Abstract
The mitigating effects of fish passes to aid fish dispersal are well recognised; however, non-sports fish species, juveniles and weak swimmers have mostly been neglected when developing solutions to fish passage issues. We studied the juvenile (fork length, FL <60 mm) swimming ability of a weakly swimming fish species, īnanga (Galaxias maculatus), up an artificial ramp to evaluate the effects of fish size, ramp slope (5°, 15°, 25°), water velocity and the potential need for resting opportunities (pools) for a successful ascent. Water velocity significantly increased and fish passage success significantly decreased with ramp angle, with only one fish passing the 25° ramp. On average, fish had to be 10 mm larger to ascend a 15° ramp angle than at 5°. Pools did not enhance passage success in this study. To facilitate fish passage at barriers for weakly swimming fish species like juvenile īnanga, ramps should provide a rough surface substrate and have an optimal angle of 5° for a 3-m ramp.
Introduction
The alteration of river systems by physical barriers is common throughout the world and has often been linked with impacts on the freshwater fish fauna up- and downstream of these structures (e.g. reduced abundances; Allibone Citation1999; Gibson et al. Citation2005). Culverts, particularly, have been found to impede fish movement (Richmond et al. Citation2007; Poplar-Jeffers et al. Citation2009) with the most predominant impacts including high water velocities both downstream and within the culvert itself (Haro et al. Citation2004), and an impassable vertical undercut drop (perch) at the culvert outlet (David et al. Citation2009; Doehring et al. Citation2011a). Thus, much research has focused on designing ways to allow fish to pass such obstacles (Kemp & O'Hanley Citation2010). Despite engineering guidelines for stream crossings, financial constraints have commonly led to poor design and installation of fish ramps, especially in urban waterways (Haro et al. Citation2004). This has resulted in ongoing upstream passage issues for migratory freshwater fish species (Gibson et al. Citation2005), especially non-sports fish species (i.e. non-salmonids).
Migration is undertaken for the purposes of locating suitable habitat, growth, reproduction and food acquisition, and is an integral part of the life history for the majority of New Zealand's native fish species (McDowall Citation2000, Citation2001). For diadromous fish species (i.e. species that need access to and from the sea to complete their life cycle) where migration is obligatory, accessibility of larvae to downstream and juvenile accessibility to upstream habitat is crucial for maintenance of populations (McDowall Citation1998; Leathwick et al. Citation2008). More than half of New Zealand's freshwater native fish species incorporate diadromous populations (McDowall Citation1998; Joy et al. Citation2000), so ideally barrier-free waterways, or more realistically, waterways with ‘transparent’ fish passage at existing barriers are essential in allowing longitudinal migrations.
Some New Zealand migratory fish species concentrate in lowland streams to spawn (McDowall Citation1993) and those streams can be disproportionately affected by urbanisation (Knaepkens et al. Citation2006; Collier et al. Citation2009). In the same way, the presence of migration barriers can be directly related to urban land use, reductions in fish species richness can also be linked to urbanisation (Magaud et al. Citation1997; Doehring Citation2009). Improving upstream passage at culverts is not only crucial because many species are an important component of New Zealand's freshwater fish fauna and they provide a food supply for many birds and predatory fish species (Jowett Citation2002; Rowe et al. Citation2002), but also because juveniles of the five migratory galaxiids make up a substantial part of New Zealand's whitebait fishery (Rowe et al. Citation1992).
Īnanga (Galaxias maculatus), the most common species in the whitebait fishery, are found widely at low elevations in New Zealand, but also in eastern and western Australia, Tasmania, South Africa and southern South America. Īnanga are reproductively mature after 1 year when they migrate downstream to estuaries in autumn to spawn a few hundred to a few thousand eggs amongst vegetation, upstream of the tidal salt wedge (Hickford et al. Citation2010). After hatching, the larvae (c. 7 mm long) migrate to sea to feed and grow, before returning to freshwater as whitebait (c. 45–50 mm, 21–23 weeks old) (McDowall Citation2000). Stocks have substantially declined in New Zealand and South America in the past 100 years (Jowett Citation2002; Mardones et al. Citation2008), and consequently fishery authorities are keen to halt the decline and are calling for a better understanding of factors affecting fish populations such as urbanisation and impediments to migration.
It is essential to focus on the swimming behaviour and ability of juvenile (fork length, FL < 60 mm) īnanga to negotiate obstacles to better understand how this species, and other juveniles of migratory fish species, are able to overcome anthropogenic barriers. The swimming capability of fish at barriers is a major criterion when investigating passage design and depends on several environmental variables (Baker & Boubée Citation2006; MacDonald & Davies Citation2007), in particular water velocities within and downstream of a barrier (Haro et al. Citation2004; Peake Citation2004; House et al. Citation2005) and/or physiological effects related to fish size, age or fatigue (Nikora et al. Citation2003).
To appreciate fully the relationship between water velocities and the swimming ability of īnanga, there are three dominant swimming modes that need to be understood (Nikora et al. Citation2003). Sustained swimming is an aerobic motion that can be maintained for an indefinite period, burst swimming is a short, high-speed anaerobic motion that cannot be maintained for prolonged periods, and finally prolonged swimming involves both aerobic and anaerobic processes of energy supply (Weihs Citation1974). Once the different swimming modes of a target species are known, ramps can be designed accordingly to enable fish to proceed past barriers (Peake Citation2004). If ramps are to be successful, fish must be able to ascend them quickly (Baker & Boubée Citation2006). Hence, for management applications, knowledge of fatigue plays a particularly important role (Hammer Citation1995), and swimming capability of the target fish species should be considered when designing fish ramps (Peake et al. Citation1997a, Citation1997b; Tudorache et al. Citation2008).
Most New Zealand native fish do not pass barriers by jumping as salmonids do (Naughton et al. Citation2007; Kemp & Williams Citation2008) and instead some ‘climb’ vertical wet rock surfaces using either their pectoral fins (e.g. kōaro, Galaxias brevipinnis) or snake-like motions (e.g. Anguillidae) (McDowall Citation1993McDowall Citation2003; Eikaas & McIntosh Citation2006). Species that cannot ‘climb’, such as īnanga and some bully species (Gobiomorphus spp.), must burst swim past high-velocity areas.
Despite any morphological and behavioural adaptations (e.g. modified climbing or swimming abilities), many species are still unable to negotiate in-stream barriers that are perched, lack wetted margins or create sustained high-velocity flows (Boubée et al. Citation1999). Īnanga are particularly affected by high water velocities (Mitchell Citation1989; Baker & Boubée Citation2006), which are commonly encountered at culvert entrances, hindering their upstream migration.
To overcome the main issues causing culverts to be juvenile īnanga migration barriers, we focused on determining the most important variables affecting fish passage. The ultimate goal was to design more effective fish passages for use in urban stream design (Doehring et al. Citation2011a). With this in mind, the objectives were to investigate how the passage success of juvenile īnanga (FL <60 mm) was influenced by changing the angle and length of a fish ramp, whether upstream movement and therefore passage success could be improved by providing resting opportunities, and the effect of fish size on juvenile īnanga passage success.
Materials and methods
Experimental design
To determine how best to reduce the effect of migration barriers on juvenile īnanga (FL <60 mm), an artificial ramp was designed to simulate conditions fish face at migration barriers. This allowed the effects of ramp length, ramp angle and the presence of pools as resting opportunities on the success or failure of īnanga to traverse fish ramps to be tested. To measure the passage success of juvenile īnanga, a 3-m long ramp constructed from corflute (polypropylene) and artificial grass (synthetic turf, TigerTurfTM, Onehunga, New Zealand) was set at one of three different angles (5°, 15° or 25°) with and without a pool at 1.5 m distance.
The angle and pool combinations used in the experiments were suitable to study climbing behaviour of īnanga, as previous research has shown this species is able to pass a 1.5-m long ramp at 5° and 15° angle without resting opportunities (Baker & Boubée Citation2006). To traverse ramps longer than 1.5 m and steeper than 15° successfully, it was assumed, however, that they need resting pools (Baker & Boubée Citation2006). Thus, installing a pool at 25° was expected to improve passage compared with the study of Baker & Boubée (Citation2006).
The experiments were carried out in October and November 2008 at Cawthron Institute in Nelson, New Zealand, between 07:00 and 20:00 h, as whitebait generally move during daylight hours (Stancliff et al. Citation1988; Doehring et al. Citation2011b). For each trial, 10 randomly selected fish of varying size, but all FL <60 mm, were placed in a lower holding tank and given 3 h to pass the ramp. After 3 h, fish that had successfully traversed the ramps were counted and measured (mm, FL). That is, fish that ascended to the upper tank were successful, but fish that were still on the ramp, in the lower tank or in the pool after 3 h were considered to have not passed the ramp. A curtain erected beside the ramp minimised disturbance by observers.
Each day up to four experiments were run (i.e. 12-h day) with slope and pool/no pool combinations randomly assigned. This meant that each 3-h experiment had a different treatment combination (i.e. 5°, 15° or 25° angle, and either a pool or no pool; sampling day as a random factor), and each treatment combination was repeated five times over the experimental period (e.g. five times the combination no pools/5°, five times the combination no pools/15°, etc.).
Experimental apparatus and ramp hydraulics
A recirculating freshwater flow system (Rule 500 gph bilge pump) pumped water through a 25-mm diameter pipe from a lower holding tank (50 l) to an upper holding tank (20 l). From the upper holding tank, water spilled onto a ramp into a lower tank (40 l) in which fish were kept (A). Water in the lowest holding tank was constantly replaced with stream water from a local stream to avoid increases in water temperatures by, for instance, potential heat produced by the pump.
Fig. 1 A, Ramp profile, and B, cross-section (all dimensions are in mm), showing the experimental set-up used to test passage success of juvenile īnanga. Artificial grass was added to the ramp surface (C) used in testing juvenile īnanga passage success to provide a rough surface allowing fish to rest during their upstream ascent without getting washed back into the lower holding bin, and D, a pool (arrow) was created at 1.5 m to provide further resting opportunities for fish. E, An īnanga juvenile on the wetted margin of the ramp. In general, fish burst swam up the ramp for approximately 1 m, before they moved to the wetted margin of the ramp in our juvenile īnanga passage success tests.
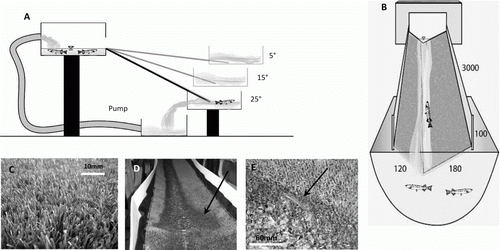
To ensure upstream passage was not restricted by water depth, mean water depth in the pools and in the centre of the ramp was kept at least twice the body depth of the largest juvenile fish tested (i.e. a minimum of 25 mm in pools and 15 mm on the ramp) by regulating flow conditions. The length of the ramp was 3500 mm with 500 mm immersed in water at the lower end so the actual ‘climbing’ length was 3000 mm. The ramp had a total width of 300 mm and was V-shaped in cross-section (B).
Water velocity (m/s) was measured using a flow meter (Flo-Mate Portable Velocity meter, Marsh-McBirney, Colorado, USA) at the centre of the ramp, 1.5 m below the outlet. Water velocities at these points were expected to be fastest. Water depth was measured with a ruler at the centre of the ramp, where water depth was expected to be deepest.
Water temperatures were measured at the beginning of each 3-h experiment with a handheld digital thermometer (0.2° accuracy). Water temperatures were not influenced by the heat produced by the pump; however, ambient air temperatures produced natural variations in water temperatures over the course of the day.
Artificial grass with a grass length of 10 mm (C) was used as ramp surface material. This material was expected to produce substratum characteristics conducive to fish ascending, as rough surfaces increase surface tension (Baker & Boubée Citation2006), enabling fish to rest. To provide further resting opportunities for fish during their ascent, pools were created by deforming the artificial grass surface from the underside with sand bags (D). Pool length (200 mm) and width (300 mm) at each angle was maintained by adjusting the height of the sand bags according to the steepness of the ramp.
Fish capture and maintenance
Juvenile īnanga (FL < 60 mm, E) were sourced from a small tributary of the Wakapuaka River at the Paremata Flats Reserve, Nelson. Fish were caught with a whitebait set-net (2-mm mesh net) and kept for up to 3 days in live boxes within a stream.
A maximum of 100 fish were transported in 20-l containers to the laboratory at least 24 h before commencement of each run of the experiment to allow fish to adjust to water temperatures before experiments started. In the laboratory, fish were kept in an aerated 30-l tank with rocks as cover for a maximum of 3 days, and fed each evening with chironomids. Tank water, which was piped from a local stream close to the laboratory and replaced daily, reflected temperatures found in the stream where fish were initially caught, ranging between 13° and 19°C over the experimental period (i.e. October–November). Daily maximum changes recorded during the course of the experiments were 2.0°C.
Fish that were used in experiments were released at the end of each day into the main stem of the Wakapuaka River, thus minimising the likelihood of recapture.
Statistical analyses
Before analyses, fish size and the number of fish to pass the ramp successfully were square-root-transformed where necessary to meet assumptions of normality and homogeneity of variances. Analysis of covariance (ANCOVA) conducted in Statistica 8.0 was used to evaluate if pool presence and ramp angle (5° and 15°) influenced water velocity. A two-sample t-test was used to test whether there was a significant difference in the number of fish passing the ramp at 15° compared with 5° slope.
To assess the effect of fish size on passing success at 5°, 15° and 25° ramp angles with one pool, a generalised mixed-effects (GME) model was used (using the lmer package in R; R Foundation Citation2007) with a binomial distribution (i.e. pass or fail). This allowed us to test the fixed effects of ramp angle (categorical variable) and fish size (continuous variable) on whether the fish were able to traverse the ramps while accounting for any variation that was related to running the experimental trials over different days (random block effect). Model simplification (Crawley Citation2007) was used with maximum likelihood (ML) estimation, where the fixed effects of ramp angle, fish size and their interaction were removed one-by-one from the model to estimate the chi-square statistic and its significance level for each of the fixed effects and their interaction on fish passing success.
A GME model (with normal distribution) was also used to determine whether resting opportunities (i.e. the presence or absence of a pool) influenced the number of fish (continuous response variable) that were successful in passing 5° and 15° ramp angles. Again, the main and interactive effects of ramp angle and pool presence/absence were tested using model simplification and ML (as above), after taking into account any variation related to sampling day (random block effect).
Results
Hydrodynamics of ramp and pools
Overall, water flow on the ramp was uniform, because of the consistent substratum texture. There was a statistically significant difference in water velocity, with slower velocities in the pools than on the ramp, and an increase in velocities with ramp angle (ANCOVA, F 1,3=27.27, P=0.01, A). However, there was a weak indication of an effect of pool presence on ramp water velocities (F 1,3=7.94, P=0.06). Water depth on the ramp and in pools also depended on ramp angle, with deeper water levels on lower ramp slopes ().
Fig. 2 A, Mean (±SE) velocity measured at 5°, 15° and 25° slope on the ramp (○) and in pools (•) in the experiments. B, Proportion (%) of juvenile īnanga passing the ramp at 5°, 15° and 25° angles with one pool.
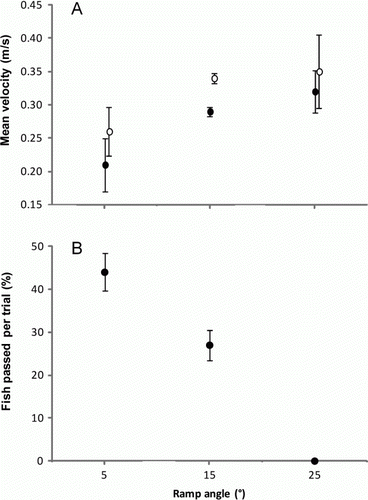
Table 1 The five treatment combinations tested in the ramp experiment evaluating juvenile īnanga passage success including three different angles with varying pool combinations
Fish behaviour
Once in the experimental channel, fish adjusted rapidly to their new environment, attempting to ascend after approximately 10 min. Fish continued to attempt upstream passage during the entire 3 h of a trial. To pass the ramp, fish would begin to swim in the main current at the base of the ramp before attempting an upstream ascent by burst swimming. Some fish burst swam up the ramp until they reached a pool or moved into the wetted margin at the sides of the ramp to rest before continuing their attempt (E). Others did not succeed in reaching a pool or get enough surface tension to rest on the wetted margin and were washed back into the lower holding tank.
The effect of angle on fish passage success
Fish were randomly selected across all trials and were variable in size, ranging from 40 to 58 mm. A two-sample t-test showed that fewer fish were able to successfully climb ramps with a 15° angle than a 5° angle, when a pool was present (t 8=2.9, P<0.05). As no fish passed at 25° with one pool (B), ANOVA could not be used because the homogeneity of variances assumption was violated. However, the 95% confidence interval for the percentage of fish passing at 15° (23–31% fish passed per trial) did not include zero, and so was significantly different from the number that did not pass at 25° (B).
The effects of fish size and pool presence on passage success
A binomial mixed-effects model indicated larger fish were able to surpass steeper angles (a; ), but there was no significant interaction between angle and fish size (a). To pass the ramp with a 50% probability, the analysis indicated fish needed to be larger than 49.6 mm at a 5° slope and larger than 54.6 mm at a 15° slope ().
Fig. 3 Predicted passing sizes of juvenile īnanga at 5° (—) and 15° (····) ramp angles with no pool determined by a logistic mixed-effects model.
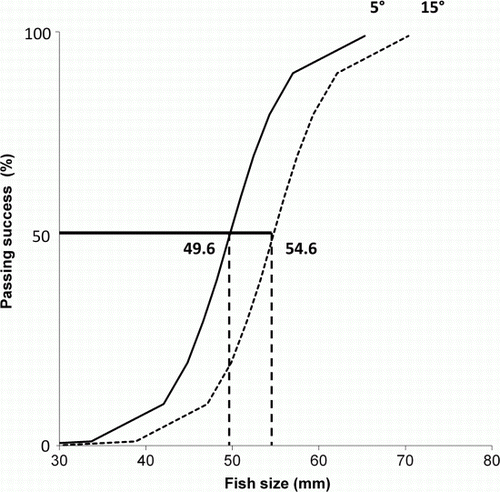
Table 2 Generalised mixed-effects models testing of the fixed main and interactive effects of (a) ramp angle (5°, 15°, 25°, one pool) and fish size on the success or failure of fish to climb the ramp, and (b) ramp angle (5° and 15°) and pool presence (no pool or one pool) on the success of failure of fish to climb the ramp, while accounting for variation because of the sampling day
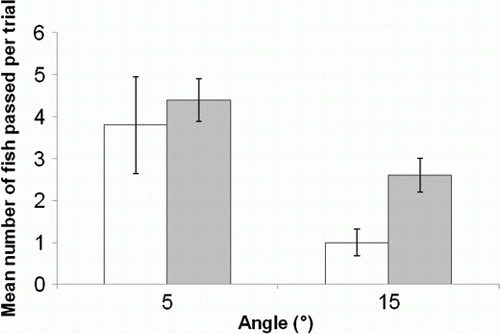
A linear GME indicated that pool presence did not significantly influence fish passage success, but fish were more likely to climb shallow ramps successfully than steep ramps (b; ). There was no significant interaction between ramp angle and pool presence (b). Although the presence of pools did not significantly improve upstream passage (P=0.08, b), there was an indication pools might be important because more fish ascended the 15° ramp on average when pools were present.
Fig. 4 Mean (±SE) number of juvenile īnanga that passed the ramp at 5° and 15° with zero pools (□) or one pool (▪).
Discussion
Fish size plays an important role in passage success, as larger fish are generally able to swim faster and for longer periods (Mitchell Citation1989; Peake Citation2004; Tudorache et al. Citation2008). However, previous studies have not considered the relationship between fish size and slope-related variables in their evaluation of fish passage design (Boubée et al. Citation2001; Knaepkens et al. Citation2006). Consistent with previous research on adult īnanga (Baker & Boubée Citation2006), the juveniles in this study needed to be larger to pass a steeper gradient. Juvenile īnanga in our study had to be on average 5 mm larger to surmount the ramp at 15° than those at 5°. This result highlights the importance of considering size and life stage of target fish species when constructing fish ramps (Nikora et al. Citation2003). When planning fish passage at culverts during, for instance, juvenile migration periods, fish ramp angle should be ideally 5° to provide successful passage for ‘non-climbing’ species or life stages.
Water velocities are likely to be one of the main parameters driving the relationship between fish size and passage success (Bouska & Paukert Citation2010). To design fish ramps successfully, water velocities on, within and downstream of the barrier need to be considered (Haro et al. Citation2004; House et al. Citation2005). The water velocity tested in this study increased with slope and only 26% of the juvenile īnanga tested were able to negotiate the 15° ramp successfully with one pool installed, compared with 44% at the 5° ramp. This result suggests that water velocities at ramps as shallow as 5° hinder more than half of the fish successfully negotiating the ramp. Therefore, angles steeper than 5° are likely to increase the velocity on fish ramps beyond the ability of weakly swimming fish species like juvenile īnanga, although this angle is still able to be surpassed over short distances (e.g. 25° over the 0.5-m ramp length; Baker & Boubée Citation2006; Tudorache et al. Citation2008).
If passing fish numbers are less than required to sustain a population, further considerations could include a biologically meaningful success rate. This includes the need to investigate how many fish need to pass in any 1 year to occupy the full extent of habitat available upstream and to sustain a population. Moreover, for velocity to be considered a barrier, it must exceed the physiological or behavioural capabilities of the target species and life stage of the fish that are expected to be migrating (Parker Citation2000; Stevenson et al. Citation2008). Increased water velocities can also cause changes in burst swim distances. Boubée et al. (Citation1999) showed the maximum burst swim distance for adult īnanga changed from 6.2 m at 0.35 m/s to 2.1 m at 1.0 m/s. These measurements were made for adult īnanga at a single channel angle. However, they illustrate how burst swim distances for juveniles could vary accordingly with fish ramp angle and hence water velocities.
Ramp length will also be an important factor when designing fish ramps and the length tested in this study simulated those required to access intermediate-sized obstacles such as small diameter culverts (diameter <1.5 m), enabling some individuals to successfully negotiate the full length of the ramp (Doehring et al. Citation2011a). However, Baker & Boubée (Citation2006) looked at juvenile īnanga (44–55 mm) climbing ability over a 1.5-m long ramp and found that 75% of fish successfully negotiated all ramps at a 15° angle, independent of surface substrate. This study showed that only 10% of fish passed at an angle of 15° over a ramp twice the length (3 m). Other factors influencing passage success may have differed between the two studies, but īnanga passage increased by approximately six times on a ramp half the length. Thus, the trade-off between ramp length and angle deserves further investigation.
In many cases, the height of a structure determines the length of a ramp, as the higher the structure, the longer the ramp must be if the angle should stay surmountable by ‘non-climbing’ fish species. Ramp length, in this case, may be a problem for fish if the distance they can travel at any one time is restricted to less than the full length of the ramp. The high energy costs involved during the upstream movement may mean fish become exhausted before they reach the end. In these situations, resting areas, such as pools, are recommended (Boubée et al. Citation1999).
Although the presence of pools had no statistically significant effect on the number of juvenile fish that passed the ramp at any angle in this study, the results suggest that pool presence could still be important. In our experiments fish attempted to ascend the entire 3 m without resting, suggesting fish may not have been exhausted enough after a 1.5-m ascent at a 25° slope actually to use the pool to rest. If pools exist at appropriate distances along a ramp (informed by the distance at which a fish fatigues for a given ramp angle), fish might potentially be more inclined to use pools rather than the edge of the ramp. However, until further data have been gathered on the effects of resting opportunities, fish ramp designs should provide at least one wetted margin with rough surface substrata to enable fish to rest during their upstream movement.
Suitable surface substrate may also offer sufficient resting opportunities for fish during their climb and likely affected passage in this study. The artificial grass used in this study provided suitable surface tension for fish to rest; however, the durability of this material in the field needs further testing. Surface substrata in general should be as rough as possible to both reduce flow and widen the wetted margin, offering fish opportunities to rest (Hegberg et al. Citation2001; Haro et al. Citation2004).
Before installing fish ramps in the field, ramp length, angle and life stage of the target fish species need to be considered. This study showed that for weak swimming species, ramps should, if possible, have a maximum angle of 5° over a 3-m length, with rough surface materials to reduce water velocities and increase surface tension. Although the installation of pools did not significantly improve upstream passage in this study, further research could indicate they may improve passage at ramps longer than 3 m. Future experiments could also incorporate juveniles of other migratory species, thereby providing valuable information for the development of optimal passage solutions for target species (MacDonald & Davies Citation2007). Finally, passage designs should be orientated toward more than just one target species to provide passage for a wide variety of fish species. However, within the group of target species, the weakest swimming species should set the passage requirements if those species are expected to occur above the structure (Boubée et al. Citation1999). Īnanga is considered one of the weakest swimming species in New Zealand (Mitchell Citation1989; Nikora et al. Citation2003; Plew et al. Citation2007). Thus, our findings can, and should be, used as a basis for passage design, improving upstream passage of a variety of fish species. This might eventually allow upstream fish diversity to expand and fish populations to be sustained in urban waterways.
Acknowledgements
This study was initiated with the encouragement and part funding of Nelson City Council, and an ENVIROLINK small advice grant. We thank the University of Canterbury for Animal Ethics approvals, and the Ministry of Fishery and the Department of Conservation for fish transfer permits. We also thank the Cawthron Institute, the Department of Conservation and the Freshwater Ecology Research Group at the University of Canterbury for providing research material and field assistance, as well as Bruno David, William Davidson and two anonymous reviewers for comments on the manuscript.
References
- Allibone , RM . 1999 . Impoundment and introductions: their impacts on native fish of the upper Waiapori River, New Zealand . Journal of the Royal Society of New Zealand , 29 : 291 – 299 .
- Baker , CF and Boubée , JAT . 2006 . Upstream passage of īnanga (Galaxias maculatus) and redfin bullies (Gobiomorphus huttoni) over artificial ramps . Journal of Fish Biology , 69 : 668 – 681 .
- Boubée JA, Jowett IG, Nichols S, Williams E 1999 . Fish passage at culverts: a review, with possible solutions for New Zealand indigenous species . NIWA report for the Department of Conservation .
- Boubée , JA , Mitchell , CP , Chisnall , BL , West , DW , Bowman , EJ and Haro , A . 2001 . Factors regulating the downstream migration of mature eels (Anguilla spp.) at Aniwhenua Dam, Bay of Plenty, New Zealand . New Zealand Journal of Marine and Freshwater Research , 35 : 121 – 134 .
- Bouska , WW and Paukert , CP . 2010 . Road crossing designs and their impact on fish assemblages of Great Plains streams . Transactions of the American Fisheries Society , 139 : 214 – 222 .
- Collier , KJ , Aldridge , BMTA , Hicks , BJ , Kelly , J , Macdonald , A , Smith , BJ and Tonkin , J . 2009 . Ecological values of Hamilton urban streams (North Island, New Zealand): constraints and opportunities for restoration . New Zealand Journal of Ecology , 33 : 177 – 189 .
- Crawley MJ 2007 . The R book . Chichester , Wiley .
- David , BO , Hamer , MP and Collier , KJ . 2009 . Mussel spat ropes provide passage for banded kokopu (Galaxias fasciatus) in laboratory trials . New Zealand Journal of Marine and Freshwater Research , 43 : 883 – 888 .
- Doehring K 2009 . Urbanisation influences on freshwater fish distribution and remediation of migratory barriers . Unpublished Master of Science thesis , University of Canterbury , , Christchurch .
- Doehring , K , McIntosh , AR and Young , RG . 2011a . Factors affecting juvenile galaxiid fish passage at culverts . Marine and Freshwater Research , 62 : 38 – 45 .
- Doehring , K , Young , RG , Hay , J and Quarterman , AJ . 2011b . Suitability of Dual-frequency Identification Sonar (DIDSON) to monitor juvenile fish movement at floodgates . New Zealand Journal of Marine and Freshwater Research , 45 : 413 – 422 .
- Eikaas , HS and McIntosh , AR . 2006 . Habitat loss through disruption of constrained dispersal networks . Ecological Applications , 16 : 987 – 998 .
- Gibson , RJ , Haedrich , RL and Wernerheirn , CM . 2005 . Loss of fish habitat as a consequence of inappropriately constructed stream crossings . Fisheries , 30 : 10 – 17 .
- Hammer , C . 1995 . Fatigue and exercise tests with fish . Comparative Biochemistry and Physiology , 112 : 1 – 20 .
- Haro , A , Castro-Santos , T , Noreika , J and Odeh , M . 2004 . Swimming performance of upstream migrant fishes in open-channel flow: a new approach to predicting passage through velocity barriers . Canadian Journal of Fisheries and Aquatic Sciences , 61 : 1590 – 1601 .
- Hegberg CH, Jacobs S, Schlindwein PA, Cohen SJ 2001 . Fish Passage in urban streams (Part 1: Regulatory and resource issues) . Proceedings of the 2001 Wetlands Engineering and River Restoration Conference, Reno, Nevada .
- Hickford , MJH , Cagnon , M and Schiel , DR . 2010 . Predation, vegetation and habitat-specific survival of terrestrial eggs of a diadromous fish, Galaxias maculatus (Jenyns, 1842) . Journal of Experimental Marine Biology and Ecology , 385 : 66 – 72 .
- House , MR , Pyles , MR and White , D . 2005 . Velocity distributions in streambed simulation culverts used for fish passage . Journal of the American Water Resources Association , 41 : 209 – 217 .
- Jowett , IG . 2002 . In-stream habitat suitability criteria for feeding īnanga (Galaxias maculatus) . New Zealand Journal of Marine and Freshwater Research , 36 : 399 – 407 .
- Joy , MK , Henderson , IM and Death , RG . 2000 . Diadromy and longitudinal patterns of upstream penetration of freshwater fish in Taranaki, New Zealand . New Zealand Journal of Marine and Freshwater Research , 34 : 531 – 543 .
- Kemp , PS and Williams , JG . 2008 . Response of migrating Chinook salmon (Oncorhynchus tshawytscha) smolts to in-stream structure associated with culverts . River Research and Applications , 24 : 571 – 579 .
- Kemp , PS and O'Hanley , JR . 2010 . Procedures for evaluating and prioritising the removal of fish passage barriers: a synthesis . Fisheries Management and Ecology , 17 : 297 – 322 .
- Knaepkens , G , Baekelandt , K and Eens , M . 2006 . Fish pass effectiveness for bullhead (Cottus gobio), perch (Perca fluviatilis) and roach (Rutilus rutilus) in a regulated lowland river . Ecology of Freshwater Fish , 15 : 20 – 29 .
- Leathwick , JR , Elith , J , Chadderton , WL , Rowe , D and Hastie , T . 2008 . Dispersal, disturbance and the contrasting biogeographies of New Zealand's diadromous and non-diadromous fish species . Journal of Biogeography , 35 : 1481 – 1497 .
- MacDonald , JI and Davies , PE . 2007 . Improving the upstream passage of two galaxiid fish species through a pipe culvert . Fisheries Management and Ecology , 14 : 221 – 230 .
- Magaud , H , Migeon , B , Morfin , P , Garric , J and Vindimian , E . 1997 . Modelling fish mortality due to urban storm run-off: interacting effects of hypoxia and un-ionized ammonia . Water Research , 31 : 211 – 218 .
- Mardones , A , Vega , R and Encina , F . 2008 . Cultivation of whitebait (Galaxias maculatus) in Chile . Aquaculture Research , 39 : 731 – 737 .
- McDowall , RM . 1993 . Implications of diadromy for the structuring and modelling of riverine fish communities in New Zealand . New Zealand Journal of Marine and Freshwater Research , 27 : 453 – 462 .
- McDowall , RM . 1998 . Fighting the flow: downstream-upstream linkages in the ecology of diadromous fish faunas in West Coast New Zealand rivers . Freshwater Biology , 40 : 111 – 122 .
- McDowall RM 2000 . The Reed Field Guide to New Zealand Freshwater Fishes . Reed Publishing (NZ) Ltd , Auckland .
- McDowall RM 2001 . Freshwater Fishes of New Zealand . Auckland , Reed Publishing (NZ) Ltd .
- McDowall , RM . 2003 . The key to climbing in the koaro . NIWA Water & Atmosphere , 11 : 16 – 17 .
- Mitchell , CP . 1989 . Swimming performances of some native freshwater fishes . New Zealand Journal of Marine and Freshwater Research , 23 : 181 – 187 .
- Naughton , GP , Caudill , CC , Peery , CA , Clabough , TS , Jepson , MA , Bjornn , TC and Stuehrenberg , LC . 2007 . Experimental evaluation of fishway modifications on the passage behaviour of adult Chinook salmon and steelhead at Lower Granite Dam, Snake River, USA . River Research and Applications , 23 : 99 – 111 .
- Nikora , VI , Aberle , J , Biggs , BJF , Jowett , IG and Sykes , JRE . 2003 . Effects of fish size, time-to-fatigue and turbulence on swimming performance: a case study of Galaxias maculatus . Journal of Fish Biology , 63 : 1365 – 1382 .
- Parker , MA . 2000 . Fish passage—culvert inspection procedures . Watershed Restoration Technical Circular , 11 : 1 – 47 .
- Peake , S . 2004 . An evaluation of the use of critical swimming speed for determination of culvert water velocity criteria for smallmouth bass . Transactions of the American Fisheries Society , 133 : 1472 – 1479 .
- Peake , S , McKinley , RS and Scruton , DA . 1997a . Swimming performance of various freshwater Newfoundland salmonids relative to habitat selection and fishway design . Journal of Fish Biology , 51 : 710 – 723 .
- Peake , SJ , Beamish , FWH , McKinley , RS , Scruton , DA and Katapodis , C . 1997b . Relating swimming performance of lake sturgeon, Acipenser fulvescens, to fishway design . Canadian Journal of Fisheries and Aquatic Sciences , 54 : 1361 – 1366 .
- Plew , DR , Nikora , VI , Larned , ST , Sykes , JRE and Cooper , GG . 2007 . Fish swimming speed variability at constant flow: Galaxias maculatus . New Zealand Journal of Marine and Freshwater Research , 41 : 185 – 195 .
- Poplar-Jeffers , IO , Petty , JT , Anderson , JT , Kite , SJ , Strager , MP and Fortney , RH . 2009 . Culvert replacement and stream habitat restoration: implications from Brook trout management in an Appalachian Watershed, U.S.A . Restoration Ecology , 17 : 404 – 413 .
- R Foundation 2007 . The R Foundation for statistical computing 2007 , R 2.6. http://www.r-project.org (accessed 19 February 2009).
- Richmond , MC , Deng , Z , Guensch , GR , Tritico , H and Pearson , WH . 2007 . Mean flow and turbulence characteristics of a full-scale spiral corrugated culvert with implications for fish passage . Ecological Engineering , 30 : 333 – 340 .
- Rowe , DK , Saxton , BA and Stancliff , AG . 1992 . Species composition of whitebait (Galaxiidae) fisheries in twelve Bay of Plenty rivers, New Zealand: evidence for river mouth selection by juvenile Galaxias brevipinnis (Günther) . New Zealand Journal of Marine and Freshwater Research , 26 : 219 – 228 .
- Rowe , DK , Smith , J and Williams , E . 2002 . Effects of turbidity on the feeding ability of adult, riverine smelt (Retropinna retropinna) and īnanga (Galaxias maculatus) . New Zealand Journal of Marine and Freshwater Research , 36 : 143 – 150 .
- Stancliff AG, Boubée JA, Palmer D, Mitchell CP 1988 . The upstream migration of whitebait species in the Lower Waikato River. New Zealand Freshwater Fisheries Report No. 96. MAFFish, Rotorua, 96 .
- Stevenson C, Kopeinig T, Feurich R, Boubée J 2008 . Culvert barrel design to facilitate the upstream passage of small fish . A report for the Auckland Regional Council. NIWA Client Report: HAM2008-046 .
- Tudorache , C , Viaene , P , Blust , R , Vereecken , H and De Boeck , G . 2008 . A comparison of swimming capacity and energy use in seven European freshwater fish species . Ecology of Freshwater Fish , 17 : 284 – 291 .
- Weihs , D . 1974 . Energetic advantages of burst swimming of fish . Journal of Theoretical Biology , 48 : 215 – 229 .