Abstract
Aspects of sea-bed structure and benthic-macroinvertebrate species composition, distribution, richness and diversity in coastal waters off eastern Waiheke Island, Hauraki Gulf, are reported. In contrast to the sole historical account of sea-bed community structure from this same region, no widely distributed assemblages of species are recognised throughout it; no two sites share the exact same complement of species; and almost all sites are less than 80% similar in their taxonomic composition, most considerably so. Species richness and diversity are reported to vary with substratum type and depth, and spatially; species occurring within muds are the least diverse and species rich, followed by those of muddy gravels, and then gravels; many taxa prove common to the three substratum types; and dominance of taxa is recognised to decrease with an increase in substratum complexity, from muds to gravels, and species richness. With the exception of invasive marine species, apparent changes in the composition of assemblages throughout this region over the eight-decade period that data span are considered artefacts of the way in which such assemblages were historically defined. We recommend historical accounts of sea-bed community distributions throughout Hauraki Gulf be interpreted with caution, especially when attempting to use such schematic depictions to determine whether changes have occurred in assemblage composition.
Introduction
Substantive accounts of the composition and distribution of sea-bed communities throughout Hauraki Gulf are few, limited largely to the pioneering works of Powell (Citation1937), based on sampling at 138 dredge stations undertaken between 1926 and 1936 throughout the gulf, and a broadly comparable but more geographically limited survey of the inner Waitemata Harbour and Rangitoto Channel undertaken by Hayward et al. (Citation1997), based on sampling at 150 dredge stations surveyed between 1993 and 1995. These studies have firmly entrenched in the minds of natural historians the concept that generalised assemblages of species occur throughout parts of Hauraki Gulf. However, biological data from neither of these accounts have been subject to more rigorous multivariate statistical evaluation, and as such the validity of purported species assemblages has not been demonstrated.
With the exception of that limited sea-bed sampling reported by Powell (Citation1937), sea-bed communities off eastern Waiheke Island have not been reported. On the basis of nine widely spread dredge stations, three collected in 1927 and six in 1933, Powell (Citation1937) recognised two sea-bed formations in this region: a widespread urchin (Echinocardium) formation/association; and a more-restricted-in distribution bivalve (Tawera+Venericardia (now Purpurocardia)) formation/association (). Powell appears to have differentiated his formations from his associations on the basis of the absence of a characteristic species, and secondarily on the basis of changes in species richness (Powell Citation1937: 371). However, we cannot reliably replicate this procedure, a problem exacerbated by Powell's boundaries also being somewhat intuitively defined and likely influenced by that information he had on the composition of sediments throughout the region.
Fig. 1 Site distribution and associated substratum type [hatched area depicts Tawera+Purpurocardia formation of Powell (Citation1937)]; insets: North Island, New Zealand; and Waiheke Island, Hauraki Gulf.
![Fig. 1 Site distribution and associated substratum type [hatched area depicts Tawera+Purpurocardia formation of Powell (Citation1937)]; insets: North Island, New Zealand; and Waiheke Island, Hauraki Gulf.](/cms/asset/8512e6f4-4026-4207-a352-7a9cb98e599a/tnzm_a_498088_o_f0001g.jpg)
The purpose of the survey reported herein was to establish series of control sites for a separate monitoring programme designed to evaluate the effects of mussel farming on sea-bed communities off Taniwhanui Point, rather than to critique Powell's sea-bed communities off eastern Waiheke Island (). As such, all sites surveyed by Powell were not resampled. Nevertheless, as a consequence of our study, we can report the benthos and sedimentary characteristics of the sea bed in a quantitative manner at many more sites spread over a greater area than has been previously reported. This represents the first fully quantitative evaluation of sea-bed communities in Hauraki Gulf, which we intend to augment with additional accounts of sea-bed fauna elsewhere (see the Monalisa Biodiversity Database for coverage, www.Monalisa.ac.nz).
Methods
The sea bed from 4–30 m between Cowes Bay and Kauri Point (36°46.52–49.85′S, 175°09.47–12.21′E), eastern Waiheke Island, was surveyed during February of 2008. Macrobenthic fauna was identified from 228 sea-bed samples, 102 samples (34 sites with three replicates at each) from along three transects extending from within an existing mussel farm off Taniwhanui Point to approximately 80 m outside its physical boundary, and 126 non-replicated grab sites spread throughout the greater survey region (). Nine further grabs were collected at representative sites for sediment grain-size analysis.
All samples, whether collected for biological purposes or for sediment grain-size analysis, were taken by Van Veen grab. This grab has a bite aperture of 0.0336 m2, but the depth to which it samples depends upon grain size and degree of substratum compaction; any grab was discarded in the event the sample was not at least ~75% full. Upon collection, samples for biological analyses were immediately fixed in a 5% buffered (sodium bicarbonate) formalin–seawater solution and returned to the laboratory. These were subsequently sieved over a 500-µm Endicott mesh, then species removed from the coarser fraction (>500 µm) and identified to the lowest common denominator, whether this be species or species-specific enumerated unknown, with the exception of all Nemertea, Nematoda and Oligochaeta, each treated as single taxonomic entities. A voucher collection of all taxa has been accessioned into the biological collections of Auckland University of Technology; full biological data from these sites, and those surveyed elsewhere throughout Hauraki Gulf are available online at the Monalisa Biodiversity database, www.Monalisa.ac.nz.
With the time and resources available, it was not possible to analyse sediment grain-size properties from each of the 228 biological sites that were surveyed; our compromise was to describe sediments from these sites as either predominantly mud, mud/gravel or gravel, based on a visual and tactile appraisal of the proportion of shell gravel and granule to mud and silt within it. To characterise sediment grain-size properties more accurately, nine additional samples were collected, three from locations classified as belonging to each aforementioned substratum type. Percentage volumes for sediment fractions (3.35, 1.18 and 1.0 mm, and 600, 500, 300, 150, 63, 11 and <11 µm) were determined by wet-sieving, with the grain-size properties of each sample characterised by five granulometric indices: median particle diameter (ϕ50), first (ϕ25) and third (ϕ75) quartiles, sorting coefficient ((ϕ75 − ϕ25)/2), and ϕ-quartile skewness ((ϕ75 + ϕ25)/2 − ϕ50); the degree of sorting of each sediment sample was characterised in accordance with the classification of Gray (Citation1981).
To determine if any depth-related differences were apparent in the composition of sea-bed communities, mud and mud/gravel sites were placed into three arbitrarily selected depth ranges, capturing the range in depth throughout the region: those more shallow than 10 m, those from 10–15 m, and those at depths >15 m (to 30 m); gravel sites were encountered within only two of these depth ranges, those more shallow than 10 m, and those between 10 and 15 m.
Biological data were analysed using PRIMER v6 (Clarke & Gorley Citation2006). Measures of biological diversity were presented in five indices: total individuals (n), Margalef's index of species richness (d), Pielou's evenness index (J’), Shannon–Wiener diversity index (H’), and Simpson dominance index (λ). To determine whether significant differences existed in these indices in different substrata, each was tested by ANOVA in SPSS 15.0. To determine the relationship between species dominance, substratum type, and species richness, Simpson index (λ), a dominance index, was plotted against species richness for each substratum.
A similarity matrix was constructed using square-root transformed data and the Bray–Curtis coefficient. Data were presented graphically using multi-dimensional scaling (MDS) ordinations and group average clustering (CLUSTER). Significant differences in the composition of species assemblages by depth and substratum type (groups) were determined using a randomised permutation test (ANOSIM; Clarke & Green Citation1988) on the similarity matrix. The main species contributing most to the within-group similarity between sites, or dissimilarity between groups were determined using the Similarity Percentage Routine (SIMPER; Clarke & Warwick Citation1994). The breakdown in SIMPER is based on Bray–Curtis coefficient of all pairs of samples within or between groups.
To determine to what extent our sampling effort captured the total number of species throughout the survey region, species accumulation curves were prepared using pooled biological data for all substrata throughout the survey region, and separately for each substratum type (), using the ‘Species-Area plot’ in PRIMER.
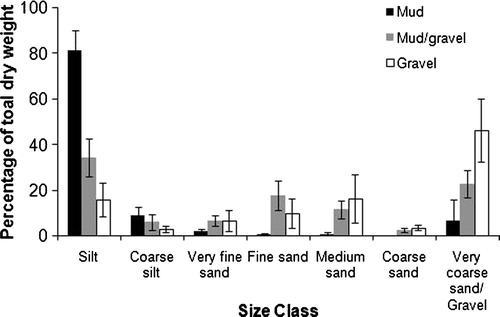
Fig. 2 Dry weight composition of sediment grain size class each of the three substrata (mud, mud/gravel and gravel).
Our data set was subject to two final, somewhat unorthodox classifications. The first of these was to determine whether any pattern in the distribution of species richness existed throughout the region, such as localised ‘hot spots’ of richness. To do this, we created a 7-point index of species richness from and unique to our data set, with relative measures of species richness ranging very low to very high (); for instance, should a sample from any site have three species in it, and the range in richness (number of species) throughout the region be 2–69 per sample, then such a site would be categorised as very low in richness (3 being 4.3% of 69). The second classification enables the spatial distribution of abundance (density) to be depicted, calculated in the same manner as species richness (); should a sample from any site have 21 individuals in it, and the range in numbers of individuals throughout the survey region for any given sample be 2–1572, then such a site would be categorised as very low in density (21 being ~0.1% of 1572). Both classifications may have limited application outside of the survey area, substratum (habitat) type or time during which this survey was undertaken.
Table 1 Classification of relative density and richness using a 7-point scale for all subtidal substratum types, eastern Waiheke Island.
Results
Within the 228 sea-bed samples off eastern Waiheke Island, 326 taxa were identified. Of these, 168 occurred beneath the mussel farm and 307 outside of it. Numbers and ranges of individuals, taxa and degree of apparent local endemism within our samples by substratum type are detailed in . Only 31.6% of taxa occurred in both muds and gravels, 40% occurred in both muds and muddy gravels, and 44.8% were common to both muddy gravels and gravels.
Table 2 Number of individuals and taxa per sample (0.0336 m2) in each substratum type.
Wet-sieved grain-size data for the three sediment samples collected from three representative sites within each substratum type are presented in and . Species accumulation curves for each of the substrata, and for them all combined are presented in . The relationship between species dominance, substratum type and species richness is depicted in .
Fig. 3 Species accumulation curves, all substrata.
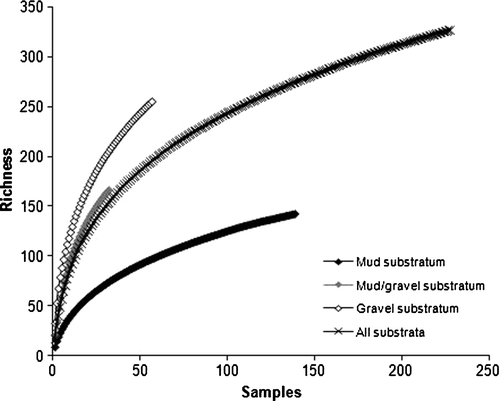
Fig. 4 The relationship between species dominance and richness, all substrata, eastern Waiheke Island.
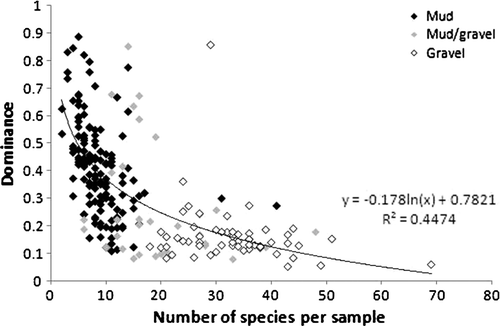
Table 3 Sediment grain size analyses by substratum type.
DIVERSE indices and community composition both differ significantly among substrata (ANOVA, P-value <0.05). Gravel samples, followed by mud/gravel samples, had the highest total number of individuals (n), Margalef's index of species richness (d), Pielou's evenness index (J’), Shannon–Wiener diversity index (H’), but lowest Simpson dominance index (λ); mud substrata had the lowest value for each DIVERSE index, with the exception of the Simpson dominance index (λ) (, ). Community composition was also significantly different among substratum types (R=0.575 − 0.951, P=0.1%), with mud/gravel sites having species assemblages intermediate between those of mud and gravel sites ().
Fig. 5 Multi-dimensional scaling plot of species assemblages within three substrata.
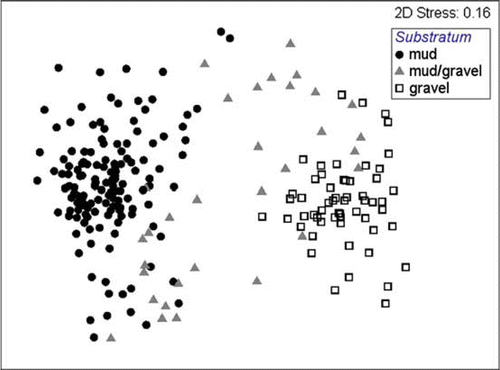
Table 4 DIVERSE indices by substratum type and depth (mean±SD), square-root transformed data.
Annelida (primarily Polychaeta) was the most species rich taxon in all substratum types; the number of polychaete taxa, in addition to those of Mollusca and Arthropoda, increased with increased sediment complexity, from muds to gravels (). Within muddy substrata, bivalves were proportionally most abundant, whereas polychaetes were proportionally (not necessarily absolutely) most abundant in mud/gravels and gravels (). The invasive bivalve Theora lubrica was the most regularly and abundantly occurring taxon in muddy samples throughout this region (). An ostracod (Ostracod sp. 2), Nemertea, three polychaete species (Prionospio sp., Heteromastus filiformis and Sthenelais sp.) were the most regularly and densely occurring taxa in mud/gravels (). Within gravels, four polychaetes (Heteromastus filiformis, Prionospio sp., spionid sp. 1 and Macroclymenella stewartensis) and one bivalve, Notocorbula zelandica, were the most regularly and abundantly occurring taxa ().
Table 5 Breakdown of average similarity in taxon richness by substratum, to Phylum, square-root transformed data.
Table 6 Breakdown of average similarity in taxon abundance by substratum to level of Class, and Phylum Nemertea (average similarity of samples in parentheses).
Table 7 SIMPER results for muds,average similarity: 39.09, square-root transformed data.
Table 8 SIMPER results for mud/gravels, average similarity: 22.09, square-root transformed data.
Table 9 SIMPER results for gravels, average similarity: 38.16, square-root transformed data.
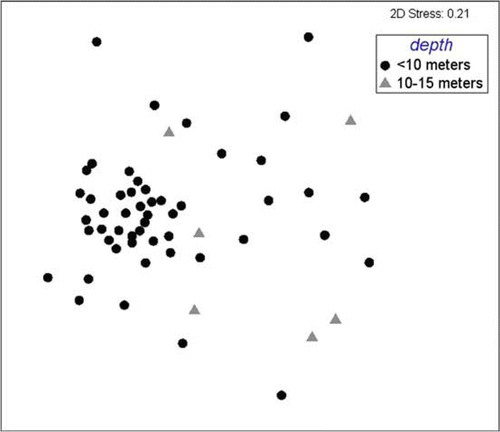
Taxa occurring in muds differed significantly by depth (R=0.046 − 0.223, P<0.5%; ), but no significant difference was apparent in any DIVERSE index. ANOSIM revealed assemblages of species from muddy gravel sites only differed significantly between sites more shallow than 10 m and those deeper than 15 m (R=0.315, P=0.3%; ), but again, DIVERSE indices were not significantly different between these depths. To the contrary, ANOSIM revealed species assemblages from gravel sites did not differ significantly between depths (), but one DIVERSE index, Margalef's index of species richness (d), was significantly different, being higher at depths 10–15 m than depths <10 m ().
Fig. 6 Multi-dimensional scaling plot of species assemblages within muds, grouped by depth (<10, 10–15, >15 m).
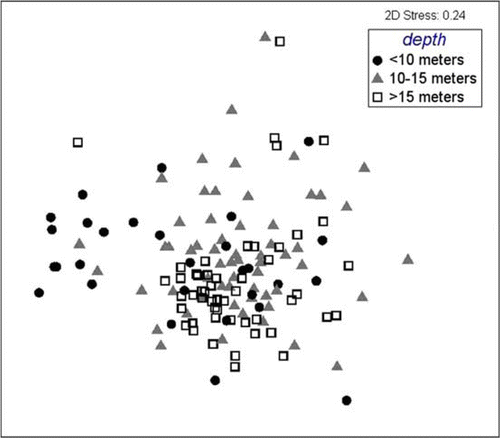
Fig. 7 Multi-dimensional scaling plot of species assemblages within mud/gravels, grouped by depth (<10, 10–15, >15 m).
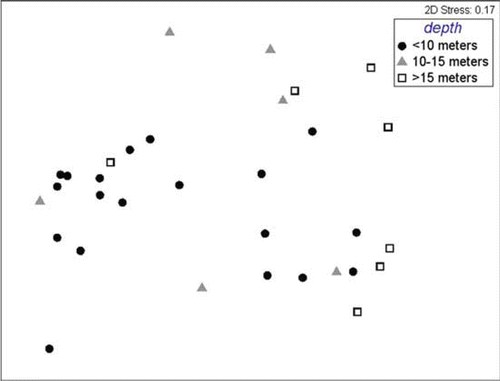
Fig. 8 Multi-dimensional scaling plot of species assemblages within gravels, grouped by depth (<10, 10–15 m).
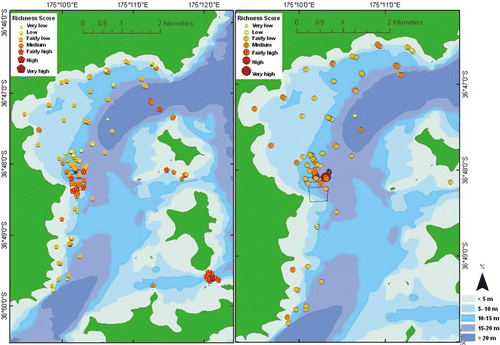
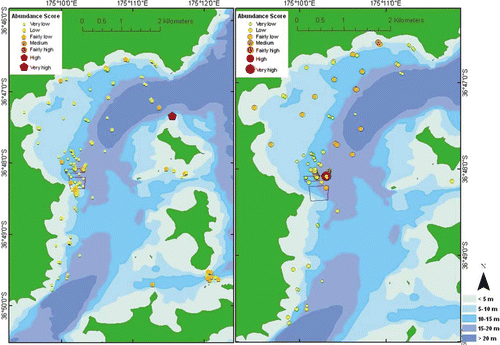
The most species-rich sites (relative to all sites throughout the survey region) prove to be those beneath and in the immediate vicinity of an existing mussel farm, in the deeper channels between Waiheke Island and Pakatoa Island, and between Rotoroa and Ponui Islands (, left). When limiting analysis to relative species richness within muds, the most species-rich sites are rather evenly distributed throughout the eastern Waiheke Island region, but many occur inside and within the immediate vicinity of the northern side of the existing mussel farm (, right) — an area and substratum type subject to more intensive sampling effort than elsewhere.
Fig. 9 Spatial distribution of relative species richness, all substrata (left), muds (right).
Those sites with the greatest density of individuals proved to be beneath and in the immediate vicinity of the existing mussel farm, in the deeper channels between Waiheke Island and Pakatoa Island, and Rotoroa and Ponui Island, and one site of relatively exceptional spionid polychaete density north of Pakatoa Island (, left). When limiting analysis to density within muds only, those sites with the greatest density of individuals are also rather evenly distributed throughout the eastern Waiheke Island region, but again many occur inside and within the immediate vicinity of the northern side of the existing mussel farm, at depths exceeding 20 m north and northeast of the mussel farm, and in the deeper parts of the channel between Waiheke and Pakatoa Islands (, right).
Fig. 10 Spatial distribution of relative species density, all substrata (left), muds (right).
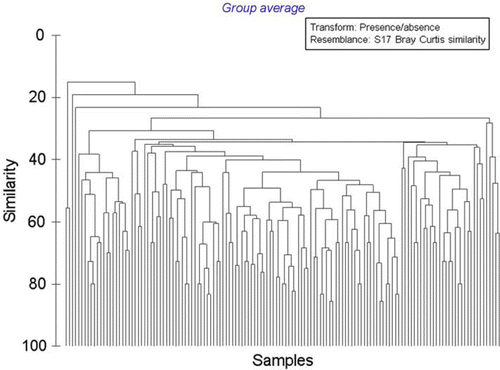
When analysis is limited to the presence or absence of taxa identified in all samples, two main clusters of species assemblages are apparent, muds (right) and gravels (left), with species assemblages occurring within muddy-gravels occurring throughout both (); the cluster using full quantitative data (not presented here) is even less clear. When limiting analysis to presence or absence of molluscan and echinoderm taxa, those two phyla largely used to characterise communities in the schemata depicted by Powell (Citation1937) and Hayward et al. (Citation1997), two main clusters of species assemblages of muds (right) and gravels (left) are again apparent, but those sites of a muddy-gravel nature similarly occur throughout both (). When analysis is limited to the most intensively sampled substratum type, muds, for which the species accumulation curve most closely approximates an asymptote (), it is especially obvious that recurring assemblages of species do not occur, in that no two sites share the same assemblage of taxa ().
Fig. 11 Dendrogram of similarity in faunal composition of all sites (n=228), all substratum types, eastern Waiheke Island, using presence/absence data.
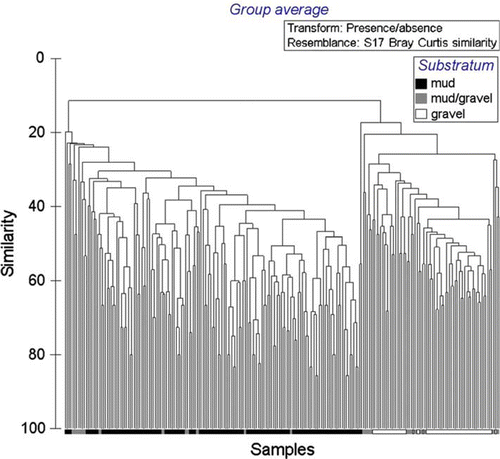
Fig. 12 Dendrogram of similarity in molluscan and echinoderm taxa (n=228), all sites, all substratum types, eastern Waiheke Island, using presence/absence data.
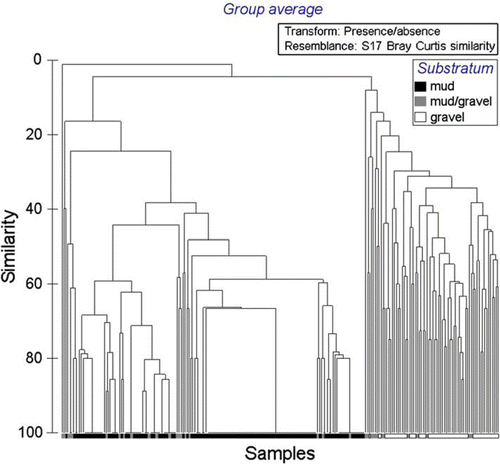
Fig. 13 Dendrogram of similarity in faunal composition of all sites, muddy substrata (n=139), eastern Waiheke Island, using presence/absence data.
Discussion
Potential changes in community structure
Benthic-invertebrate assemblages off eastern Waiheke Island were attributed to one of two formations by Powell (Citation1937): a Venericardia (now Purpurocardia) formation between Rotoroa Island and Ponui Island, and off the eastern side of Ponui Island (), and an Echinocardium formation throughout most of the rest of the region. Too few samples were collected at sites falling within Powell's Purpurocardia (as Venericardia) formation in this current study to fully compare/contrast with Powell's earlier formations, although our results show no recurring or widely distributed community structures occur within areas attributed to this formation/association, or that of his Echinocardium formation/association.
Only four of Powell's (1937) nine sea-bed stations off eastern Waiheke Island occur within the area re-surveyed herein; for only two of Powell's stations did he provide species inventories (Powell's J3, J4); we have resampled only one of these stations (J4). A further two sites surveyed by Powell, for which no species inventories were provided, were also resampled (Powell's J1, J6), both attributed to his Tawera+Purpurocardia formation between Rotoroa and Ponui Islands. On the basis of recent sampling, neither Tawera nor Purpurocardia characterise species assemblages within gravels in this region, although the small, thick-shelled bivalve, Notocorbula zelandica, a species recorded from more than 95% of gravel samples, does appear appropriate for this purpose ().
Excluding Powell's unidentified polychaetes, 32% (nine of 28 taxa, ) reported by him from two of four sea-bed stations within formations for which he furnished taxonomic inventories (Powell Citation1937: 370–384) were not re-identified during our sampling. Two of Powell's taxa were likely misidentified (Petrolisthes elongatus and Nectocarcinus antarcticus); Powell's ‘small, pink holothurian’ was probably one of Ocnus brevidentis or, more likely, Trochodota dendyi, both of which occur throughout this region, the latter being more common. One notable addition to the faunal inventory from this region is the establishment of the invasive bivalve taxon Theora lubrica; this species now characterises well-sorted muddy sediments off eastern Waiheke Island, in addition to those muddy sediments in Waitemata Harbour and Rangitoto Channel reported by Hayward et al. (Citation1997). Having been relatively recently recognised from New Zealand waters, first recorded in 1971 (Climo Citation1976), this species has become widespread throughout Waitemata Harbour and Hauraki Gulf; it was not recognised by Powell in his surveys seven decades earlier (Powell Citation1979: 451).
Table 10 Nomenclature of taxa reported by Powell (1937) and this study.
Species assemblages occurring within muds differ markedly from those of gravels, but those of muddy-gravels are somewhat transitional between the two ( and ). Muds, muddy gravels and gravelly substrata are also patchily distributed throughout the eastern Waiheke Island region (). Because of this, and because of the high level of dissimilarity in assemblage structure between the greatest majority of samples throughout this region, regardless of what transformation we subject our data to ( and ), we do not recognise recurring, spatially discrete assemblages of species akin to those of Powell (Citation1937) off eastern Waiheke Island. Accordingly, we make no attempt to depict schematically the distributions of any communities throughout this region. The reality is that too few sites were sampled by Powell (Citation1937) for him to have generalised sea-bed communities throughout this region, and the same may apply in our study (with a combined surface area of all grabs sampled being only 7.66 m2). An alternative, plausible argument (Gray Citation1981) is that discrete assemblages of species do not exist, as the distributions of species overlap, with one community grading into another; alternatively, the scale of our survey was too small, and the range of habitats too limited for any discrete community types to develop, although we consider this to be less likely given the intensity of our sampling, the range in substratum types experienced, and the relatively large size of the survey area.
Assemblage structure
Although recurring assemblages do not occur in the region, some relationships were apparent in the composition of assemblages by substratum type. The moderately to moderately well-sorted muds were characterised by bivalves, ostracods, polychaetes and amphipods; the very poorly sorted mud/gravel-dwelling in- and epifaunal species assemblages were characterised by polychaetes, ostracods, amphipods, bivalves and ophiuroids; and the poorly sorted gravel-dwelling in- and epifaunal species assemblages were characterised by polychaetes, bivalves, pagurid crabs, gastropods, ostracods, ophiuroids and nemertean worms.
In this study, the higher numbers of taxa and diversity in gravel samples could be related to the poorly sorted nature of the substratum, with such structurally heterogeneous substrata providing more niche space, therefore elevating diversity (Gray Citation1981); conversely, the well-sorted, structurally homogeneous muds had the least diverse communities ( and ). However, this relationship is not without exception; for example, Ellingsen & Gray (Citation2002) discerned no relationship between species richness and sediment properties (sorting coefficient and percentage of silt) on the Norwegian continental shelf. Regardless of whether the individual substrata are classified as well sorted, moderately well, poorly or very poorly sorted, when the incidence of gravel is used as a proxy for increased structure or complexity, it is apparent that elevated diversities and densities of benthic invertebrate taxa are encountered (). These findings are consistent with those of Dewas (Citation2008) for sea-bed communities off Otata Island, Hauraki Gulf, where the density and richness of benthic invertebrates in adjoining types of sea-bed, complex valves of the bivalve Tucetona laticostata interspersed with rhodoliths, and less-structured and extensively fragmented coarse sands, were greater in the former.
Of the 326 taxa recorded from the 228 grab samples throughout this region, 142 taxa were identified from 139 muddy sites, 166 taxa from 32 muddy gravel sites, and 255 taxa from 57 gravel sites; less than 50% of the taxa were common to any two of these substratum types, rendering the assemblages of each substratum unique. In fact, no two grab samples had the exact same complement of taxa either (), effectively rendering each surveyed site unique. None of the species accumulation curves prepared for individual substrata reached an asymptote, although that for muds most closely did; it is apparent that many additional taxa occur within each substratum type in this region, particularly within the coarser substrata, and on the basis of monitoring exercises throughout this region, seasonally also (Wong Citation2009). Although none of these accumulation curves reached an asymptote, this could be typical of marine benthic-community studies (Gray Citation2002). A major factor contributing to this in our study is the relative rarity of species throughout the survey region, with almost one third of taxa (109 taxa) recorded from a single site only.
As species richness increased, concomitant with an increase in substratum complexity, there also was a tendency for dominance of taxa to decrease (, Tables ). This relationship is more typical of terrestrial systems (e.g. Odum Citation1971; Hill Citation1973) than marine systems, with Birch (Citation1981) recognising an inverse relationship between dominance and species richness in the marine environment, and Gray (2002) recognising no relationship between these two at all.
During the survey period, February 2008, little variation was apparent in the distribution of species richness throughout the eastern Waiheke Island region, but density was more variable. Muds and mud/gravel substrata had minimum and maximum densities of individuals of 119 m−2 and 17,440 m−2, and 238 m−2 and 6905 m−2 respectively (with mean densities of 1458 m−2 and 2351 m−2, respectively); the greatest density of individuals throughout this region was encountered in gravels, with minimum and maximum densities of 1339 m−2and 46,786 m−2, of mean density 5512 m−2. The value of the density schema used herein is that it enables a comparison of the relative densities of individuals in sea-bed samples throughout Hauraki Gulf, at least for those limited locations, dates and depths that have been surveyed in a quantitative manner. Because of non-standardised sampling volumes or surface areas, available density data have had to be standardised to number of individuals per m2 to enable some comparison to be made; we do realise the limitations in this approach, especially given the total sea-bed area sampled in this study was 7.66 m2 only. To date, those areas throughout Hauraki Gulf with the greatest densities of individuals, in decreasing order, occur off Otata Island in Tucetona/rhodolith-based shell gravels (142,385 m−2; Dewas Citation2008); eastern Waiheke Island in shell gravels (with limited Tucetona and no rhodoliths, 46,786 m−2; data reported herein); muds off eastern Waiheke Island (17,440 m−2; herein); muddy gravels off eastern Waiheke Island (6905 m−2; herein); muds off eastern Motutapu Island (maximum densities 5832 m−2; Dewas Citation2008); undefined substrata (potentially all of muds, mud/gravels and gravels) in Rangitoto Channel (maximum density 4440 m−2; Roberts Citation1990); muds off eastern Motutapu Island (mean density 1797 m−2; Dewas Citation2008); and muds proximal to the mussel farm in the Firth of Thames (mean density 115.5 m−2 outside and 84 m−2 inside a mussel farm, de Jong Citation1994).
The density scale proposed for off eastern Waiheke Island obviously will vary spatially, and temporally given both Dewas (Citation2008) and Wong (Citation2009) report greatest densities of individuals in samples surveyed during mid-winter. Accordingly, this classification must be used with some caution when extrapolating to other areas throughout Hauraki Gulf, and obviously would not apply for the sea-bed off Otata Island, the most dense and species rich area thus-far recognised in Hauraki Gulf. Other quantitative studies throughout the region are lacking, or data are not presented in a manner enabling direct comparison.
Those sites within 80 m of the boundary of the existing mussel farm, and within the farm itself had the greatest densities of individuals per sample, and those to the north of the farm, and northeast and south of the farm in deeper waters, had the least numbers of individuals per sample. On the basis of species richness and density of benthic invertebrates throughout the region ( and ), the mussel farm would not appear to be having any demonstrable negative or large-scale effect on either. The results of temporal monitoring of sea-bed communities beneath and at progressively increasing distances from this mussel farm will be reported separately.
The lack of knowledge on the distribution and composition of sea-bed communities throughout Hauraki Gulf is lamentable. Without this information, no party is in any position to make informed decisions on the relative merits of any area for conservation, development, or, and topical at present, harbour spoil disposal. Despite this, each activity is routinely called for or undertaken in Hauraki Gulf. There is a compelling case for ongoing surveys to determine present-day patterns in the geographic and temporal distribution and density of species, augmented with more systematic research to identify many currently problematic taxa. This information is largely absent and urgently required by agencies to ensure sustainable management of Hauraki Gulf marine resources (HGF Citation2008).
Acknowledgements
We wish to acknowledge all staff and students in the Earth and Oceanic Sciences Research Institute, and School of Applied Sciences at Auckland University of Technology that assisted in data collection. Special thanks are due to Emma Beatson for skippering the AUT vessel Taniwha, and Drs Lindsey White, AUT, Martin Cryer (New Zealand Ministry of Fisheries) and one anonymous referee for their constructive comments on an earlier draft of this manuscript. Funding for this study was provided by the Earth & Oceanic Sciences Research Institute, and School of Applied Sciences, Auckland University of Technology.
Related Research Data
References
- Birch , DW . 1981 . Dominance in marine ecosystems . The American Naturalist , 118 : 262 – 274 .
- Clarke , KR and Green , RH . 1988 . Statistical design and analysis for a ‘biological effects’ study . Marine Ecology Progress Series , 46 : 213 – 226 .
- Clarke , KR and Gorley , RN . 2006 . PRIMER v6: User manual/tutorial , Plymouth : PRIMER-E Ltd .
- Clarke KR Warwick RM 1994 Change in marine communities: an approach to statistical analysis and interpretation Plymouth Marine Laboratory, Natural Environment Research Council
- Climo , FM . 1976 . The occurrence of Theora (Endopleura) lubrica Gould, 1861 (Mollusca: Bivalvia: Semelidae) in New Zealand . Auckland Institute and Museum Conchology Section Bulletin , 1 : 11 – 16 .
- Dewas SEA 2008 Benthic-invertebrate diversity of Tucetona laticostata (Mollusca: Bivalvia) biogenic substrata in Hauraki Gulf Unpublished M.App.Sc. thesis, Auckland University of Technology
- de Jong RJ 1994 The effects of mussel farming on the benthic environment Unpublished M.Sc. thesis, University of Auckland
- Ellingsen , KE and Gray , JS . 2002 . Spatial patterns of benthic diversity—is there a latitudinal gradient along the Norwegian continental shelf? . Journal of Animal Ecology , 71 : 373 – 389 .
- Gray , JS . 1981 . The ecology of marine sediments. An introduction to the structure and function of benthic communities , Cambridge : Cambridge University Press .
- Gray , JS . 2002 . Species richness of marine soft sediments . Marine Ecology Progress Series , 224 : 285 – 297 .
- Hayward , BW , Stephenson , AB , Morley , M , Riley , JL and Grenfell , HG . 1997 . Faunal changes in Waitemata Harbour sediments, 1930s–1990s . Journal of the Royal Society of New Zealand , 27 : 1 – 20 .
- HGF 2008 Tikapa Moana — Hauraki Gulf state of the environment report Hauraki Gulf Forum Auckland Regional Council
- Hill , MO . 1973 . Diversity and evenness: a unifying notation and its consequences . Ecology , 54 : 427 – 432 .
- Odum , EP . 1971 . Fundamentals of ecology , Saunders .
- Powell , AWB . 1937 . Animal communities of the sea-bottom in Auckland and Manukau Harbours . Transactions and Proceedings of the Royal Society of New Zealand , 66 : 354 – 401 .
- Powell , AWB . 1979 . New Zealand Mollusca: marine, land and freshwater shells , Collins .
- Roberts RD 1990 Impact of dredge spoil at sea Unpublished M.Sc. thesis, University of Auckland
- Wong KLC 2009 The effects of green shelled mussel mariculture on benthic communities in Hauraki Gulf Unpublished M.App.Sc. thesis, Auckland University of Technology