969 (Araucariaceae). Austral Entomology. doi:10.1111/aen.12234.
- Moncalvo JM, Lutzoni FM, Rehner S, Johnson J, Vilgalys R. 2000. Phylogenetic relationships of agaric Fungi based on nuclear large subunit ribosomal DNA sequences. Systematic Biology. 49:278–305. doi: 10.1093/sysbio/49.2.278
- Nicolas AN, Plunkett GM. 2014. Diversification times and biogeographic patterns in apiales. The Botanical Review. 80:30–58. doi: 10.1007/s12229-014-9132-4
- Perrie LR, Shepherd LD. 2009. Reconstructing the species phylogeny of Pseudopanax (Araliaceae), a genus of hybridising trees. Molecular Phylogenetics and Evolution. 52:774–783. doi: 10.1016/j.ympev.2009.05.030
- Rambaut A, Drummond AJ. 2009. Tracer. MCMC Trace analysis tool version v1.5.0. http://tree.bio.ed.ac.uk/software/tracer/.
- Rehner SA, Samuels GJ. 1994. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycological Research. 98:625–634. doi: 10.1016/S0953-7562(09)80409-7
- Rikkinen J, Poinar G. 2000. A new species of resinicolous Chaenothecopsis (Mycocaliciaceae, Ascomycota) from 20 million year old Bitterfeld amber, with remarks on the biology of resinicolous fungi. Mycological Research. 104:7–15. doi: 10.1017/S0953756299001884
- Rikkinen J, Tuovila H, Beimforde C, Seyfullah LJ, Perrichot V, Schmidt AR. 2014. Chaenothecopsis neocaledonica sp. nov.: The first resinicolous mycocalicioid fungus from Araucariaceae. Phytotaxa. 173:49–60. doi: 10.11646/phytotaxa.173.1.4
- Rikkinen J, Beimforde C, Seyfullah, LJ, Perrichot V, Schmidt K, Schmidt AR. 2016. Resinogalea humboldtensis gen. et sp. nov., a new resinicolous fungus from new Caledonia, placed in Bruceomycetaceae fam. nova (Ascomycota). Annales Botanici Fennici. 53:205–215. doi: 10.5735/085.053.0408
- Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 19:1572–1574. doi: 10.1093/bioinformatics/btg180
- Samuels GJ, Buchanan DE. 1983. Ascomycetes of New Zealand. 5 Mycocalicium schefflerae sp. nov., its ascal ultrastructure and Phialophora anamorph. New Zealand Journal of Botany. 21:163–170. doi: 10.1080/0028825X.1983.10428540
- Schmidt A. 1970. Anatomisch-taxonoische Untersuchungen an europäischen Arten der Flechtenfamilie Caliciaceae. Mitteilungen aus dem Staatsinstitut für Allgemeine Botanik Hamburg. 13:111–166.
- Schwarz G. 1978. Estimating the dimension of a model. The Annals of Statistics. 6:461–464. doi: 10.1214/aos/1176344136
- Selva SB, Tuovila H. 2016. Two new resinicolous mycocalicioid fungi from the Acadian Forest: one new to science, the other new to North America. The Bryologist. 119:417–422. doi: 10.1639/0007-2745-119.4.417
- Tibell L. 1984. A reappraisal of the taxonomy of Caliciales. Beiheft zur Nova Hedwigia. 79:597–713.
- Tibell L. 1987. Australasian Caliciales. Symbolae Botanicae Upsalienses. 27:1–276.
- Tibell L, Titov A. 1995. Species of Chaenothecopsis and Mycocalicium (Caliciales) on Exudate. The Bryologist. 98:550–560. doi: 10.2307/3243587
- Tibell L, Vinuesa M. 2005. Chaenothecopsis in a molecular phylogeny based on nuclear rDNA ITS and LSU sequences. Taxon. 54:427–442. doi: 10.2307/25065370
- Tibell L, Wedin M. 2000. Mycocaliciales, a new order for nonlichenized calicioid fungi. Mycologia. 92:577–581. doi: 10.2307/3761518
- Titov, A. 2006. Mikokalizievye griby (porjadok Mycocaliciales) Golarktiki [Mycocalicioid fungi (the order Mycocaliciales) of Holarctic]. Moskva: KMK Scientific Press.
- Tuovila H. 2013. Sticky business – diversity and evolution of Mycocaliciales (Ascomycota) on plant exudates. Helsinki: Publications in Botany from the University of Helsinki 44. https://helda.helsinki.fi/handle/10138/39265.
- Tuovila H, Cobbinah JR, Rikkinen J. 2011a. Chaenothecopsis khayensis, a new resinicolous calicioid fungus on African mahogany. Mycologia. 103:610–615. doi: 10.3852/10-194
- Tuovila H, Larsson P, Rikkinen J. 2011b. Three resinicolous North American species of Mycocaliciales in Europe with a re-evaluation of Chaenothecopsis oregana Rikkinen. Karstenia. 51:37–50.
- Tuovila H, Beimforde C, Schmidt AR, Rikkinen J. 2013. Stuck in time – a new Chaenothecopsis species with proliferating ascomata from Cunninghamia resin and its fossil ancestors in European amber. Fungal Diversity. 58:199–213. doi: 10.1007/s13225-012-0210-9
- Tuovila H, Davey ML, Yan L, Huhtinen S, Rikkinen J. 2014. New resinicolous Chaenothecopsis species from China. Mycologia. 106:989–1003. doi: 10.3852/13-178
- Vilgalys R, Hester M. 1990. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriology. 172:4238–4246. doi: 10.1128/jb.172.8.4238-4246.1990
- White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR Protocols: a guide to methods and applications. New York: Academic Press; p. 315–322.