Abstract
Single reaches in three lowland streams in Canterbury, New Zealand, were enclosed with stop-nets and fish removed by repeated electric fishing passes through the day and following night during base flow conditions in late summer. The proportion of fish caught in each pass varied between species and increased with fish size. Electric fishing efficiency declined in the second and third daytime passes, to 0.73 and 0.79 of the first pass respectively, but increased in the first and second night-time passes to 1.87 and 2.02. Population estimates, based on the first two or three daytime passes, accounted for only 62% and 82% of the population estimated using all five day and night passes. A high percentage of small benthic species remained hidden within the substrate during the day but emerged at night and became more vulnerable to electric fishing. Single or multiple daytime passes can be used to estimate the approximate density of most native fish species and size classes in small lowland streams and rivers provided calculations take into account fish hidden within the substrate and marginal vegetation.
Introduction
Single- and multiple-pass depletion electric fishing techniques have been used for over 50 years to determine the density, biomass and habitat requirements of native fish in New Zealand streams and rivers (Burnet Citation1952). Single-pass techniques are commonly used during synoptic surveys while the more labour-intensive and expensive multiple-pass and mark–recapture methods are used less frequently (Jowett & Richardson Citation1996; Glova et al. Citation1998; David et al. Citation2010).
Population estimates are subject to several potential errors. Multiple-pass estimates can be biased because of declines in electric fishing efficiency in later passes (Schnute Citation1983; Riley & Fausch Citation1992; Peterson et al. Citation2004; Mantyniemi et al. Citation2005; Rosenberger & Dunham Citation2005) and under these circumstances, four or more passes are needed to calculate fish abundance (Otis et al. Citation1978; Rosenberger & Dunham Citation2005). Moreover, an unknown proportion of fish may remain buried in the substrate and are unaffected by electric fishing because the substrate acts as a conductive shield (Lobon-Cervia et al. Citation1990; Naismith & Knights Citation1990; Beaumont et al. Citation2002). In addition, quantitative electric fishing surveys are rarely undertaken at night when benthic species emerge from cover and become more vulnerable to capture. Recent studies in the Waipara and Ashley Rivers in the South Island of New Zealand (Davey et al. in press) showed that daytime electric fishing was less effective than at night. Catch rates at night increased by 67% for bluegill bullies (Gobiomorphus hubbsi), 62% for torrentfish (Cheimarrichthys fosteri) and 37% for upland bullies (Gobiomorphus breviceps). Also studies elsewhere have shown that electric fishing at night, primarily in lakes from boats, catches more species, larger fish and greater numbers of fish than fishing during the day (Reynolds Citation1996; Beaumont et al. Citation2002).
Accurate information on the density and biomass of native fish is required to monitor the effects of water abstraction, agricultural and urban developments, and other changes on lowland and coastal streams and rivers in New Zealand. The diversity and abundance of native fish in these waters are highest in riffle and run habitats (Glova Citation1988) and many species are relatively small, benthic dwelling and nocturnally active (McDowall Citation1990). Electric fishing is used extensively throughout New Zealand to determine the diversity and abundance of introduced and native fish but sampling efficiency has rarely been examined (Jowett & Richardson Citation1996; Jowett et al. Citation2005; Graynoth et al. Citation2008a) and no previous studies have been undertaken on the effectiveness of consecutive day/night electric fishing. Therefore, the objectives of this study were to determine the efficiency of electric fishing in lowland streams for a range of different species and sizes of native fish. Specific aims were to determine trends in efficiency during multiple-passes and to determine the proportion of fish that were hidden within the substrate and unavailable to daytime electric fishing.
Methods
Study area
Single reaches in three lowland Canterbury streams and rivers were stop-netted and repeatedly electric fished through the day and following night, during base flow conditions in late summer. Birdlings Brook is a small groundwater fed tributary of Lake Ellesmere (A) (172°18′ E, 43°47′ S) and Pigeon Bay Stream is a small rain-fed hill stream on Banks Peninsula (B) (172°53′ E, 43°41′ S), whereas the Waipara River (C) (172°46′ E, 43°07′ S) is a small gravel bed river situated 50 km north of Christchurch. Both Pigeon Bay Stream and the Waipara River have been intensively electric fished in recent years and were known to support a high diversity and density of fish (Glova et al. Citation1998; Jowett et al. Citation2005; Graynoth et al. Citation2008a). The particular reaches were chosen for study because of their accessibility and the lack of deep unfishable pools.
Figure 1 Photographs of the reaches studied in A, Birdlings Brook; B, Pigeon Bay Stream and C, the Waipara River showing variations in channel form, riparian vegetation and substrate composition.

Measurements taken of random widths (n=15−46) and depths (n=41−115) within each reach (). Substrate composition for each reach was estimated visually by two observers as the percentage of vegetation, silt (<0.0625 mm), sand (0.0625–2 mm), fine gravel (2–10 mm), coarse gravel (10–64 mm), cobble (64–264 mm) and boulder (>264 mm).
Table 1 Physical features of reaches surveyed. Boulder (>264 mm), cobble (64–264 mm), coarse gravel (8–64 mm), fine gravel (2–8 mm), sand (0.06–2 mm) and silt (<0.06 mm).
The composition of the stream bed varied between streams (, ). Pigeon Bay Stream was primarily composed of boulders and cobbles, Birdlings Brook was composed of fine gravel, silt and macrophytes, whereas the riffle and run surveyed in the Waipara River was intermediate in composition and comprised mainly cobbles. All three streams contained clear water and were relatively shallow (mean depths 0.10–0.19 m) and slow flowing (0.14–0.31 m/s).
Electric fishing methods
The initial study was carried out in Pigeon Bay Stream in January 2009 (). The reach was enclosed for the entire duration of the study with dual stop-nets (2–5-mm mesh). Single, 2-mm mesh stop-nets were used at the top and bottom of the reaches in Birdlings Brook and the Waipara River in February 2010.
Each reach was electric fished three times during the day (passes 1–3) and twice at night (passes 4 and 5). In Pigeon Bay Stream we used a generator-powered, 300 W, electric-fishing machine fitted with a single, semi-circular, pole-mounted anode (45 cm maximum diameter) generating 220 volts pulsed DC (100 Hz) with a pulse width of 2 ms. The negative earth was placed downstream, outside of the study section. A portable, battery-powered, machine (Kainga 300, NIWA Instrument Systems, Christchurch, New Zealand) with the same electrical output and anode was used in Birdlings Brook and the Waipara River. There was no difference in ‘fishing power’ between the generator or backpack machine in these relatively shallow streams and the fish responded to the electrical current in a similar fashion. Most members of the electric fishing team of three to five people had >30 years fishing experience and all were equipped with Polaroid glasses. We fished 3-m-long, 1–2-m-wide lanes across the entire width of the stream and then worked upstream in a zigzag fashion (David et al. Citation2010). At night we used LED headlamps, hand-held 9V lamps and a LED torch attached to the hand-held anode. A submerged LED light on a pole was also used at night in the Waipara River. The side illumination and lack of surface reflection enabled us to see the escape reaction of fish—fish that were not stunned tended to follow the lines of the current and either dived into the substrate or moved to the left or right of the field.
Fish were attracted to the anode, stunned and collected in small dip-nets and in hand-held, 1-m-wide stop-nets (2.5-mm mesh). These are small versions of two-person apron seines (lipseins) (Elson Citation1962) and are very effective for catching small fish, especially in fast-flowing riffles and runs (Amiro Citation1990). Several sweeps were made through each lane and we attempted to capture all visible and stunned fish using hand nets and the portable stop-net. Once we had caught all visible fish and could no longer draw any more out of cover, we then moved sideways to the next lane. Benthic species that avoided the nets tended to dart into the substrate and fish that escaped capture were left after 20–40 s searching. No large shoals of fish were present and net saturation did not occur (Beaumont et al. Citation2002). Each pass ranged from 61 to 165 min, which included all the time spent electric fishing, netting and placing the catch into buckets (). Efforts were made to minimise variation in time spent fishing during successive passes but inevitably differences in the number of fish present and the increased difficulty of fishing at night meant there were some differences in time spent fishing per pass. The entire stream bed was fully accessible and all habitats were thoroughly fished. Rest periods between the start of passes 1 and 2 averaged 150 min and 125 min between the start of passes 2 and 3, respectively. We then waited 300 min until it was fully dark before commencing pass 4 and another 155 min before starting pass 5.
Table 2 Duration of fishing effort (minutes) and total catch in each pass.
Fish were anesthetised using 2-phenoxyethanol, identified, measured (total length, TL; ±1.0 mm) and held in live boxes for the duration of each survey. It was difficult to identify the smallest bullies (Gobiomorphus) (<30 mm) to species in the field and while most were classified as upland bullies a small percentage may have been common bullies (Gobiomorphus cotidianus). For data analysis, the eels were divided into three size classes (with size class limits varying between species and river) in order to determine the effects of repeated electric fishing on different sizes of fish (Peterson et al. Citation2004). The weight of individual eels was estimated from length–weight equations (Francis & Jellyman Citation1999).
Data analysis
Fish population densities and electric fishing efficiency were estimated using depletion sampling techniques (Seber Citation1982). Daytime population estimates were based on either the first two passes using the Zippin maximum likelihood method (Zippin Citation1956) or the first three passes using a maximum likelihood jack-knife method (Seber Citation1982; Pollack & Otto Citation1983). Population estimates with loge-based confidence intervals were calculated using the CAPTURE module of the MARK programme www.cnr.colostate.edu/~gwhite/mark/mark.htm (accessed 2 March 2009; Rexstad & Burnham Citation1991). These calculations assume populations were closed; fishing effort was equal in all passes; and the probability of capture of individual fish remained constant (Zippin Citation1958; Burnham et al. Citation1987).
The total population for each species and size class (N) was calculated by adding the total catch during the day to Zippin estimates of the fish remaining at night, based on passes 4 and 5. Increases in catches between passes 4 and 5 (Appendix) caused the failure of the Zippin model (which relies on a decrease in consecutive catches) and therefore N and upper confidence intervals (95%) were determined subjectively for four of the 27 species and size classes examined. Electric fishing efficiency values for day and night passes were then calculated using cumulative catches and N.
We hypothesised that electric fishing efficiency (p) depended upon the species and size of fish present, and environmental factors associated with the river and the time of fishing—such as light levels, water turbidity, depths, velocities, substrate composition and instream cover (Peterson et al. Citation2004). To test these hypotheses, catches in passes 1–5 (n 1−5) were related to the estimated number of fish remaining in the reach at the start of each pass (nr), the mean length of fish caught during that pass (ml) and to four categorical factors: species caught (sp), a species grouping based on their vulnerability to fishing (efgp), river (r) and pass number (pn). Generalised additive and linear models with a negative binomial distribution and a fixed theta were used in the MASS package in R (Venables & Ripley Citation2002, R Development Core Team Citation2009). The estimated number of fish remaining in the reach (nr) was logged and introduced into the regression model as an offset variable. The offset is distinguished from other coefficients in the regression model by being carried through as a constant with a coefficient of 1.0. To log offsets with zero values, 0.5 was added to four zero estimates of the numbers remaining (total sample size n=108). The models have the algebraic form of:
where β0 is a constant and a, b and c are coefficients.
A negative binomial model with an offset was adopted to avoid problems associated with the calculation of ratios such as p (Green Citation1979; Jackson et al. Citation1990). A variety of likely candidate models were developed and compared following information-theoretic methods using Akaike's information criterion and Akaike weights corrected for small sample bias (AICc) (Hilborn & Mangel Citation1997; Burnham & Anderson Citation2002). Estimated p values (p est ) and confidence limits were calculated from these models by dividing estimated catches by nr. The adequacy of the regression models and negative binomial distribution were tested by plotting residuals against fitted values and from quantile–quantile plots.
Results
Species present and trends in electric fishing catches
Nine species of native fish were caught in Pigeon Bay Stream, eight in the Waipara River but only three in Birdlings Brook, which also supported low numbers of juvenile (0+) brown trout (Salmo trutta), an introduced species (). Fish were present at exceptionally high densities, compared with other streams in New Zealand (Jowett & Richardson Citation1996) and large numbers were caught; 411 in Birdlings Brook (1.5 per m2), 1159 in Pigeon Bay Stream (5.7 per m2) and 2251 in the Waipara River (12.1 per m2) (Appendix).
Table 3 Fish species caught, scientific names, size classes (mm) of eels and short codes.
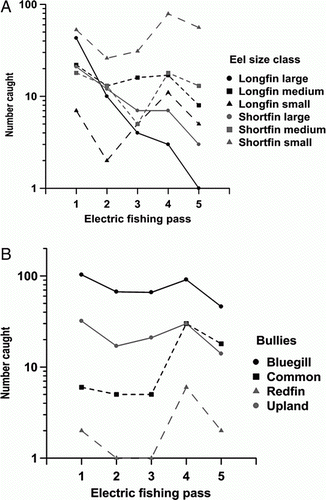
Representative trends in catches of bullies and different size classes of eels over successive passes are shown in . Catches usually declined steeply between the first and second pass followed by a gentle decline, or increase, in catch between the second and third pass (, Appendix). Catches then increased substantially in the first night-time pass (No. 4), often exceeding catches in the first pass, and then declined in the fifth and final pass. The time spent fishing declined in most reaches in the second and fifth pass but increased in the fourth pass () and was positively related to catches (r=0.29 − 0.80). Catch rates (fish per min) declined in repeated passes during each day and night period as fish were removed from the reach.
Figure 2 Trends in catches (log) of different size classes and species of eels and species of bullies in consecutive electric fishing passes in Pigeon Bay Stream. Passes 1–3 were during the day while passes 4 and 5 were at night.
Effects of repeated passes on length frequency of fish caught
Large eels were more vulnerable to capture than smaller eels and were removed from the population at a faster rate (). The mean length of eels declined by between 10 and 40 mm in consecutive passes but there were only small declines in the lengths of most other species (Appendix and ). Common and upland bullies, inanga (Galaxias maculatus) and Canterbury galaxias (Galaxias vulgaris) declined by 0.8–6.1 mm in mean length between consecutive passes while bluegill bullies increased by 0.5 mm between passes.
Figure 3 Dot plot of the length frequency of longfin eels caught in consecutive passes in the three reaches combined.
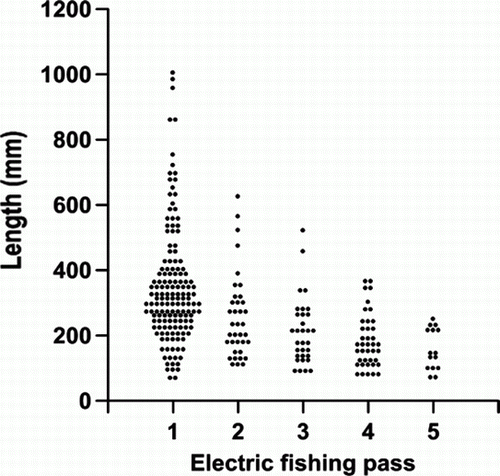
Table 4 Statistical significance and magnitude of changes in mean length (mm) in consecutive passes; analysis of covariance using river as a categorical factor.
Population estimates derived from five-pass day and night fishing
Initial analyses showed that fish species and size classes could be divided into three electric fishing efficiency groups (efgp) (low, medium and high). Canterbury galaxias and torrentfish were relatively easy to catch and were classified into a high-efficiency group while juvenile lamprey (ammocoetes) (Geotria australis), inanga, small and medium-sized shortfin eels (Anguilla australis) were classified into a low-efficiency group.
Minimum and maximum 95% confidence intervals (CI) ranged from 98% to 108% of N for the high-efficiency group (n=3) to 96% to 120% for the medium-efficiency group (n=18), whereas the low-efficiency group (n=6) ranged from 79% to approximately 203%. Therefore, N was accurately predicted for most species and size classes of fish. However, there was considerable uncertainty about the abundance of several fish species in Pigeon Bay Stream. Catches of ammocoetes remained almost constant throughout all passes (Appendix) and the slow decline in catches of small and medium-sized shortfin eels () made it difficult to estimate the total numbers present.
Efficiency of electric fishing
Akaike's information criteria for the various candidate models () showed model 1 was the most plausible. Negative binomial models provided a good fit to the data with reasonably normally distributed residuals. The residuals of model 1 became over dispersed (dispersion parameter of 1.45) when a quasi-Poisson model (Venables & Ripley Citation2002) was used. There was no evidence for any curvilinear relationships between the residuals of model 1 and either fish length or the numbers of fish remaining.
Table 5 Akaike's information criterion (AICc)and Akaike weights for competing negative binomial models (theta = 58) of the catches of each species and size classes (n 1 − 5) in each electric fishing pass.
Model 1 showed that electric fishing efficiency was highest for large fish (, ) and for night-time fishing. Efficiency in the second and third passes averaged 0.73 and 0.79 of the first pass (i.e. exponent of coefficients in ). Pass 4, the first night pass, was 1.87 more efficient than pass 1 and pass 5 was of similar high efficiency (2.02).
Figure 4 Estimated electric fishing efficiency (±1 SE) for pass 1 plotted against fish length (mm) for three species and size groups. High, torrentfish and Canterbury galaxias; low, small and medium shortfin eels, ammocoetes and inanga; medium, longfin and large shortfin eels, bullies and juvenile brown trout ().
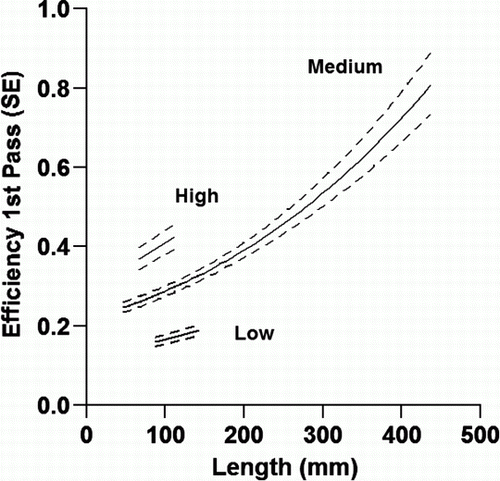
Table 6 Coefficients of model 1 (Table 5) predicting catch from fish remaining.
There was some support for candidate models that included the river and numbers of fish remaining (models 2 and 3), but virtually no support for the remaining models that included length interactions and individual fish species (). Model 2 () indicated efficiency was highest in Birdlings Brook and slightly, but not significantly, lower (0.87) in the other two rivers. Model 3 indicated that efficiencies declined slightly (a drop in p of less than 0.03) when high densities of fish were present.
Population estimates derived from single and multiple-pass daytime electric fishing
Estimates of first-pass electric fishing efficiency can be calculated from model 1 () and used to predict the total numbers present (i.e. N=n 1/p). Depending on size and species, from 16% to 81% of the fish present were caught in the first pass (average 30%). An alternative and preferred approach is to use the strong correlation (r=0.96) between catches in the first pass (n 1) and the total numbers present (N). Coefficients for the best predictive equation are given in .
Table 7 Coefficients of negative binomial equations predicting the total population present (N) from the numbers, size and species of fish caught in (a) the first daytime pass (b) population estimates derived from passes 1 and 2 (Zippin method) (c) population estimates derived from passes 1–3 (Pollack & Otto method).
Population estimates derived from removal estimates in multiple daytime passes underestimated N. Two-pass Zippin estimates averaged 62% of N (range 19–102%), whereas three-pass Pollack & Otto estimates were also low and averaged 82% of N (range 24–110%). Excluding inanga (24%), because of the presence of possible migrants in night-time passes (see below), the lowest three-pass estimates recorded were for small shortfin eels (47%), medium-sized shortfin eels (53% and 87%), redfin bullies (Gobiomorphus huttoni) (50%), common bullies (31% and 86%), bluegill bullies (61% and 88%) and ammocoetes (81%).
Multiple-pass population estimates were therefore corrected using information on fish size and efficiency of capture (b and c). These revised estimates were similar to, or slightly better, than estimates derived from first pass catches. The percentage deviance explained increased slightly from 94.6% for the first-pass method to 96.0% for the three-pass method.
Eel biomass estimates
Eels were the largest fish present and totalled 88% of the biomass of fish caught. There was a rapid decline in the biomass of eels caught in consecutive passes during both the day and night (). The biomass of eels caught in the first pass ranged from 43% to 92% of the total and was strongly related (r=0.92) to the log of the mean weight of eels present (). However, the biomass caught in the first pass at night (pass 4) was usually higher than expected (based on daytime catches), indicating that more eels became available and vulnerable to electric fishing at night.
Table 8 Biomass (g) of eels removed in consecutive passes.
Discussion
Factors causing variations in electric fishing efficiency
Species behaviour
The major variations in electric fishing efficiency between species were probably related to differences in fish behaviour and the habitats occupied during the day and night. Torrentfish were particularly susceptible to electric fishing possibly because they are relatively large (mean length 74 mm) and live in shallow, fast-flowing riffles (Jowett & Richardson Citation1995), where stunned fish are easily caught in a hand-held stop-net. Canterbury galaxias were also easy to catch possibly because they feed in open water on invertebrate drift (Glova & Sagar Citation1989b; Glova et al. Citation1992).
Benthic species that live in the interstitial spaces beneath coarse substrates in riffles and runs, such as juvenile longfin eels, torrentfish, bluegill, redfin and upland bullies (McDowall Citation1990), were more vulnerable to electric fishing than subterranean species, such as small shortfin eels and ammocoetes, which burrow in fine gravels and silt in stream margins and slow flowing pools. Electric fishing is known to be inefficient for small eels (Lobon-Cervia et al. Citation1990), especially small shortfins that live a cryptozoic life (Burnet Citation1952; Glova & Jellyman Citation2000), and for ammocoetes that require pulsed applications of power and repeated passes to draw them from their burrows to the surface for capture (Weisser & Klar Citation1990; Kelso & Todd Citation1993; Henderson et al. Citation2007).
Free swimming, pool-dwelling, pelagic species, such as inanga and adult trout, can also be difficult to catch and often require multiple anodes or boat mounted electric fishing equipment (Burnet Citation1968; Mahon Citation1980; Jowett Citation2002). We found an increase in inanga catches at night, especially in the Waipara River, and suspect this may be related to the nocturnal downstream migration of mature fish. One inanga was found gilled in the top stop-net in the Waipara River at the end of the study and others may have penetrated this stop-net. The survey was undertaken 4 days before the new moon and might have coincided with the downstream migration of fish from the main river to their estuarine spawning areas (Burnet Citation1965; Taylor Citation2002).
Fish size and density
Electric fishing is highly efficient for large fish because their size makes them more susceptible to electric current, they are attracted from a greater distance and remain stunned for a longer period and are more visible to fishers (Burnet Citation1952, Citation1959; Vibert Citation1963; Mahon Citation1980). Thus we found that electric fishing was most effective for large fish, like adult longfin eels Anguilla dieffenbachii, but was ineffective for small bully fry (<20 mm). Although fish density varied substantially between species and rivers, density had little effect on electric fishing efficiency in this study, indicating that simple ratios of catch to the population present could be used to determine electric fishing efficiency.
Environmental factors
With the exception of bully fry, most fish were not visible during the day and hid beneath the substrate or marginal vegetation. This is typical behaviour, since many native fish such as eels, torrentfish, redfin bullies and galaxiids are nocturnally active, feeding or moving to different habitats at night (Glova et al. Citation1987; Glova & Sagar Citation1989a; Glova & Jellyman Citation2000; McCullough & Hicks Citation2002). Therefore, it was not unexpected to find a substantial increase in electric fishing efficiency at night as fish emerged from cover.
Previous studies, in a range of New Zealand rivers, found there was no significant variation in electric fishing efficiency with substrate size or depth, but that efficiency increased in slow flowing water (Jowett & Richardson Citation1996). In the Waipara River, efficiency did not vary between seasons and flows (Jowett et al. Citation2005) with the exception of upland bully where efficiency was highest in winter, probably because of reductions in periphyton cover and increases in fish size. We found that efficiency was highest in Birdlings Brook and suspect, based on electric fishing studies elsewhere (Burnet Citation1952; Jowett & Richardson Citation1996), this could be related to its relatively narrow width (2.76 m), low water velocities and finer substrate with less interstitial spaces for fish to hide. However, more rivers would need to be surveyed to measure the relative effects on efficiency of these and other environmental factors such as instream cover, surface turbulence, water temperatures, conductivity and turbidity (e.g. Rodgers et al. Citation1992; Hayes & Baird Citation1994; Peterson et al. Citation2004; Rosenberger & Dunham Citation2005).
Sampling gear and methods used
The electric fishing equipment and procedures used in this study were broadly similar to those employed elsewhere in New Zealand (Glova & Duncan Citation1985; Jowett & Richardson Citation1996; Glova et al. Citation1998; Hickey & Closs Citation2006; David et al. Citation2010). However, the study objectives and high densities of small fish present meant that our surveys took longer than usual. The first pass took 44–64 min per 100 m2 compared with 10 min per 100 m2 in other surveys (David et al. Citation2010). The total number of fish collected ranged from 83% to 95% of the estimated total population (N) and was similar to, or exceeded, population estimation guidelines of 60–85% (Mahon Citation1980; Cowx Citation1983) and, in addition, catches in the last pass declined by more than 50% for 15 of the 27 samples, thereby agreeing with other removal sampling criteria (Rodgers et al. Citation1992; Jowett & Richardson Citation1996).
Electric fishing has the potential to injure and kill fish (Burnet Citation1952; Vibert Citation1963) and sufficient time should be left between multiple passes for fish to recover, with recommended rest periods ranging from 30 min to 2 h (Peterson & Cederholm Citation1984; Bohlin et al. Citation1989; Odenkirk & Smith Citation2005). Although rest periods during this study were relatively long (2–5 h), not all fish survived and we observed two dead longfins (183 and 244 mm), a few dead bullies and one torrentfish in Pigeon Bay Stream. These fish could have been killed either by electric fishing or by being trampled on by the fishing team.
Most species of native fish are sedentary during the day and may make small, limited movements at night (Glova et al. Citation1987; Glova & Sagar Citation1989a; McDowall Citation1990; Davey et al. in press). However, large eels are the exception and can move substantial distances from daytime resting habitat in pools to feed in riffles and runs at nights (Jellyman & Sykes Citation2003, Graynoth and Jellyman pers. obs.). The stop-nets used in this study appeared to be fish-tight for most species, with the possible exception of inanga, and would have minimised potential biases in population estimates arising from emigration and immigration.
Variations in efficiency between passes
We found major differences in pass efficiency with declines in the proportion of fish caught in the second and third daytime passes and increases in the two night-time passes. Declines in efficiency during daytime surveys have often been reported elsewhere and are caused by changes in physiology and fish behaviour (Schnute Citation1983; Bohlin & Cowx Citation1990; Riley & Fausch Citation1992). For example, previously shocked fish may actively avoid the current and burrow deeper within the substrate or cover (Mahon Citation1980; Peterson et al. Citation2004).
The increased availability of fish at night in New Zealand rivers has not been previously noted, with the exception of CitationDavey et al. (in press). Insufficient passes were undertaken at night to detect declines in efficiency, but as only small numbers of fish were caught during the fifth pass, we suspect total population estimates for most species were close to the actual numbers present.
Effectiveness of daytime single-pass fishing compared with multiple-pass day and night surveys
There are several advantages in undertaking single-pass rather than multiple-pass surveys. Single-pass surveys can save time and costs, increase spatial coverage and can increase the precision of total population estimates in large catchments (Mitro & Zale Citation2000; Reid et al. Citation2008). Because of this, single-pass surveys and generalised models of capture efficiency have often been used (Jowett & Richardson Citation1996; Kruse et al. Citation1998; Rosenberger & Dunham Citation2005; Laplanche Citation2010). We found little evidence for differences in electric fishing efficiency between rivers, which suggests single-pass surveys during the day could be used to predict fish numbers and sizes, at least in rivers similar to those studied. However, multiple-pass (probably four- to six-pass) surveys are still needed for burrowing species, such as small shortfin eels and ammocoetes, and when precise population estimates are required in specific locations.
Although single-pass surveys at night would be more efficient than daytime surveys, operator safety and access issues could make these impractical in some areas.
Implications for previous estimates of fish densities in New Zealand rivers
Previous investigations on the density of native fish in New Zealand rivers assumed electric fishing efficiency remained constant in all passes and made no adjustments for fish hidden within the substrate (Jowett & Richardson Citation1996; Jowett et al. Citation2005; Graynoth et al. Citation2008a). Multiple-pass daytime surveys, in riffles and runs within 38 medium to large New Zealand rivers, detected no significant difference in capture probability between species (Jowett & Richardson Citation1996). Efficiencies for eight native fish species averaged 0.75 (range 0.54–0.86) and were substantially higher than those recorded for the same species in the present study. Similarly multiple daytime pass efficiencies for five native fish species caught in riffles and runs in the Waipara River (Jowett et al. Citation2005) averaged 0.59 (range 0.38–0.74) and were almost double those found for these species in the present study (average 0.30, range 0.18–0.40). This is presumably because no account was made for declines in efficiency in later passes and fish hidden in the substrate.
Previous estimates of the survival, density and biomass of longfin eels in rivers and streams throughout New Zealand were also based on single-pass surveys but took into account changes in efficiency with eel length (Graynoth & Niven Citation2004; Graynoth et al. Citation2008b; Graynoth & Booker Citation2009). Based on the present study, the equations used (i.e. p=0.451+0.000867ml and p=0.25+0.00118ml) (Graynoth et al. Citation2008a) appear to overestimate fishing efficiency for small eels and underestimate efficiency for large fish. Tests showed this overestimate increased biomass estimates by a minor amount (~2%) because over 65% of the biomass is derived from relatively large eels (>400 mm) that are easily caught.
In conclusion, although the densities of small fish may have been underestimated in previous studies, the relative abundance of fish and daytime habitat preferences should remain largely unchanged. This is because first-pass abundance and multiple-pass population estimates are closely related to the actual populations present, as shown in the present study.
Species diversity
All fish species present were caught in the first daytime electric fishing pass, with the single exception of inanga in the Waipara River. As discussed this could be related to the downstream spawning migration of mature inanga from upstream pools into the reach surveyed. However, a single giant bully Gobiomorphus gobioides (Valenciennes, 1837) was captured in a fyke net situated just downstream of the 48-m reach electric fished in Pigeon Bay Stream. This supports recommendations that relatively long reaches, of about 150 m, should be fished to determine species richness (Patton et al. Citation2000; David & Hamer Citation2010) using a variety of both intensive and extensive methods (David et al. Citation2002; McCullough & Hicks Citation2002; Hickey & Closs Citation2006).
Mark–recapture methods
Further work is needed to determine the effectiveness of native fish population estimates based on the combination of removal and mark–recapture techniques (Otis et al. Citation1978; Bohlin et al. Citation1989; Rosenberger & Dunham Citation2005). Mark–recapture techniques provided better population estimates for subterranean and surface dwelling terrestrial salamanders than removal methods (Bailey et al. Citation2004) and the same may be true for crayfish (Rabeni et al. Citation1997) and native fish in New Zealand. Mark–recapture methods are also more precise than the three-sample removal method when the probability of capture is less than 0.30 (Seber Citation1982), as is the situation with many small native fish.
On the other hand, mark–recapture methods are relatively slow and expensive compared with removal methods and some native fish are too small or difficult to individually mark with fin clips, tags or latex injections (Burnet Citation1969; McEwan Citation2009). Also fish behaviour can be altered for up to 15–24 h by capture, handling and marking and this could introduce additional error into population estimates (Mesa & Schreck Citation1989; Bohlin & Cowx Citation1990; Peterson et al. Citation2004; Rosenberger & Dunham Citation2005).
Conclusions
Single or multiple daytime passes can be used to estimate the approximate density of most native fish species and size classes in small lowland streams and rivers, provided population estimates take into account the decline in efficiency in later passes and the numbers of fish concealed within the substrate and instream cover. However, multiple day/night passes are needed if the objective is to determine the density of burrowing species such as small shortfin eels and ammocoetes. Multiple day/night passes may also be required if there are large areas of complex habitat and coarse substrate where fish can hide. Also, with one exception, all fish species present were caught in the first daytime electric fishing pass.
Further studies, possibly using PIT tags and mark–recapture techniques, are needed to determine the numbers, size and species of native fish that live an entirely subterranean existence in the shallow hyporheic zone and remain inaccessible both to day and night-time electric fishing.
Acknowledgements
We thank NIWA staff, Lindsay Hawke, Julian Sykes, Doug Booker, Joanna Lessard and Peter Robinson for assistance in the field and Andrew Davey for copies of his draft manuscript on day night habitat use in the Waipara and Ashley Rivers. Doug Booker and Clive Howard-Williams reviewed the manuscript. We also appreciate the helpful and insightful comments made by two anonymous referees. This study was partially supported by the FRST Taonga Species research programme (Contract C01×0511) and FRST Capability Fund (2009) 0, Fund 231).
References
- Amiro , PG . 1990 . “ Evaluation of some electrofishing techniques used to estimate populations of juvenile Atlantic salmon in enclosed areas of streams ” . In Developments in electric fishing , Edited by: Cowx , IG . 174 – 85 . Oxford : Fishing News Books .
- Bailey , LL , Simons , TR and Pollock , KH . 2004 . Comparing population size estimators for plethodontid salamanders . Journal of Herpetology , 38 : 370 – 380 .
- Beaumont WRC , Taylor AAL , Lee MF , Welton JS 2002 . Guidelines for electric fishing best practice . Environment Agency R&D Technical Report W2-054/TR .
- Bohlin , T and Cowx , IG . 1990 . “ Implications of unequal probability of capture by electric fishing on the estimation of population size ” . In Developments in electric fishing , Edited by: Cowx , IG. 145 – 155 . Oxford : Fishing News Books .
- Bohlin , T , Hamrin , SF , Heggberget , TG , Rasmussen , G and Saltveit , SJ . 1989 . Electrofishing—theory and practice with special emphasis on salmonids . Hydrobiologia , 173 : 9 – 43 .
- Burnet , AMR . 1952 . Studies on the ecology of the New Zealand freshwater eels 1. The design and use of an electric fishing machine . Australian Journal of Marine and Freshwater Research , 3 : 111 – 125 .
- Burnet , AMR . 1959 . Electric fishing with pulsatory direct current . New Zealand Journal of Science and Technology , 2 : 46 – 56 .
- Burnet , AMR . 1965 . Observations of the spawning migrations of Galaxias attenuatus (Jenyns) . New Zealand Journal of Science , 8 : 79 – 87 .
- Burnet AMR 1968 . A study of the relationships between brown trout and eels in a New Zealand stream . New Zealand Marine Department Fisheries Technical Report 26 .
- Burnet , AMR . 1969 . The growth of New Zealand freshwater eels in three Canterbury streams . New Zealand Journal of Marine and Freshwater Research , 3 : 376 – 384 .
- Burnham , KP and Anderson , DR . 2002 . “ Model selection and multimodel inference ” . In A practical information-theorectic approach , 2nd edition , New York : Springer .
- Burnham , KP , Anderson , DR , White , GC , Brownie , C and Pollock , KH . 1987 . Design and analysis methods for fish survival experiments based on release-recapture , Bethesda , MD : American Fisheries Society .
- Cowx , IG . 1983 . Review of the methods for estimating fish population size from survey removal data . Fisheries Management , 14 : 67 – 82 .
- Davey AJH , Booker DJ , Kelly D in press . Diel variation in stream fish microhabitat selection: implications for instream flow assessment . : Aquatic Conservation: Marine and Freshwater Ecosystems .
- David , BO , Closs , GP and Arbuckle , CJ . 2002 . Distribution of fish in tributaries of the lower Taieri/Waipori Rivers, South Island, New Zealand . New Zealand Journal of Marine and Freshwater Research , 36 : 797 – 808 .
- David BO , Hamer M 2010 . Regional guidelines for ecological assessments of freshwater environments . Standardised fish monitoring for wadeable streams . Environment Waikato Regional Council Report ISSN 1177–9284 .
- David , BO , Hamer , MP , Collier , KJ , Lake , MD , Surrey , GM , McArthur , K , Nicholson , C , Perrie , A and Dale , M . 2010 . A standardised sampling protocol for robust assessment of reach-scale fish community diversity in wadeable New Zealand streams . New Zealand Journal of Marine and Freshwater Research , 44 : 177 – 187 .
- Elson , PF . 1962 . The Pollett Apron Seine . Journal of Fisheries Research Board of Canada , 19 : 93 – 100 .
- Francis , RICC and Jellyman , DJ . 1999 . Are mean size data adequate to monitor freshwater eel fisheries? . Marine and Freshwater Research , 50 : 355 – 366 .
- Glova , GJ . 1988 . Fish density variations in the braided Ashley River, Canterbury, New Zealand . New Zealand Journal of Marine and Freshwater Research , 22 : 9 – 15 .
- Glova , GJ and Duncan , MJ . 1985 . Potential effects of reduced flow on fish habitats in a large braided river, New Zealand . Transactions of the American Fisheries Society , 114 : 165 – 181 .
- Glova , GJ and Jellyman , DJ . 2000 . Size-related differences in diel activity of two species of juvenile eel (Anguilla) in a laboratory stream . Ecology of Freshwater Fish , 9 : 210 – 218 .
- Glova , GJ , Jellyman , DJ and Bonnett , ML . 1998 . Factors associated with the distribution and habitat of eels (Anguilla spp.) in three New Zealand lowland streams . New Zealand Journal of Marine and Freshwater Research , 32 : 255 – 269 .
- Glova , GJ and Sagar , PM . 1989a . Feeding in a nocturnally active fish Galaxias brevipinnis in a New Zealand stream . Australian Journal of Marine and Freshwater Research , 40 : 231 – 240 .
- Glova , GJ and Sagar , PM . 1989b . Prey selection by Galaxias vulgaris in the Hawkins River . New Zealand Journal of Marine and Freshwater Research , 23 : 153 – 161 .
- Glova , GJ , Sagar , PM and Docherty , CR . 1987 . Diel feeding periodicity of torrent fish (Cheimarrichthys fosteri Haast) in two braided rivers of Canterbury, New Zealand . New Zealand Journal of Marine and Freshwater Research , 21 : 555 – 561 .
- Glova , GJ , Sagar , PM and Naslund , J . 1992 . Interaction for food and space between populations of Galaxias vulgaris Stokell and juvenile Salmo trutta L. in a New Zealand stream . Journal of Fish Biology , 5 : 909 – 926 .
- Graynoth E , Booker DJ 2009 . Biomass of longfin eels in medium to large rivers . New Zealand Fisheries Assessment Report 2009/44 .
- Graynoth , E , Francis , RICC and Jellyman , DJ . 2008a . Factors influencing juvenile eel (Anguilla spp.) survival in lowland New Zealand streams . New Zealand Journal of Marine and Freshwater Research , 42 : 153 – 172 .
- Graynoth E , Jellyman DJ , Bonnett M. 2008b . Spawning escapement of female longfin eels . New Zealand Fisheries Assessment Report 2008/7 .
- Graynoth , E and Niven , K . 2004 . Habitat for female longfinned eels in the West Coast and Southland , 5 – 32 . New Zealand : Science for Conservation .
- Green , RH . 1979 . Sampling design and statistical methods for environmental biologists , New York : John Wiley and Sons .
- Hayes , JW and Baird , D . 1994 . Estimating relative abundance of juvenile brown trout in rivers by underwater census and electrofishing . New Zealand Journal of Marine and Freshwater Research , 28 : 243 – 253 .
- Henderson P , Seaby R , Somes R 2007 . Removal sampling 2. Report . Lymington , , Hampshire , Pisces Conservation Ltd .
- Hickey , MA and Closs , GP . 2006 . Evaluating the potential of night spotlighting as a method for assessing species composition and brown trout abundance: a comparison with electrofishing in small streams . Journal of Fish Biology , 69 : 1513 – 1523 .
- Hilborn , R and Mangel , M . 1997 . The ecological detective. Confronting models with data , Princeton , NJ : Princeton University Press .
- Jackson , DA , Harvey , HH and Somers , KM . 1990 . Ratios in aquatic sciences: statistical shortcomings with mean depth and the morphoedaphic index . Canadian Journal of Fisheries and Aquatic Sciences , 47 : 1788 – 1795 .
- Jellyman , DJ and Sykes , JRE . 2003 . Diel and seasonal movements of radio-tagged freshwater eels, Anguilla spp., in two New Zealand streams . Environmental Biology of Fishes , 66 : 143 – 154 .
- Jowett , IG . 2002 . In-stream habitat suitability criteria for feeding inanga (Galaxias maculatus) . New Zealand Journal of Marine and Freshwater Research , 36 : 399 – 407 .
- Jowett , IG and Richardson , J . 1995 . Habitat preferences of common, riverine New Zealand native fishes and implications for flow management . New Zealand Journal of Marine and Freshwater Research , 29 : 13 – 23 .
- Jowett , IG and Richardson , J . 1996 . Distribution and abundance of freshwater fish in New Zealand rivers . New Zealand Journal of Marine and Freshwater Research , 30 : 239 – 255 .
- Jowett , IG , Richardson , J and Bonnett , ML . 2005 . Relationship between flow regime and fish abundances in a gravel-bed river, New Zealand . Journal of Fish Biology , 66 : 1419 – 1436 .
- Kelso , JRM and Todd , PR . 1993 . Instream size segregation and density of Geotria australis ammocoetes in two New Zealand streams . Ecology of Freshwater Fish , 2 : 108 – 115 .
- Kruse , CG , Hubert , WA and Rahel , FJ . 1998 . Single-pass electrofishing predicts trout abundance in mountain streams with sparse habitat . North American Journal of Fisheries Management , 18 : 940 – 946 .
- Laplanche , C . 2010 . A hierarchial model to estimate fish abundance in alpine streams by using removal sampling data from multiple locations . Biometrical Journal , 52 : 1 – 13 .
- Lobon-Cervia , J , Bernat , Y and Rincon , PA . 1990 . Effects of eel (Anguilla anguilla L.) removals from selected sites of a stream on its subsequent densities . Hydrobiologia , 206 : 207 – 216 .
- Mahon , R . 1980 . Accuracy of catch-effort methods for estimating fish density and biomass in streams . Environmental Biology of Fishes , 5 : 343 – 360 .
- Mantyniemi , S , Romakkaniemi , A and Arjas , E . 2005 . Bayesian removal estimation of a population size under unequal catchability . Canadian Journal of Fisheries & Aquatic Sciences , 62 : 291 – 300 .
- McCullough , CD and Hicks , BJ . 2002 . Estimating the abundance of banded kokopu (Galaxias fasciatus Gray) in small streams by nocturnal counts under spotlight illumination . New Zealand Natural Sciences , 27 : 1 – 14 .
- McDowall , RM . 1990 . “ New Zealand Freshwater Fishes ” . In A natural history and guide , Auckland : Heinemann Read .
- McEwan , AJ . 2009 . Fine scale spatial behaviour of indigeneous riverine fish in a small New Zealand stream , Massey University, Master of Science in Conservation Biology .
- Mesa , MG and Schreck , CB . 1989 . Electrofishing mark–recapture and depletion methodologies evoke behavioral and physiological changes in cutthroat trout . Transactions of the American Fisheries Society , 118 : 644 – 658 .
- Mitro , MG and Zale , AV . 2000 . Predicting fish abundance using single-pass removal sampling . Canadian Journal of Fisheries & Aquatic Sciences , 57 : 951 – 961 .
- Naismith , IA and Knights , B . 1990 . Modelling of unexploited and exploited populations of eels (Anguilla anguilla L.), in the Thames Estuary . Journal of Fish Biology , 37 : 975 – 986 .
- Odenkirk , J and Smith , S . 2005 . Single- versus multiple-pass boat electrofishing for assessing smallmouth bass populations in Virginia Rivers . North American Journal of Fisheries Management , 25 : 717 – 724 .
- Otis , DL , Burnham , KP , White , GC and Anderson , DR . 1978 . Statistical inference from capture data on closed animal populations . Wildlife Monographs , 62 : 135
- Patton , TM , Hubert , WA , Rahel , FJ and Gerow , KG . 2000 . Effort needed to estimate species richness in small streams on the Great Plains in Wyoming . North American Journal of Fisheries Management , 20 : 394 – 398 .
- Peterson , JT , Thurow , RF and Guzevich , JW . 2004 . An evaluation of multipass electrofishing for estimating the abundance of stream-dwelling salmonids . Transactions of the American Fisheries Society , 133 : 462 – 475 .
- Peterson , NP and Cederholm , CJ . 1984 . A comparison of the removal and mark–recapture methods of population estimation for juvenile coho salmon in a small stream . North American Journal of Fisheries Management , 4 : 99 – 102 .
- Pollack , KH and Otto , MC . 1983 . Robust estimation of population size in closed animal populations from capture-recapture experiments . Biometrics , 39 : 1035 – 1049 .
- Rabeni , CF , Collier , KJ , Parkyn , SM and Hicks , BJ . 1997 . Evaluating techniques for sampling stream crayfish (Paranephops planifrons) . New Zealand Journal of Marine and Freshwater Research , 31 : 693 – 700 .
- R Development Core Team 2009 . R: A language and environment for statistical computing. Vienna, R Foundation for Statistical Computing . ISBN 3-900051-07-0, URL http://www.R-project.org .
- Reid , SM , Jones , NE and Yunker , G . 2008 . Evaluation of single-pass electrofishing and rapid habitat assessment for monitoring redside dace . North American Journal of Fisheries Management , 28 : 50 – 56 .
- Rexstad E , Burnham K 1991 . User's guide for interactive program CAPTURE . Colorado Fish and Wildlife Research Unit, Colorado State University , Fort Collins , CO .
- Reynolds , J . 1996 . “ Electrofishing ” . In Fisheries techniques , 2nd edition , Edited by: Murphy , BR and Willis , DW . 221 – 253 . Bethesda , MD : American Fisheries Society .
- Riley , SC and Fausch , KD. 1992 . Underestimation of trout population size by maximum-likelihood removal estimates in small streams . North American Journal of Fisheries Management , 12 : 768 – 776 .
- Rodgers , JD , Solazzi , MF , Johnson , SL and Buckman , MA . 1992 . Comparison of three techniques to estimate juvenile coho salmon populations in small streams . North American Journal of Fisheries Management , 12 : 79 – 86 .
- Rosenberger , AE and Dunham , JB . 2005 . Validation of abundance estimates from mark–recapture and removal techniques for rainbow trout captured by electrofishing in small streams . North American Journal of Fisheries Management , 25 : 1395 – 1410 .
- Schnute , J . 1983 . A new approach to estimating populations by the removal method . Canadian Journal of Fisheries and Aquatic Sciences , 40 : 2153 – 2169 .
- Seber , GA . 1982 . The estimation of animal abundance and related parameters , 2nd edition , London : Griffin .
- Taylor MJ 2002 . The national inanga spawning database: trends and implications for spawning site management . Science for Conservation Report . No. 188 .
- Venables , WN and Ripley , BD . 2002 . Modern applied statistics with S , 4th edition , London : Springer .
- Vibert , R . 1963 . Neurophysiology of electric fishing . Transactions of the American Fisheries Society , 92 : 265 – 275 .
- Weisser , JW and Klar , GT . 1990 . “ Electric fishing for sea lampreys (Petromyzon marinus) in the Great Lakes region of North America ” . In Developments in electric fishing , Edited by: Cowey , CB . 59 – 64 . Oxford : Fishing News Books .
- Zippin , C . 1956 . An evaluation of the removal method of estimating animal populations . Biometrics , 12 : 163 – 189 .
- Zippin , C . 1958 . The removal method of population estimation . Journal of Wildlife Management , 22 : 82 – 90 .