Abstract
Few studies have compared hyperbenthic communities among estuaries, in either New Zealand or elsewhere. This study presents the results of a single late summer survey of the lower reaches of 26 estuarine systems along the Otago coastline from the Clutha River to Oamaru, comparing hyperbenthic communities among both permanently open and intermittently closed estuaries. Intermittently closed estuaries are periodically closed from the ocean because of the formation of a berm across the mouth. A total of 33 taxa were captured with mysids and amphipods, the most widespread hyperbenthic taxa, being common in both permanently open and intermittently closed Otago estuaries. The mysid, Tenagomysis novae-zealandiae, and amphipod, Paracalliope fluviatilis, were widespread, found in 74% and 68% of all estuaries sampled, respectively. In contrast, most other taxa were present in relatively few estuaries, with 17 species recorded from only 10% of estuaries. There was a significant difference in the community composition among permanently open and intermittently closed estuaries. The results highlight the potential sensitivity of New Zealand estuarine communities to management regimes that may alter their open/closed status.
Introduction
While a large number of ecological studies have been conducted in estuaries, studies of single large permanently open estuaries located close to centres of human habitation are over-represented in the literature (Edgar et al. Citation1999; Teske & Wooldridge Citation2001; Hirst Citation2004). However, a large proportion of estuarine systems are very small. For example, along the Australian NSW coastline, the majority of the 950 estuarine systems are small and intermittently closed from the sea (Roy et al. Citation2001), while in South Africa 70% of estuaries are open only temporarily (Nozais et al. Citation2001; Perissinotto et al. Citation2002; Froneman Citation2004). Although large river estuaries are relatively common in New Zealand, there is also an abundance of small and intermittently closed estuaries that have received relatively limited attention.
Consequently, few studies worldwide have examined the distribution and spatial variation of biota among different types of estuaries. For example, comparisons of estuarine systems completed to date suggest that there can be significant differences among hyperbenthic communities in different estuary types (e.g. Mees et al. Citation1995; Teske & Wooldridge Citation2001; Desmond et al. Citation2002; Dye & Barros Citation2005a,Citationb). Whether an estuary is permanently open or intermittently closed, in particular, can cause marked physico-chemical variation among estuaries including differences in salinity, temperature, inundation, nutrients and production (Kozlowski-Suzuki & Bozelli Citation2004; Thomas et al. Citation2005). Consequently, significant differences in the hyperbenthic communities present in permanently open or intermittently closed systems have been observed; for example in Australia (e.g. Hirst Citation2004; Dye & Barros Citation2005a,Citationb) and South Africa (e.g. Teske & Wooldridge Citation2001). It is presumed that this pattern is also likely to occur in New Zealand.
Worldwide, estuarine hyperbenthic communities have also received only limited study (Mees et al. Citation1995). These species occupy a critical niche, providing linkages between the planktonic and detrital food webs and larger secondary consumers (e.g. Webb Citation1973; Fockedey & Mees Citation1999; Cabral & Ohmert Citation2001; Maes et al. Citation2003). Mysid shrimps are typically the most abundant hyperbenthos in temperate hyperbenthic estuarine communities, dominating the estuarine fauna in many regions (Azeiteiro & Marques Citation1999; Roast et al. Citation1999; San Vicente & Sorbe Citation1999; Mouny et al. Citation2000). The distribution of mysids is relatively unknown in New Zealand (Chapman & Lewis Citation1976; Jones et al. Citation1989), but it is likely that they are abundant in many estuaries.
There is growing concern in New Zealand over the impact of both deteriorating water quality and climate change on our aquatic ecosystems (Larned et al. Citation2004; McDowell and Wilcock Citation2008). Estuaries are vital to our understanding of these impacts, as they potentially integrate these impacts from over the whole catchment and often transport pollution to the ocean because of their role as the conduit of freshwater run-off (Scharler & Baird Citation2003; Davies & Eyre Citation2005). In particular, intermittently closed estuaries are thought to be especially vulnerable because of longer residence times and limited opportunities for flushing (Haines et al. Citation2006; Schallenberg et al. Citation2010). Hence it is important to gain a baseline understanding of estuarine taxa and how estuarine communities vary among systems.
The aim of this paper is to (1) provide a late-summer spatial comparison of hyperbenthos composition among representative estuaries along the Otago coastline; and (2) focus on mysid distribution and variation in relative abundance between permanently open and intermittently closed estuarine systems. It is hypothesised that the presence of a barrier across the estuary mouth will create physico-chemical differences that will lead to distinct hyperbenthic communities among these two estuarine types.
Methods
Study sites
Twenty-six estuaries were chosen using 1:50,000 topographic maps in a stratified design in order to obtain representatives from a wide variety of estuary types and sizes over a large spatial scale along the Otago coastline (refer to for name and location of estuaries sampled). Estuaries were sampled along approximately 250 km of the south east coast of the South Island of New Zealand from the Clutha River (46.33775°N, 169.80696°E) in the south to Awamoa Creek (45.14268°N, 170.93506°E) near Oamaru in the north. All the types of estuarine systems in Otago are represented in the study including large river estuaries (e.g. Clutha River), drowned river estuaries (e.g. Taieri River), coastal lagoons (e.g. Reids Stream) and coastal inlets (e.g. Hoopers Inlet). Sixteen estuaries sampled were known to close frequently or were closed by sand bars at the time of sampling and were thus identified as intermittently closed systems.
Table 1 Site number, estuary name, location of sample and typology as sampled on 17–20 February 2004.
Sampling protocol
Sampling was conducted at night on 17–20 February 2004 during a period of settled weather with no significant rainfall. Night-time sampling is known to maximise both catch per unit effort and the number of species caught, and is important in order to avoid any crepuscular bias in invertebrate activity (Guest et al. Citation2003). While such a short sampling period offers only a snapshot of the distribution and variation in estuarine fauna, it was conducted in order to avoid seasonal or significant climatic variation confounding patterns of spatial variation among estuaries. Furthermore, as the analyses treated each estuary as an individual sampling unit, an attempt was made to maximise the number of estuaries sampled over as short a time span as possible. All intermittently closed estuaries apart from Awamoa were closed during sampling.
The following environmental variables were recorded at each estuary: connection to the sea (open versus closed), salinity, temperature, latitude and longitude. In order to maximise the number of estuaries sampled and to increase the spatial extent of the study, only one hyperbenthos sample was collected in each estuary. A qualitative sample was taken at each estuary using a 240 mm width by 200 mm height handheld sweep net with 500-µm mesh. Each sample was collected over 2 min with the net swept in front of the sampler and constantly moving over new area, and covered approximately 15 m2. Sampling was limited to locations by the estuary mouth, allowing for a standardisation of habitat type across estuaries with areas only of soft sand/mud substrates sampled and in water greater than 20 cm deep. Only the water column just above the benthos was sampled and areas of fast-flowing water were avoided. After collection, all samples were preserved in 70% ethanol.
In the laboratory, the presence/absence of all invertebrate taxa collected was recorded for each sample. Because of their dominance, the number of each mysid species by sample was also counted to allow comparison of relative abundance across estuaries. PRIMER 5 was used to investigate patterns in the data. Two Bray–Curtis similarity matrices were created with standardised presence/absence data. One measured the similarities in taxa present among estuaries. The other measured the similarities among the taxa in which estuary they were present. Multidimensional scaling (MDS) with 1000 restarts was used to compare assemblage structure among estuaries. MDS was also used to compare the differences in estuary occupation among taxa. Differences among communities in temporarily open/closed and permanently open estuarine systems, and between high and low salinity communities, were examined in detail. ANOSIM (1000 permutations) comparing estuarine communities was conducted to test the a priori hypothesis that there was a difference between temporarily open/closed and permanently open estuaries.
Results
Physical and chemical variation
There was no apparent relationship between geographic location (longitude and latitude) and type of estuarine system, with the 16 intermittently closed systems evenly dispersed amongst the 10 permanently open systems (). Similarly, there were no detectable relationships between geographic location and the suite of environmental variables recorded. Salinity was significantly different between permanently open and intermittently closed systems (two-sample t-test, t-stat = 4.20, P<0.01, df = 25) with mean salinity of 23.9 PSU in open estuaries and 7.8 PSU in intermittently closed systems. No significant difference in mean temperature was observed among permanently and intermittently closed systems.
Community structure
Thirty-three taxa were collected across the sampled estuaries. However, only 16 taxa were recorded in 10% or more of the estuaries sampled (Table 2). The total number of taxa found in permanently open estuaries was 21, while 20 taxa were recorded in intermittently closed systems. Mean taxa richness was equivalent with 5.3±0.6SE and 5.8±0.7SE taxa for open and intermittently closed systems respectively (analysis of variance; ANOVA, P>0.05, F = 0.27, df = 1). Mysid and amphipod taxa were widespread and accounted for 22.5% and 25.5% of taxa recorded, respectively. The two most widespread species were Tenagomysis novae-zealandiae and Paracalliope fluviatilis, being present in 74% and 68% of the estuarine systems sampled respectively ().
Table 2 Taxa list with percentage prevalence in 26 Otago estuaries, and presence of taxa in permanently open, intermittently closed or both estuarine systems.
SIMPER analysis () indicates that both T. novae-zealandiae and P. fluviatilis were widespread in both open and intermittently closed estuaries and contributed over 50% to the similarity across each estuary type. The amphipod Paracorophium excavatum was important in open systems being present in 64% of these systems compared with 31% of closed systems. Conversely, chironomid taxa were important in closed systems, being present in 63% of closed systems compared with 25% of open systems. These taxa collectively explained 75% of the similarity among open or among closed systems. The remaining taxa contributed to no more than 10% each of the similarity among permanently open or intermittently closed systems.
Table 3 Output of SIMPER analysis comparing the contributions of species similarities among (A) permanently open estuaries and the similarities among (B) intermittently closed estuaries.
MDS ordination of taxa presence/absence arranged estuaries by estuarine type (permanently open/intermittently closed) (2D Stress 0.14) (A). Comparison of community composition in permanently open and intermittently closed estuaries by ANOSIM indicated that communities in these types of system were significantly different from each other (ANOSIM; Global R=0.249, P<0.01). Separation of estuarine types was less evident when ordered by high and low salinity (B). Along MDS axis 2, three significantly different groups of taxa could be distinguished determined by the presence of only T. novae-zealandiae or only P. fluviatilis or both species together (C).
Figure 1 Multidimensional scaling (MDS) ordination of the similarity among estuaries sampled based on presence/absence of taxa. A, Key indicates whether estuary is classified as permanently open or intermittent closed. B, Key indicates whether estuary is classified as high (>10 ppt) or low (<10 ppt) salinity. C, Key indicates estuaries classified by the presence of Tenagomysis novae-zealandiae or Paracalliope fluviatilis or both.
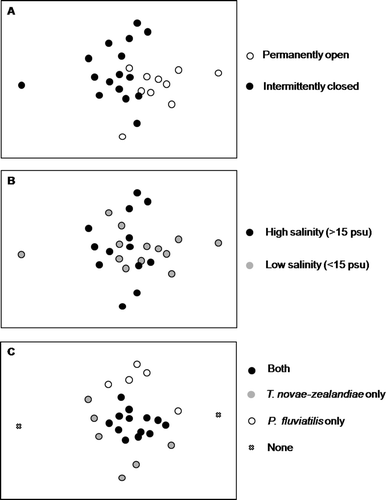
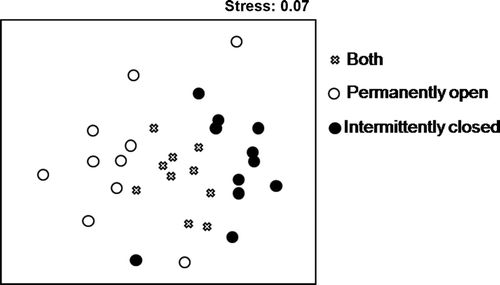
Ordination by MDS of similarity among species taxa distribution also indicated separation of estuary communities by their permanently open or intermittently closed status (2D Stress 0.07). Three significantly different groups of taxa could be distinguished in , with species present exclusively in intermittently closed estuaries to the right side of MDS axis 1 and species present exclusively in open estuaries to the left side of MDS axis 1, while species present in both estuary types occupied the middle of MDS axis 1. These three groups are significantly different from each other (ANOSIM Global R=0.33, P<0.01). No relationships between taxa and habitat variables were apparent along MDS axis 2. Whether a taxa is present exclusively in permanently open, intermittently closed or in both types of systems is indicated in .
Figure 2 Multidimensional scaling (MDS) ordination of similarity among all taxa sampled based on presence or absence in estuaries. Key indicates whether taxa is present in: both permanently open and intermittently closed estuaries, exclusively in intermittent closed estuaries or exclusively in open estuaries
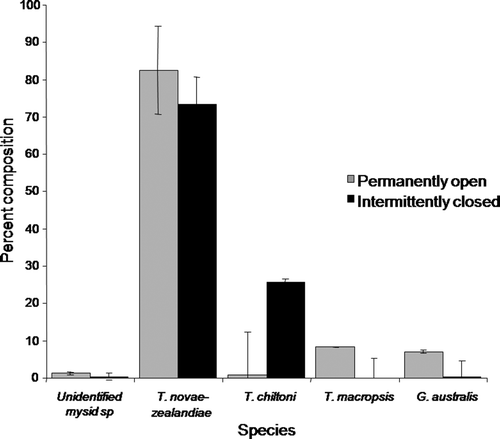
Numerically, T. novae-zealandiae was the dominant mysid in both systems, comprising about 75% of mysids in temporarily open/closed systems and 85% of mysid numbers in open systems (). T. chiltoni was relatively less abundant or absent in open systems, whilst T. macropsis and Gastrosaccus australis were relatively more abundant in open systems.
Figure 3 The mean relative abundance (±SE) of mysid species recorded in open and temporarily open/closed estuaries along the Otago coastline, 17–20 February 2004, n=28.
Discussion
Species distribution and richness
This study provides a late summer baseline survey of hyperbenthic species that inhabit the lower reaches of Eastern Otago estuaries. The broad spatial extent of this study enabled sampling from a wide range of estuarine types, but precluded examination of spatial or temporal variation within estuaries. Despite the limited extent of sampling within estuaries, some clear patterns were evident in Otago hyperbenthic estuarine communities that inhabit the mouth regions of estuaries. Mysids (G. australis, Tenagomysis chiltoni, Tenagomysis macropsis and T. novae-zealandiae) and amphipods (Melita awa, Paracorophium sp. and P. fluviatilis) were widespread among estuaries. Geographically, mysids were the most widespread group of hyperbenthic fauna in the Otago estuaries with only three estuaries having no mysids recorded. This pattern of mysid and amphipod dominance appears to be a common feature of hyperbenthic communities in temperate estuaries worldwide (e.g. Mees et al. Citation1995; San Vicente & Sorbe Citation1999; Mouny et al. Citation2000; Hirst Citation2004).
Relatively few taxa sampled were widely distributed across the sampled estuaries, with only two species, P. fluviatilis and T. novae-zealandiae, being recorded from >50% of estuaries sampled. On average, each taxon was recorded from only 15% of sites sampled. Hyperbenthic estuarine communities dominated by the presence of rare taxa was also reported by Hirst (Citation2004) who found that nearly 50% of species collected in south east Australian estuaries were found at only one site. Drake et al. Citation2002 reported 134 fish and hyperbenthic species in the Guadalquivir Estuary (Spain) and concluded that over half (72) were present at only a few sites. Whilst it is unlikely that most hyperbenthic taxa are restricted to only a few estuaries in Otago, it is likely that as estuaries harbour temporally dynamic communities, only a few estuaries along the same coastline will harbour similar communities at any one time.
Open versus intermittently closed estuaries
Over half of the estuaries studied were closed at the time of sampling highlighting the significance of this estuarine type along the Otago coastline. As hypothesised a priori, a significant difference in hyperbenthic communities between open and intermittently closed estuaries was observed, with significant differences in overall community composition and in the relative abundance of mysid species. Species more typically associated with marine environments, such as T. macropsis (Chapman & Lewis Citation1976; Sutherland & Closs Citation2001a), were recorded more frequently from permanently open estuaries, whereas taxa generally associated with freshwater habitats, particularly various insects and T. chiltoni (Chapman & Lewis Citation1976), were recorded more frequently in the intermittently closed estuarine systems. Mysids were recorded from all estuarine types; four species were present in open estuaries, whereas only T. chiltoni and T. novae-zealandiae were recorded from intermittently closed estuaries. The occurrence of only these two species in intermittently closed systems is consistent with records of mysid abundance collected over one annual cycle in the intermittently closed Kaikorai Lagoon, where only T. chiltoni and T. novae-zealandiae were abundant and observed to reproduce during 2004 (Lill et al. Citation2010).
The greater separation of the sampled estuaries into groups based on estuarine type (permanently open versus intermittently closed), rather than by salinity, indicate community differences are most likely driven by a number of inter-related factors related to estuarine closure, rather than a single factor (Hirst Citation2004; Dye & Barros Citation2005a,Citationb). Communities in intermittently closed estuaries are influenced by isolation from marine environments and the wide fluctuations in various physico-chemical factors associated with the occasional and irregular intrusion of seawater, compared with the cyclical tides that occur in permanently open estuaries (Edgar et al. Citation1999; Teske & Wooldridge Citation2001; Kibirige & Perissinotto 2003b; Kozlowski-Suzuki & Bozelli Citation2004; Dye & Barros Citation2005a,Citationb). The degree of isolation because of barriers across the mouth of an estuarine system has consequences for its trophic and community structures (Dye & Barros Citation2005a,Citationb; Thomas et al. Citation2005), as well as the size and genetic make-up of populations of species present (Carr et al. Citation2003). In the permanently open Taieri River estuary, a system sampled in this study, T. macropsis were recorded drifting into the estuary on incoming tides (Sutherland & Closs Citation2001a) and juveniles of fish with marine adult life-history stages have also been recorded (Sutherland & Closs Citation2001b), indicating that tidal exchange is important for maintaining the presence of certain species. Tidal conditions in intertidal creeks have been shown to be a major factor structuring hyperbenthic communities, especially mysid and amphipod assemblages (Hampel et al. Citation2003). Also, freshwater inflow is limited in intermittently closed estuaries and the rate of freshwater inflow has a major influence on community structure and diversity (Teske & Wooldridge Citation2001; Gillanders & Kingsford Citation2002).
The difference between permanently open and intermittently closed estuarine communities observed in this study is consistent with studies conducted overseas. Desmond et al. (Citation2002) conducted a long-term study of three Californian estuaries and suggested that the closure of estuaries might play an important role in estuarine community structure including increased invertebrate density and decreased fish numbers. Edgar et al. (Citation1999) reported that the presence of a bar across the estuary is the factor most strongly correlated with community structure in Tasmanian estuaries. Kibirige & Perissinotto (2006) found that the opening of a lagoon increased the zooplankton diversity and drastically altered the food web structure. Hirst (Citation2004) compared 28 estuaries in south-eastern Australia and found a significant difference in macrofaunal assemblages between open and closed estuaries. Teske & Wooldridge (Citation2001) describe high hyperbenthic density in intermittently closed estuaries while species diversity is highest in permanently open estuaries in thirteen estuaries in South Africa. The results of this study provide further support for the view that altering the open/closed status of an estuary, for example, by altering upstream flow regimes or artificially opening natural sand barriers, will have significant impacts on estuarine community structure and composition.
Acknowledgements
Many thanks go to the staff in the Department of Zoology for their support throughout the study. Candida Savage and Aparna Lal provided additional advice and comments on early drafts of the manuscript. Thanks also to the two anonymous reviewers who reviewed this paper.
References
- Azeiteiro , UMM and Marques , JC . 1999 . Temporal and spatial structure in the suprabenthic community of a shallow estuary (western Portugal: Mondego river estuary) . Acta Oecologica–International Journal of Ecology , 20 : 333 – 342 .
- Cabral , HN and Ohmert , B . 2001 . Diet of juvenile meagre, Argyrosomus regius, within the Tagus estuary . Cahiers de Biologie Marine , 42 : 289 – 293 .
- Carr , MH , Neigel , JE , Estes , JA , Andelman , S , Warner , RR and Largier , JL . 2003 . Comparing marine and terrestrial ecosystems: Implications for the design of coastal marine reserves . Ecological Applications , 13 : S90 – S107 .
- Chapman , MA and Lewis , MH . 1976 . An introduction to the freshwater Crustacea of New Zealand , Auckland : William Collins Limited .
- Davies , PL and Eyre , BD . 2005 . Estuarine modification of nutrient and sediment exports to the Great Barrier Reef Marine Park from the Daintree and Annan River catchments . Marine Pollution Bulletin , 51 : 174 – 185 .
- Desmond , JS , Deutschman , DH and Zedler , JB . 2002 . Spatial and temporal variation in estuarine fish and invertebrate assemblages: analysis of an 11-year data set . Estuaries , 25 : 552 – 569 .
- Drake , P , Arias , AM , Baldo , F , Cuesta , JA , Rodriguez , A , Silva-Garcia , A , Sobrino , I , Garcia-Gonzalez , D and Fernandez-Delgado , C. 2002 . Spatial and temporal variation of the nekton and hyperbenthos from a temperate European estuary with regulated freshwater inflow . Estuaries , 25 : 451 – 468 .
- Dye , AH and Barros , F . 2005a . Spatial patterns of macrofaunal assemblages in intermittently closed/open coastal lakes in New South Wales, Australia . Estuarine, Coastal and Shelf Science , 64 : 357 – 371 .
- Dye , AH and Barros , F . 2005b . Spatial patterns in meiobenthic assemblages in intermittently open/closed coastal lakes in New South Wales, Australia . Estuarine Coastal and Shelf Science , 62 : 575 – 593 .
- Edgar , GJ , Barrett , NS and Last , PR . 1999 . The distribution of macroinvertebrates and fishes in Tasmanian estuaries . Journal of Biogeography , 26 : 1169 – 1189 .
- Fockedey , N and Mees , J . 1999 . Feeding of the hyperbenthic mysid Neomysis integer in the maximum turbidity zone of the Elbe, Westerschelde and Gironde estuaries . Journal of Marine Systems , 22 : 207 – 228 .
- Froneman , PW . 2004 . Food web dynamics in a temperate temporarily open/closed estuary (South Africa) . Estuarine Coastal and Shelf Science , 59 : 87 – 95 .
- Gillanders , BM and Kingsford , MJ . 2002 . Impact of changes in flow of freshwater on estuarine and open coastal habitats and the associated organisms . Oceanography and Marine Biology , 40 : 233 – 309 .
- Guest , MA , Connolly , RM and Loneragan , NR . 2003 . Seine nets and beam trawls compared by day and night for sampling fish and crustaceans in shallow seagrass habitat . Fisheries Research , 64 : 185 – 196 .
- Haines , PE , Tomlinson , RB and Thom , BG . 2006 . Morphometric assessment of intermittently open/closed coastal lagoons in New South Wales, Australia . Estuarine, Coastal and Shelf Science , 67 : 321 – 332 .
- Hampel , H , Cattrijsse , A and Vincx , M . 2003 . Tidal, diel and semi-lunar changes in the faunal assemblage of an intertidal salt marsh creek . Estuarine Coastal and Shelf Science , 56 : 795 – 805 .
- Hirst , AJ . 2004 . Broad-scale environmental gradients among estuarine benthic macrofaunal assemblages of south-eastern Australia: implications for monitoring estuaries . Marine and Freshwater Research , 55 : 79 – 92 .
- Jones , MB , Greenwood , JG and Greenwood , J . 1989 . Distribution, body size, and brood characteristics of 4 species of mysids (Crustacea, Peracarida) in the Avon-Heathcote Estuary, New Zealand . New Zealand Journal of Marine and Freshwater Research , 23 : 195 – 199 .
- Kibirige , I and Perissinotto , R . 2003a . In situ feeding rates and grazing impact of zooplankton in a South African temporarily open estuary . Marine Biology , 142 : 357 – 367 .
- Kibirige , I and Perissinotto , R . 2003b . The zooplankton community of the Mpenjati Estuary, a South African temporarily open/closed system . Estuarine Coastal and Shelf Science , 58 : 727 – 741 .
- Kozlowsky-Suzuki , B and Bozelli , RL . 2004 . Resilience of a zooplankton community subjected to marine intrusion in a tropical coastal lagoon . Hydrobiologia , 522 : 165 – 177 .
- Larned , ST , Scarsbrook , MR , Snelder , TH , Norton , NJ and Biggs , BJF . 2004 . Water quality in low-elevation streams and rivers of New Zealand: recent state and trends in contrasting land-cover classes . New Zealand Journal of Marine and Freshwater Research , 38 : 347 – 366 .
- Lill , AWT , Lal , A and Closs , GP . 2010 . Life history and reproduction of two abundant mysids (Mysidacea: Mysidae) in an intermittently open New Zealand estuary . Marine and Freshwater Research , 61 : 1 – 9 .
- McDowell , RW and Wilcock , RJ . 2008 . Water quality and the effects of different pastoral animals . New Zealand Veterinary Journal , 56 : 289 – 296 .
- Maes , JL , de Brabandere , L , Ollevier , F and Mees , J . 2003 . The diet and consumption of dominant fish species in the upper Scheldt estuary, Belgium . Journal of the Marine Biological Association of the United Kingdom , 83 : 603 – 612 .
- Mees , J , Fockedey , N and Hamerlynck , O . 1995 . Comparative-study of the hyperbenthos of 3 European estuaries . Hydrobiologia , 311 : 153 – 174 .
- Mouny , P , Dauvin , JC and Zouhiri , S . 2000 . Benthic boundary layer fauna from the Seine Estuary (eastern English Channel France): spatial distribution and seasonal changes . Journal of the Marine Biological Association of the United Kingdom , 80 : 959 – 968 .
- Nozais , C , Perissinotto , R and Mundree , S . 2001 . Annual cycle of microalgal biomass in a South African temporarily-open estuary: nutrient versus light limitation . Marine Ecology-Progress Series , 223 : 39 – 48 .
- Perissinotto , R , Nozais , C and Kibirige , I . 2002 . Spatio-temporal dynamics of Phytoplankton and microphytobenthos in a South African temporarily-open estuary . Estuarine Coastal and Shelf Science , 55 : 47 – 58 .
- Roast , SD , Widdows , J and Jones , MB . 1999 . Respiratory responses of the estuarine mysid Neomysis integer (Peracarida: Mysidacea) in relation to a variable environment . Marine Biology , 133 : 643 – 649 .
- Roy , PS , Williams , RJ , Jones , AR , Yassini , I , Gibbs , PJ , Coates , B , West , RJ , Scanes , PR , Hudson , JP and Nichol , S . 2001 . Structure and function of south-east Australian estuaries . Estuarine Coastal and Shelf Science , 53 : 351 – 384 .
- San Vicente , C and Sorbe , JC . 1999 . Spatio-temporal structure of the suprabenthic community from Creixell beach (western Mediterranean) . Acta Oecologica-International Journal of Ecology , 20 : 377 – 389 .
- Schallenberg , M , Larned , ST , Hayward , S and Arbuckle , C . 2010 . Contrasting effects of managed opening regimes on water quality in two intermittently closed and open coastal lakes . Estuarine, Coastal and Shelf Science , 86 : 587 – 597 .
- Scharler , UM and Baird , D . 2003 . The influence of catchment management on salinity, nutrient stoichiometry and phytoplankton biomass of Eastern Cape estuaries, South Africa . Estuarine, Coastal and Shelf Science , 56 : 735 – 748 .
- Sutherland , DL and Closs , GP . 2001a . Diel patterns of mysid drift (Crustacea: Mysidacea) in the Taieri River estuary, New Zealand . New Zealand Journal of Marine and Freshwater Research , 35 : 197 – 200 .
- Sutherland , DL and Closs , GP . 2001b . Spatial and temporal variation in the abundance and composition of ichthyoplankton in a large South Island, New Zealand river estuary . New Zealand Journal of Marine and Freshwater Research , 35 : 1061 – 1069 .
- Teske , PR and Wooldridge , T . 2001 . A comparison of the macrobenthic faunas of permanently open and temporarily open/closed South African estuaries . Hydrobiologia , 464 : 227 – 243 .
- Thomas , CM , Perissinotto , R and Kibirige , I . 2005 . Phytoplankton biomass and size structure in two South African eutrophic, temporarily open/closed estuaries . Estuarine Coastal and Shelf Science , 65 : 223 – 238 .
- Webb , BF . 1973 . Fish populations of the Avon-Heathcote Estuary 3. Gut contents . New Zealand Journal of Marine and Freshwater Research , 7 : 223 – 234 .