Abstract
Periphyton standing crop is often measured as chlorophyll a but there is increasing interest in using visual assessments of periphyton cover. Visual methods are rapid and require no sample analysis, but can be regarded as subjective with high inter-operator variability. We investigated variability in periphyton standing crop across operators, rivers and time, as determined visually and from chlorophyll a measurements. We found that visual assessments (comprising percentage cover estimates of up to eight periphyton categories) distinguished sites and occasions as effectively as chlorophyll a. Furthermore, an estimate of chlorophyll a could be derived from the visual assessments. Because our surveys were conducted in only three rivers, general applicability of the derivation of estimated chlorophyll a from the visual assessments warrants further investigation. The current recommendation of 20 views was sufficient for realistic visual assessments of average cover. Overall, our results indicated that inter-operator variability in visual assessments need not be a major concern, given adequate training.
Introduction
Increasing pressure on river water resources in New Zealand, along with growing land use intensification, has highlighted concerns about effects on river ecosystems (New Zealand Conservation Authority Citation2011). Ecosystem responses to changes in, for example, flow regime, nutrient and sediment inputs, and riparian shading, are often most visible in primary production, especially periphyton (benthic algae). Changes have been documented in both the biomass and community composition of periphyton following installation of upstream impoundments (e.g. Benenati et al. Citation2000; Flinders & Hart Citation2009; Wu et al. Citation2009; Lessard et al. Citation2013) and in association with nutrient changes (e.g. Chétalat et al. Citation1999; Dodds et al. Citation2002; Black et al. Citation2011). Such changes have implications for higher trophic levels (Bourassa & Cattaneo Citation2000; Suren et al. Citation2003) and for human and animal health (Heath et al. Citation2011). Understanding linkages between primary production and flow characteristics, nutrient concentrations and other catchment changes is now critical in New Zealand because regional authorities are obliged to set limits to protect river ecosystems and other values (Ministry for the Environment Citation2011). Advances in quantifying such linkages were made during the 1980s and 1990s (Biggs & Close Citation1989; Biggs Citation1996), including the development of preliminary empirical relationships applicable to gravel-bed rivers, linking nutrient concentrations, accrual times (time between floods) and periphyton biomass measured as chlorophyll a (Biggs Citation2000a). Further development of the relationships was recommended to improve applicability within specific regions and river types (Biggs Citation2000a).
Chlorophyll a has long been the standard method for estimating stream periphyton biomass (e.g. Biggs & Kilroy Citation2000 and references therein; Dodds et al. Citation2002; Hambrook et al. Citation2007) because all types of algae contain chlorophyll a and the metric is assumed to reflect the total amount of live algae in a sample. Estimates of mean chlorophyll a are obtained by quantitative sampling of periphyton at multiple locations in the reach of interest (to encompass spatial variability) followed by laboratory analyses on individual or pooled samples, all of which represents significant field and laboratory effort. Thus, the resources required for the extensive surveys necessary to refine existing primary production–environment relationships and for subsequent biological monitoring, may be prohibitive. Consequently, there is increasing interest in New Zealand in using visual assessments to monitor total periphyton standing crop (Biggs Citation2000b). These visual assessments take the form of estimating percentage cover of different types of periphyton. Methods for such detailed surveys exist but have yet to be rigorously tested for consistency and variability (e.g. RAM-2; Biggs & Kilroy Citation2000).
Visual assessments have several advantages over chlorophyll a and other laboratory measurements of biomass, such as ash-free dry mass. They provide a direct estimate of both the degree of coverage and the type of periphyton at a river site. Percentage cover estimates require no interpretation and are readily understood by the public. Visual assessments are cost-efficient because they can be carried out rapidly and produce immediate results. A perceived disadvantage is that visual estimates can be regarded as subjective, only semi-quantitative (Stevenson & Bahls Citation1999), and prone to high inter-operator bias and variability. Therefore, the use of visual assessments for setting and monitoring standards may be viewed as unreliable compared to quantitative laboratory measurements of chlorophyll a. A further disadvantage of visual assessments of periphyton cover is that the output is generally multivariate, which is more difficult to interpret or use as a standard or guideline than a single value representing biomass. However, estimates of chlorophyll a are also subject to variability attributable to inter-operator differences. For example, bias and/or variability may arise through choice of substrata, location sampled (if removing a defined area of periphyton), estimates of rock surface area (if sampling whole rocks; Bergey & Getty Citation2006), and intensity of scraping or brushing of the sampled area.
In the present study, our objective was to compare the ability of visual estimates and chloophyll a measurements to detect variability in periphyton standing crop among rivers and over time whilst taking into account operator variability. We tested the hypothesis that inter-operator variability associated with estimating periphyton status using visual assessments would be no greater than that associated with estimates of mean chlorophyll a using quantitative sampling. Support for this hypothesis would imply that visual assessments are capable of detecting changes in periphyton biomass between rivers and over time as effectively as chlorophyll a.
In addition we used our analysis to:
| 1. | confirm optimum numbers of views or samples required to obtain an acceptable estimate of mean periphyton coverage and chlorophyll a within a reach; | ||||
| 2. | develop a method for reducing visual estimates to a single index by determining a chlorophyll a equivalent; and | ||||
| 3. | investigate sources of potential bias in chlorophyll a measurements, in particular the effect of non-random selection of rocks. | ||||
We expected that (3) might assist in understanding any discrepancies between observed biomass (measured as chlorophyll a) and estimated biomass (chlorophyll a equivalent derived from visual estimates). Ultimately we wished to provide guidance on the suitability of visual estimates of periphyton cover as a basis for decision-making, such as setting flow and nutrient limits to protect ecological and other values in rivers.
Methods
Site locations
The surveys were conducted in three rivers in lowland Canterbury, New Zealand, with contrasting water chemistry and catchment characteristics (). The Selwyn River is hill-fed with a significant proportion of its catchment in native forest or other native vegetation (e.g. tussock grassland). The study reach was shaded by trees and had variable periphyton cover including red and green filamentous algae (Rhodophyta, Chlorophyta and Xanthophyceae [Ochrophyta]), cyanobacterial mats and diatom-dominated mats. The Cust River is a spring-fed, channelised stream draining intensively farmed pasture and residential smallholdings. The study reach was partly shaded. Despite high nutrient concentrations, periphyton standing crop was generally much lower and more uniform than in the Selwyn River. The Waipara River is a larger hill-fed catchment again largely developed for agriculture. The catchment is distinctive in this part of Canterbury because of a high proportion of limestone geology. The surveyed reach was unshaded and periphyton standing crop varied considerably over both space and time.
Table 1 Locations and characteristics of the three study sites on the Cust, Selwyn and Waipara Rivers, Canterbury, New Zealand.
We restricted surveys to a single habitat type (runs), which is recommended for surveys aimed at detecting changes over time (Stevenson & Bahls Citation1999). Each survey reach included at least 30 m of run habitat with water depth of 0.6 m or less. Surveys were conducted in each river in April, June and September 2011.
Field procedures
On each sampling occasion at each site, a 30 m length of run habitat was marked out into 12 transects approximately 3 m apart. Ten points were marked at even spacing along each of the 12 transects by placing a painted rock on the river bed. These rocks defined 120 survey areas for visual assessments and for collection of rocks for quantitative sampling of periphyton.
Visual assessments were conducted using a system of categories developed in conjunction with end-users of the techniques in New Zealand (e.g. Kilroy et al. Citation2008; ). The aim of the system was to minimise the number of periphyton categories included, but as far as possible to separate cover that represented different concentrations of chlorophyll a and/or was of special interest to river managers (e.g. potentially toxic Phormidium mats). Three observers independently assessed visual cover of periphyton at each defined survey point using circular underwater viewers with a diameter of 350 mm (Nuova Rade, Genova, Italy). All observers viewed the same areas by locating the painted rock just out of view on the left hand side of the viewed circle. The bottom of the underwater viewer was divided into quarters to facilitate and standardise estimates of percentage cover. The three observers had differing levels of experience in visual assessments, from very experienced to inexperienced. Training was given to all observers at the start of the study, to ensure that the categories were understood.
Table 2 Categories of periphyton distinguished in the visual assessments.
Following the visual assessments, four transects were randomly assigned to each observer for collection of samples for chlorophyll a analysis (40 samples per observer). A single rock, or a sample of smaller particles, was randomly retrieved from within each pre-defined viewing circle. Before sample collection, periphyton cover on the rock, and x, y and z dimensions, were recorded. For rocks larger than about 60 mm diameter, a plastic lid (55 mm diameter) was placed on the upper rock surface to define the sampled area and all periphyton not covered by the lid was removed. Periphyton was then transferred from the defined area into a sample container with minimal river water using a combination of a sharp blade and small brushes with stiff bristles. For smaller particles, the lid was inverted over the substratum and the collection of smaller particles filling the lid was retrieved and scraped/brushed. Samples were kept cool and in the dark, returned to the laboratory within 5 hours of collection and frozen until analysis. To determine whether there was bias in sampled rock sizes, a separate random collection was made by a fourth person of 10 rocks on each of 12 transects in between the marked transects, and the x, y and z dimensions measured. These rocks were sampled using the standard Wolman methodology (Wolman Citation1954).
Laboratory analyses
Thawed samples were blended to a homogeneous slurry and made up to a known volume. Measured subsamples were filtered through glass fibre filters (Whatman GF/C, GE Healthcare Life Sciences, Fairfield, CT), from which chlorophyll a was extracted in boiling 95% ethanol for 5 minutes. Chlorophyll a concentration was determined from readings of absorbance at 665 nm (with correction for phaeophytins) after overnight incubation at 4°C, following the method in Biggs and Kilroy (Citation2000).
Data analysis
Differences in visual estimates among sites, occasions and observers
Mean percentage cover for all survey combinations (i.e. combinations of river, survey occasion, observer and periphyton category) was summarised in bar graphs. Two datasets of the visual observations were used: 1) all 120 visual assessments made by each observer at each site on each occasion (i.e. observations viewed at the same locations by different observers); and 2) the same 120 assessments divided into three sets of 40 visual assessments made by each of the three observers at each site at locations that were also sampled for chlorophyll a (i.e. assessments at different locations for each observer). We performed 3-way ANOVAs, with observer, site and occasion as factors, on each visual assessment category (for each of the two datasets) and on chlorophyll a (for 40 samples). For visual assessments, we used mean percentages on transects because the high proportion of zeroes in the raw data precluded satisfactory transformation to achieve normality and homoscedasticity of data. A lack of significant interactions among observers across sites and survey occasions would indicate consistency of assessments among observers, especially if there were also significant differences between sites and over time.
Optimum sample size
We used bootstrapping to determine the optimum sample size for visual assessment observations for each survey combination. Mean percentage cover by each visual category was calculated 100 times from n randomly resampled visual observations (with replacement), where n was 2 to 120 in steps of 2. Boxplots were generated from these bootstrap estimates of the means to demonstrate the relationship between numbers of samples and resampled mean percentage cover for each survey combination. For every set of 100 resampled combinations we calculated the difference between the bootstrap estimates and the observed mean of all 120 samples. We assumed that an acceptable level of variability was for 95% of the bootstrap estimates to be within 10% of the observed mean and determined the lowest numbers of samples required to meet this condition. A similar procedure was conducted for chlorophyll a except that we calculated the proportion of the bootstrap estimates that were within 20% of the observed mean. The higher percentage was used because chlorophyll a is an open-ended metric that can reach 500 mg m−2 or more, and±10% is unlikely to be ecologically significant. As an alternative approach we calculated the proportion of bootstrap estimates that were above established thresholds of chlorophyll a for maintenance of a range of instream values. The thresholds were 15, 50, 120 and 200 mg m−2, for protection of benthic biodiversity (mean and maximum), aesthetics/recreation and trout habitat/angling values (Biggs Citation2000b), respectively.
Estimating chlorophyll a from visual assessments
The mean chlorophyll a equivalent of each visual category was determined as the geometric mean of chlorophyll a determined from samples taken from rocks that were covered by a single periphyton category. Mean assessments of percentage cover on the river bed in each survey combination were then converted to a derived chlorophyll a using these empirical conversion factors. We generated derived chlorophyll afor each survey combination using 10, 20 and 40 views of the river bed (i.e. on 1, 2 and 4 transects). Observer-specific and overall relationships between mean derived chlorophyll a and observed chlorophyll a were determined using linear regression on log10-transformed data. We also performed 3-way ANOVAs, as above, with derived chlorophyll a (log-transformed) as the dependent variable, to determine differences across observers, sites and occasions.
Bias in rock selection and effect on chlorophyll a
An estimate of rock size (exposed surface area) was calculated from the x, y and z dimensions (in cm) by applying the equation in Biggs and Kilroy (Citation2000):
Differences among observers (including the fourth person who made an independent random rock collection) and survey occasions were determined using 2-way ANOVA at each site with log10-transformed data. For each survey combination we used linear regression to assess the relationship between observed chlorophyll a per unit area and rock size.
Results
General observations
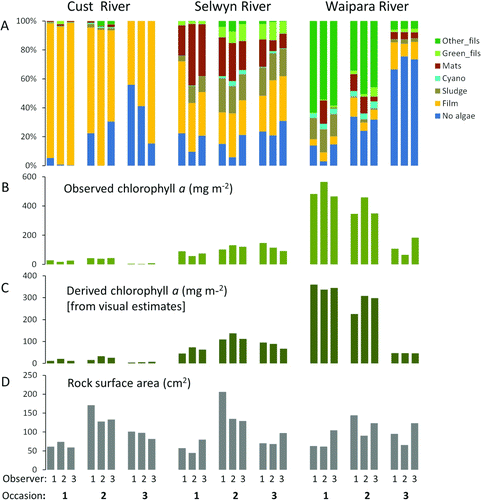
Periphyton cover differed at the three sites and also changed over time. Mean cover by different categories of periphyton (calculated from all 120 views and from the 120 views divided into three sets of 40 views, each assessed independently by one observer) was generally characteristic of each site and each survey (A), with the overlapping and independent assessments very similar to each other. The main discrepancies among observers appeared to be in estimates of No algae, Film and Mats. No periphyton in the Didymo category was observed during any of the surveys.
Figure 1 Summary of measurements made by three observers at three rivers sites on three occasions. A, Estimates of mean periphyton cover in seven visual categories with each observer viewing 40 different points. B, Mean chlorophyll a determined from quantitative samples collected from the same 40 points viewed in A. C, Mean chlorophyll a derived from visual assessments of percentage cover weighted by a conversion factor (see text for details). D, Mean size of rocks sampled for chlorophyll a, as surface area estimated from x, y and z dimensions.
Variability and optimum sample size for visual assessments and chlorophyll a
Differences in periphyton over time and among sites were greater than differences among observers. Using either the full dataset of 120 points, or the three sets of 40 points independently assessed by each observer, two of the seven categories (No algae and Mats) differed across observers (). In contrast, all cover categories differed among sites and five of the seven differed over time. Significant interactions for survey×observer and site×observer for the categories No algae, Film and Mats confirmed that observers were not recording these categories consistently.
Table 3 Summary results (P-values) of 3-way ANOVAs to determine differences in estimates of % cover of seven visual categories of periphyton, across observers, survey occasions and sites, and showing interactions between observers and survey occasions and sites. (a) Data from a 120-point dataset (all observers viewed the same locations); (b) data from a 40-point dataset (observers all viewed different locations). Data were mean percentage cover estimates along transects of 10 views. Significant differences are in bold type.
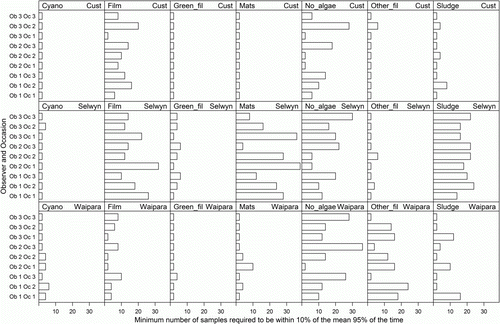
Variability in estimates of mean percentage cover by each visual periphyton category declined as sample number increased (e.g. ), with a rapid decline with increasing sample numbers up to 10. The maximum number of samples required to achieve 95% confidence of being within 10% of the observed mean from 120 samples was less than 20 for 89% of the 189 survey combinations (). Twenty survey combinations required more than 20 samples to achieve a 95% confidence of being within 10% of the true mean, including representatives from all three sites (Cust, 2; Selwyn, 14; Waipara, 4) and from five of the seven categories (all except Cyano and Green_fil). For one of these combinations (Mats in the Selwyn on occasion 1 by observer 2) 95% of bootstrap estimates did not fall within 10% of the observed mean even when n was 120. This indicates that there was high variability within these 120 samples. For the remaining 19 combinations, up to 40 samples were required to achieve a 95% confidence of being within 10% of the observed mean.
Figure 2 Cover by mats at the Selwyn River site in June 2011 (visit 2) assessed independently by three observers at 120 pre-determined viewing locations. Each box plot is derived from 100 random selections (with replacement) of 2, 4, 6, etc. observations (views).
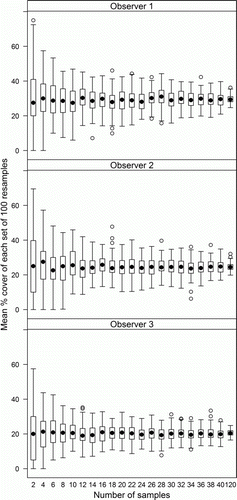
Figure 3 Minimum numbers of views required to achieve 95% confidence of being within 10% of the true mean, calculated for every survey combination and visual assessment category using bootstrap resampling on the 120 views assessed by each observer in each survey.
Mean observed chlorophyll a (B) differed among surveys and sites, but not observers, and the interaction terms were also non-significant (). The number of samples required for at least 95% of bootstrap estimates to be within 20% of the observed mean for 40 samples was < 20 for only three survey combinations (read off ). In 11 of the 27 survey combinations (41%), the proportion of bootstrap estimates within 20% of the observed mean was less than 80% even with 40 samples. Chlorophyll a was pooled across observers for the threshold analysis. For some sites, on some occasions, nearly all chlorophyll a samples were either less than the given threshold or greater than the given threshold, and therefore very few samples would be required to confidently establish that mean chlorophyll a was either below or above that threshold (). For example, the Cust on occasion 3 had very low chlorophyll a; therefore nearly all bootstrap estimates had means less than 15 mg m−2 even when the number of samples was two. At the other extreme, the Waipara on occasions 1 and 2 had very high chlorophyll a; therefore nearly all bootstrap estimates were greater than 200 mg m−2 even when the number of samples was low. This meant that for 21 of the 36 (58%) site–occasion–threshold combinations, five or fewer samples were required to be 95% confident that the mean was either above the threshold or below the threshold and for a further 11 combinations, 25 or fewer samples were required. However, when the observed mean chlorophyll a was near to the given threshold approximately half the bootstrap estimates had means that were greater than the threshold regardless of number of samples. This meant that for four of the 36 (31%) survey combinations, one could not be 95% confident that the mean was either above the given threshold or below the given threshold even with 120 samples.
Figure 4 Proportion of the means of 100 randomly re-sampled sets of observations of chlorophyll a (for each site, occasions and observer combination), which were within 20% of the mean of all 120 observations. Numbers of samples in each resampled set ranged from two to 40. Each row shows the results from one observer.
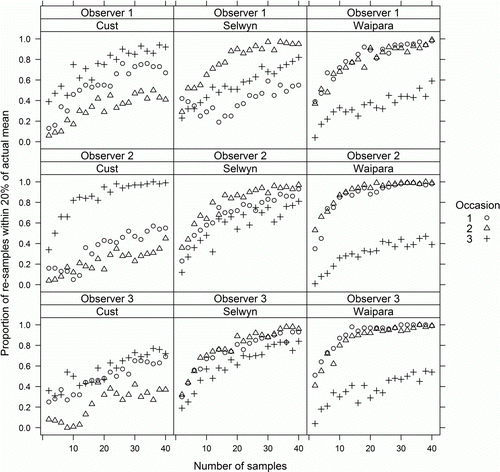
Figure 5 Proportion of the means of 100 randomly resampled sets of observations of chlorophyll a (with data from three observers combined), which were greater than 15, 50, 120 and 200 mg m−2 (bottom row to top row). Numbers of samples in each resampled set ranged from two to 40.
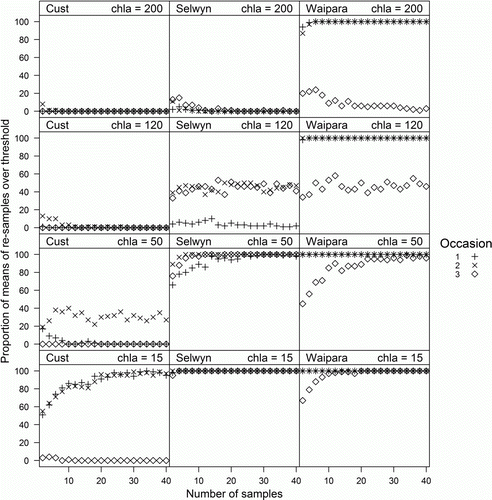
Table 4 Summary results (P-values) of 3-way ANOVAs to determine differences in observed and derived chlorophyll a across observers, survey occasions and sites. Chlorophyll a was estimated from the visual assessments using conversion factors determined from cover on individual rocks (see ). Significant differences are in bold type.
Chlorophyll a derived from visual assessments
At least seven rocks were available for calculation of a mean chlorophyll a equivalent (conversion factor) for each visual assessment category (range 7 [Cyano] to 218 [Film]). Mean chlorophyll a equivalents varied from 0.75 mg m−2 (No algae) to 599 mg m−2 (Cyano) (). The chlorophyll a equivalent, derived from the visual assessments using the conversion factors, was strongly correlated with observed chlorophyll a (), with only slight variations among observers (data not shown). The strongest correlation was achieved using mean values calculated from all 40 viewing points (R2=0.902, P<0.001) but using just 20 points (on two random transects) produced relationships only marginally weaker (). Derived chlorophyll a (C) was lower than observed (paired t-test, P<0.001) and was on average 71% of observed chlorophyll a. Derived chlorophyll a differed among sites and occasions but not among observers (). Non-significant interactions between observers and sites and occasions indicated that the estimates were consistent. This result held for derived values from all 40 views from which samples for chlorophyll a were collected (i.e. a different set of 40 views for each observer), and for random sets of lower numbers of views (e.g. n=12, ). Variability among observers (as the coefficient of variation) in derived chlorophyll a was similar to that for observed chlorophyll a across all sites and surveys (paired t-test, P=0.399).
Figure 6 Chlorophyll a derived from visual estimates made by three observers at three sites on three occasions, using the conversion factors in , plotted against measured chlorophyll a. Each point is the mean of chlorophyll a samples and visual assessments at 20 points. The solid line indicates the best fit regression (R2=0.889, P<0.001) and the dashed line is the 1:1 relationship.
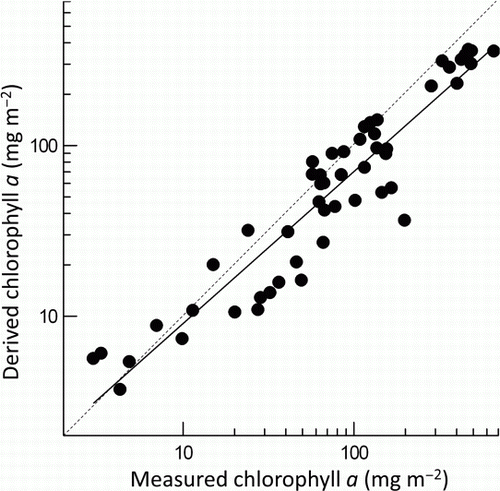
Table 5 Chlorophyll a equivalents for the seven periphyton categories, determined from measurements of chlorophyll a from samples with 100% cover by each category.
Table 6 Summary results (P-values) of pairwise comparisons (following significant ANOVAs) of mean rock size in collections made by three observers (obs 1, 2 and 3) for chlorophyll a samples, and by a fourth person as a separate random collection (random). Significant differences are in bold type.
Bias in rock selection and effect on observed chlorophyll a
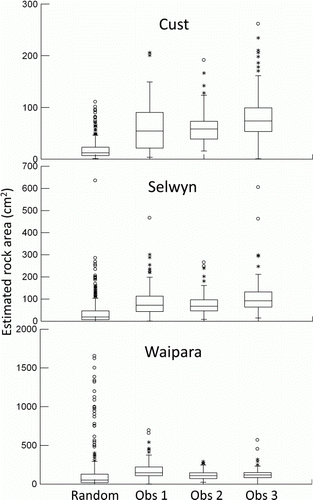
The mean size of rocks sampled for chlorophyll a (D) was larger, on average, than that in the random collection at each site (, ). Observed chlorophyll a was weakly and positively related to rock size using the combined data from all observers, sites and occasions (r = 0.33, P<0.001).
Figure 7 Distributions of rock sizes (as estimated exposed surface area calculated from x, y and z dimensions) collected at the Cust, Selwyn and Waipara River survey sites by three observers for sample collection for chlorophyll a, plus a random collection made by a fourth person. Total n=120 for observers 1, 2 and 3, and n = 360 for the random collection, pooled from three survey occasions.
Discussion
Visual assessment-based methods for estimating cover by various organisms have been used extensively in terrestrial studies (e.g. the Braun Blanquet scale, Wikum & Shanholtzer Citation1978), and in studies of aquatic macrophytes (e.g. Champion & Tanner Citation2000; Wood et al. Citation2012). However, use of visual estimates for monitoring stream periphyton appears to be relatively unusual compared to the widespread use of chlorophyll a as a measure of standing crop (but see Bowden et al. Citation1994; Necchi et al. Citation1995; Arscott et al. Citation1998; Benstead et al. Citation2007; Rodriguez et al. Citation2007). In the present study, a derived estimate of chlorophyll a (using conversion factors from visual assessments of periphyton cover in up to seven visual categories) distinguished standing crop among sites, and among survey occasions within sites, as effectively as chlorophyll a determined from quantitative samples. Therefore, our hypothesis that inter-operator variability associated with estimating periphyton status using visual assessments would be no greater than that associated with estimates of mean chlorophyll a using quantitative sampling was supported.
We identified significant inter-operator variability in estimates of percentage cover by the No algae, Film and Mat categories. Distinguishing No algae from Film is clearly problematic because there was measurable chlorophyll a in some samples brushed from stones which were visually assessed as having no perceptible cover by algae. However, the conversion procedure highlighted that confusion between the No algae and Film categories is relatively unimportant, particularly if the visual estimates are to be used to derive a single index to approximate the total standing crop of live algae at a site. Compared with other visual categories, both represent small amounts of standing crop.
The largest discrepancies in assigning area to the Mat category occurred in the first survey at two of the sites. This highlights the importance of training and experience in achieving consistency. Practical instruction is likely to be the most effective way to reduce variability in interpretation of categories, and this should encompass a range of river types (Hannaford et al. Citation1997). In New Zealand, training has been achieved through practical courses, and through the use of photographic guides to the different cover categories (e.g. Kilroy et al. Citation2008). Periodic checks for consistency (for example, prior to a change in operators on long-term surveys) are also recommended.
Different assessments could also arise from bias in estimates of percentage cover. For example, one operator might consistently assign a higher or lower percentage cover to the more obvious visual categories, such as green filaments. Operator bias is a potential problem in both visual and measurement-based assessments of stream habitat (Faustini & Kaufman Citation2007; Daniels & McCusker 2011). Point transect methods (e.g. Arscott et al. Citation1998) are generally considered to be more objective (and therefore less subject to bias) than estimating percentage cover by different visual categories or taxa. However, in studies of marine benthic organisms, accuracy and repeatability of visual estimates in quadrats (equivalent to the viewed circles in the present study) compared favourably with random-point methods (Dethier et al. Citation1993) or were no different (Drummond & Connell Citation2005). Furthermore, the visual estimate methods were more efficient (less time-consuming) because they allowed rapid coverage of a larger area (i.e. more replication). There was no evidence of bias in visual assessments between the three observers in the present study, despite each having a different level of experience at the start of the study. Our results therefore confirm the value of previous visual estimates of periphyton cover such as those carried out as part of national water quality monitoring surveys since 1990 (Davies-Colley et al. Citation2011), which have been used to detect 20-year trends in periphyton cover in large rivers (Quinn & Raaphorst Citation2009). The method used in the present study was developed in an effort to provide a more precise methodology that might potentially reduce the long-term costs of monitoring as an alternative to acquiring laboratory-based data on stream periphyton standing crop.
The visual assessment methods currently recommended in New Zealand generally specify 20 views, usually on 2 or 4 transects, to obtain a mean value for periphyton cover (Biggs & Kilroy Citation2000; Kilroy et al. Citation2008). This number was derived from unpublished trials, and is supported by the present results using a relatively stringent test (i.e. for 95% confidence of a difference no greater than 10% from the mean of a large number of views). Our results demonstrated that the optimum number of views is both site- and time-specific. For example, one of the sites (Selwyn) had very heterogeneous periphyton cover over both space and time, and therefore may represent a worst-case scenario in terms of the required intensity of sampling.
In contrast, the number of samples required to obtain a precise estimate of chlorophyll a (within 20% of the mean of all 40 samples in each survey combination) was generally higher. Because chlorophyll a is an open-ended scale, variability among samples can be much higher than for estimates of percentage cover. At the same time, quite large percentage differences in chlorophyll a are not necessarily ecologically significant. For example, the 50% differences between 400 and 600 or 2 and 3 mg m−2 chlorophyll a are probably unimportant. Achieving low percentage differences in the mean is generally only important when we are concerned about whether a chlorophyll a guideline has been breached or not. The numbers of samples required for this were reasonable for practical purposes (five to 25) except when the true mean was close to a threshold.
Current New Zealand periphyton guidelines include both chlorophyll a and percentage cover and both are being applied by river managers (Biggs Citation2000b). Therefore, conversion of multivariate visual cover data to a chlorophyll a equivalent is a practical approach. The approach allows for both overall standing crop estimates and use of visual estimates for specific purposes such as monitoring of percentage cover by potentially toxic cyanobacterial mats (Wood et al. Citation2008).
Despite its wide usage, laboratory-measured chlorophyll a has drawbacks as a metric of algal biomass, including variability as a result of environmental conditions such as light, temperature and community composition (Baulch et al. 2008; Kasprzak et al. Citation2008). Consequently, the conversion factors derived in this study may be biased towards a particular community composition and environment. However, although the three sites surveyed differed in character, the conversion factors derived from the combined data yielded good correspondence with observed chlorophyll a, although biased towards lower values. Further trials on a wider range of rivers would be required to determine whether conversion factors need to be site- or region-specific. The results from an earlier study investigating the relationship between visual estimates and chlorophyll a suggests that there may be consistency across regions (Kilroy et al. Citation2010). Data from monthly periphyton monitoring at 46 sites in the Manawatū-Wanganui region (North Island), revealed strong correspondence between observed and derived chlorophyll a, and conversion factors for the categories Film, Sludge and Mat were very similar to those derived in the present study (4, 65 and 115 mg m−2, compared to 9, 72 and 118 mg m−2, respectively).
An explanation for variability in chlorophyll a estimates among observers might be a tendency to select rocks of a certain size, because rock size and chlorophyll a density tend to be positively related (Uehlinger Citation1991). Our data corroborated this positive relationship, but there was no indication of bias in rock selection between observers. Overall bias in rock selection (a tendency by all three observers to sample rocks larger than the mean size in a separate random rock collection) may explain why derived chlorophyll a was consistently lower than the observed values. Therefore, we suggest that an expectation of lower chlorophyll a derived from visual assessments compared to observed chlorophyll a from samples on individual rocks should be considered when applying the current guidelines (Biggs Citation2000b) for acceptable periphyton biomass in rivers.
It should be noted that biomonitoring using benthic algae often goes beyond simply assessing biomass. Taxonomy-based methods (e.g. using diatoms) are widely used in Europe and North America to identify gradients of degradation (Eloranta & Sioninen Citation2002; Potapova & Charles Citation2007; Kelly et al. Citation2008), and generally distinguish ecological conditions at sites better than biomass alone (Raunio & Soininen Citation2007; Thomas et al. Citation2011). The use of visual categories of periphyton also shows more potential for assessing ecological differences among sites than chlorophyll a alone (Thomas et al. Citation2011). Furthermore, visual assessments may be more useful than biomass estimates because changes to nutrient concentrations and flow regime may not always produce the predicted response in periphyton biomass. For example, there may be a change in community composition rather than in biomass. At the very least, visual assessments could provide data that may assist in understanding such changes, and could be followed up with more detailed taxonomic analyses if necessary.
Conclusion
This study confirmed that it is possible to achieve inter-operator consistency in visual assessments of a range of periphyton categories in rivers using three observers with different levels of experience, but all receiving the same training at the start of the study. The largest discrepancies in visual assessments among observers were for categories of low periphyton biomass, which had an insignificant effect on conversion of the multivariate visual assessment data to a single index based on the chlorophyll a content of each visual category. The visual assessments and the derived chlorophyll a equivalent distinguished periphyton standing crop between sites and occasions as effectively as observed chlorophyll a, but also provided potentially useful information about the type of periphyton growing at each site. We also confirmed the current recommendation (Biggs & Kilroy Citation2000) that 20 views are sufficient to obtain a realistic average periphyton composition, even at a highly heterogeneous site. The surveys were conducted in only three rivers and it is likely that conversion of visual estimates to a single index may require region- or even river-specific conversion factors, which will entail intensive field collections initially. However, our results indicate that inter-operator variability need not be a major concern for ongoing visual assessments, once field observers are trained and familiar with the methodology. We consider that visual assessments of periphyton cover are promising as a defensible method for estimating the biomass of primary production in rivers as an aid to decision-making connected with protection of ecological and other values in rivers.
Acknowledgements
This research was funded by New Zealand Ministry of Science and Innovation, Environmental Flows Programme (C01X0308) and The NIWA Core-Funded programme Sustainable Water Allocation. We thank Kate McArthur, Carol Nicholson and Jon Roygard of Horizons Regional Council for devising and trialling visual assessments in a regional periphyton monitoring programme; and Dave Kelly of Environment Canterbury for help with devising preliminary trials to assess inter-operator variability in visual assessments of periphyton. Chris Cunningham and Helena Campbell (NIWA Christchurch laboratory) provided a fast and efficient sample analysis service.
References
- Arscott , DB , Bowden , WB and Finlay , JC . 1998 . Comparison of epilithic algal and bryophyte metabolism in an arctic tundra stream . Journal of the North American Benthological Society , 17 : 210 – 227 . doi: 10.2307/1467963
- Baulch , HM , Turner , MA , Findlay , DL , Vinebrooke , RD and Donahue , WF . 2009 . Benthic algal biomass – measurement and errors . Canadian Journal of Fisheries and Aquatic Sciences , 66 : 1989 – 2001 . doi: 10.1139/F09-122
- Benenati , EP , Shannon , JP , Blinn , DW , Wilson , KP and Hueftle , SJ . 2000 . Reservoir-river linkages: Lake Powell and the Colorado River, Arizona . Journal of the North American Benthological Society , 19 : 742 – 755 . doi: 10.2307/1468131
- Benstead , JP , Green , AC , Deegan , LA , Peterson , BJ , Slavik , K , Bowden , WB and Hershey , AE . 2007 . Recovery of three arctic stream reaches from experimental nutrient enrichment . Freshwater Ecology , 52 : 1077 – 1089 . doi: 10.1111/j.1365-2427.2007.01723.x
- Bergey , EA and Getty , GM . 2006 . A review of methods for measuring the surface area of stream substrates . Hydrobiologia , 556 : 7 – 16 . doi: 10.1007/s10750-005-1042-3
- Biggs , B . 1996 . “ Patterns in benthic algae of streams ” . In Algal ecology: freshwater benthic ecosystems , Edited by: Stevenson , R , Bothwell , ML and Lowe , RL . 31 – 56 . San Diego , CA : Academic Press .
- Biggs , BJF . 2000a . Eutrophication of streams and rivers: dissolved nutrient – chlorophyll relationships for benthic algae . Journal of the North American Benthological Society , 19 : 17 – 31 . doi: 10.2307/1468279
- Biggs , BJF . 2000b . New Zealand periphyton guideline: detecting, monitoring and managing enrichment of streams , Wellington , , New Zealand : Ministry for the Environment .
- Biggs , BJF and Close , ME . 1989 . Periphyton biomass dynamics in gravel bed rivers: the relative effects of flows and nutrients . Freshwater Biology , 22 : 209 – 231 . doi: 10.1111/j.1365-2427.1989.tb01096.x
- Biggs , BJF and Kilroy , C . 2000 . Stream periphyton monitoring manual. Published by NIWA for Ministry for the Environment , 226 Christchurch , , New Zealand : NIWA .
- Black , RW , Moran , PW and Frankforter , JD . 2011 . Response of algal metrics to nutrients and physical factors and identification of nutrient thresholds in agricultural streams . Environmental Monitoring and Assessment , 175 : 397 – 417 . doi: 10.1007/s10661-010-1539-8
- Bowden , WB , Finlay , JC and Maloney , PE . 1994 . Long-term effects of PO4 fertilization on the distribution of bryophytes in an arctic river . Freshwater Biology , 32 : 445 – 454 . doi: 10.1111/j.1365-2427.1994.tb01138.x
- Bourassa , N and Cattaneo , A . 2000 . Responses of a lake outlet community to light and nutrient manipulation: effects on periphyton and invertebrate biomass and composition . Freshwater Biology , 44 : 629 – 639 . doi: 10.1046/j.1365-2427.2000.00610.x
- Champion , PD and Tanner , CC . 2000 . Seasonality of macrophytes and interaction with flow in a New Zealand lowland stream . Hydrobiologia , 441 : 1 – 12 . doi: 10.1023/A:1017517303221
- Chétalat , J , Pick , F , Morin , A and Hamilton , P . 1999 . Periphyton biomass and community composition in rivers of different nutrient status . Canadian Journal of Aquatic Sciences , 56 : 560 – 569 . doi: 10.1139/f98-197
- Daniels , MD and McCusker , MH . 2010 . Operator bias characterizing stream substrates using Wolman pebble counts with a standard measurement template . Geomorphology , 115 : 194 – 198 . doi: 10.1016/j.geomorph.2009.09.038
- Davies-Colley , RJ , Smith , DG , Ward , RC , Bryers , GG , McBride , GB , Quinn , JM and Scarsbrook , MR . 2011 . Twenty years of New Zealand's river water quality monitoring network: benefits of careful design and consistent operation . Journal of the American Water Resources Association , 47 : 750 – 771 . doi: 10.1111/j.1752-1688.2011.00554.x
- Dethier , MN , Graham , ES , Cohen , S and Tear , LM . 1993 . Visual versus random-point percent cover estimations: ‘objective’ is not always better . Marine Ecology Progress Series , 96 : 93 – 100 . doi: 10.3354/meps096093
- Dodds , WK , Smith , VH and Lohman , K . 2002 . Nitrogen and phosphorus relationships to benthic algal biomass in temperate streams . Canadian Journal of Fisheries and Aquatic Sciences , 59 : 865 – 874 . doi: 10.1139/f02-063
- Drummond , SP and Connell , SD . 2005 . Quantifying percentage cover of subtidal organisms on rocky coasts: a comparison of the costs and benefits of standard methods . Marine and Freshwater Research , 56 : 865 – 876 . doi: 10.1071/MF04270
- Eloranta , P and Soininen , J . 2002 . Ecological status of some Finnish rivers evaluated using benthic diatom communities . Journal of Applied Phycology , 14 : 1 – 7 . doi: 10.1023/A:1015275723489
- Faustini , JM and Kaufmann , PR . 2007 . Adequacy of visually classified particle count statistics from regional stream habitat surveys . Journal of the American Water Resources Association , 43 : 1293 – 1315 . doi: 10.1111/j.1752-1688.2007.00114.x
- Flinders , CA and Hart , DD . 2009 . Effects of pulsed flows on nuisance periphyton growths in rivers: a mesocosm study . River Research and Applications , 25 : 1320 – 1330 . doi: 10.1002/rra.1244
- Hambrook Berkman JA , Canova , MG 2007 . Algal biomass indicators (ver. 1.0): US Geological Survey techniques of water-resources investigations , book 9, chap. A7, section 7.4, August, available online only from http://pubs.water.usgs.gov/twri9A/ .
- Hannaford , MJ , Barbour , MT and Resh , VH . 1997 . Training reduces observer variability in visual-based assessments of stream habitat . Journal of the North American Benthological Society , 16 : 853 – 860 . doi: 10.2307/1468176
- Heath , MW , Wood , SA and Ryan , KG . 2011 . Spatial and temporal variability in Phormidium mats and associated anatoxin-a and homoanatoxin-a in two New Zealand rivers . Aquatic Microbial Ecology , 64 : 69 – 79 . doi: 10.3354/ame01516
- Kasprzak , P , Padisak , J , Koschel , R , Krienitz , L and Gervais , F . 2008 . Chlorophyll a concentration across a trophic gradient of lakes: an estimator of phytoplankton biomass? . Limnologica , 38 : 327 – 338 . doi: 10.1016/j.limno.2008.07.002
- Kelly , M , Juggins , S , Guthrie , R , Pritchard , S , Jamieson , J , Rippey , B , Hirst , H and Yallop , M . 2008 . Assessment of ecological status in UK rivers using diatoms . Freshwater Biology , 53 : 403 – 422 .
- Kilroy C , Biggs BJF , Death R 2008 . A periphyton monitoring plan for the Manawatū-Wanganui region . NIWA Client Report CHC2008-03. http://www.envirolink.govt.nz/PageFiles/459/446-HZLC50.pdf . Christchurch , New Zealand , NIWA . 43
- Kilroy C , Nicholson C , McArthur K , Chakraborty M , Roygard J 2010 . Review of Horizons Regional Council periphyton monitoring programme 2009 . NIWA Client Report CHC2010-59 . Christchurch , New Zealand , NIWA . 96
- Lessard J , Arscott DB , Booker DJ , Larned S , Snelder TJ , Suren AM 2013 . Dam design must provide for both high and low flows: a case study from the Ōpuha Dam , South Island, New Zealand . Journal of Environmental Management 5 459 – 473 .
- Ministry for the Environment 2011 . National policy statement for freshwater management 2011: implementation guide . Wellington , , New Zealand , Ministry for the Environment . 50
- Necchi , O , Branco , LHZ and Branco , CCZ . 1995 . Comparison of three techniques for estimating periphyton abundance in bedrock streams . Archiv für Hydrobiologie , 134 : 393 – 402 .
- New Zealand Conservation Authority 2011 . Protecting New Zealand's rivers . Wellington , New Zealand Conservation Authority . 68
- Potapova , M and Charles , DF . 2007 . Diatom metrics for monitoring eutrophication of rivers of the United States . Ecological Indicators , 7 : 48 – 70 . doi: 10.1016/j.ecolind.2005.10.001
- Quinn JM , Raaphorst E 2009 . Trends in nuisance periphyton cover at New Zealand national river water quality network sites 1990–2006 . NIWA Client Report 2008-194. http://www.mfe.govt.nz/publications/water/periphyton-nz-national-20river-quality-network-1990-2006/index.html . Wellington , , New Zealand , Ministry for Environment . 22
- Raunio , J and Soininen , J . 2007 . A practical and sensitive approach to large river periphyton monitoring: comparative performance of methods and taxonomic levels . Boreal Environment Research , 12 : 55 – 63 .
- Rodriguez , RR , Jiménez , JC and Delgado , CM . 2007 . Microhabitat and morphometric variation in two species of Prasiola (Prasiolales, chlorophyta) from streams in central Mexico . Aquatic Ecology , 41 : 161 – 168 . doi: 10.1007/s10452-006-9068-9
- Stevenson J , Bahls LL 1999 . Periphyton protocols . In : Barbour MT , Gerritsen J , Snyder BD , Stribling JB . Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish, , 2nd edition EPA 841-B-99-002 . Washington , DC , Office of Water, US Environmental Protection Agency . Chapter 6, pp. 6.1–6.23.
- Strong , CW . 1966 . An improved method for obtaining density from line-transect data . Ecology , 47 : 311 – 313 . doi: 10.2307/1933781
- Suren , A , Biggs , B , Duncan , M , Bergey , E and Lambert , P . 2003 . Benthic community dynamics during summer low-flows in two rivers of contrasting enrichment 2. Invertebrates . New Zealand Journal of Marine and Freshwater Research , 37 : 71 – 83 . doi: 10.1080/00288330.2003.9517147
- Thomas , KE , Kluke , A , Hall , RI , Paterson , AM and Winter , JG . 2011 . Assessment of benthic algal biomonitoring protocols to evaluate effects of shoreline development on the nearshore zone of Precambrian Shield lakes in Ontario . Lake and Reservoir Management , 27 : 398 – 413 . doi: 10.1080/07438141.2011.633307
- Uehlinger , U . 1991 . Spatial and temporal variability if the periphyton biomass in a prealpine river (Necker, Switzerland) . Archiv Fur Hydrobiologie , 123 : 219 – 237 .
- Wikum , DA and Shanholtzer , GF . 1978 . Application of the Braun-Blanquet cover-abundance scale for vegetation analysis in land development studies . Environmental Management , 2 : 323 – 329 . doi: 10.1007/BF01866672
- Wolman , MG . 1954 . A method of sampling coarse river bed material. Transactions . American Geophysical Union , 35 : 951 – 956 .
- Wood , KA , Stillman , RA , Clarke , RT , Daunt , F and O'Hare , MT . 2012 . Measuring submerged macrophyte standing crop in shallow rivers: a test of methodology . Aquatic Botany , 102 : 28 – 33 . doi: 10.1016/j.aquabot.2012.04.006
- Wood SA , Hamilton DP , Paul WJ , Safi KA , Williamson WM 2008 . New Zealand guidelines for cyanobacteria in recreational fresh waters: interim guidelines . Publication 981 . Wellington , , New Zealand , Ministry for the Environment . 82
- Wu , N , Tang , T , Zhou , S , Jia , X , Li , D , Liu , R and Cai , Q . 2009 . Changes in benthic algal communities following construction of a run-of-river dam . Journal of the North American Benthological Society , 28 : 69 – 79 . doi: 10.1899/08-047.1