Abstract
Floods are an integral part of flowing river systems, yet our understanding of freshwater fish behaviour during floods is limited. In 2008, 67 redfin bullies (Gobiomorphus huttoni), 22 shortjaw kōkopu (Galaxias postvectis) and 29 kōaro (Galaxias brevipinnis) were tagged with passive integrated transponder (PIT) tags and monitored in a 100-m study reach before, during or after three floods. Detection patterns of tagged fish indicated that they may be displaced or may move to areas with large substrates during floods. Two individuals returned to the same locations during multiple floods, suggesting that previous experience may influence refuge selection for some individuals. While small changes in community composition occurred that could be attributed to concurrent habitat changes, overall a remarkable level of persistence of individual fish was observed in the tagged community.
Introduction
The ecological integrity of flowing water environments depends on their variable nature (Poff et al. Citation1997) and the harm that alteration of this hydrologic variability (through dams, diversions, drainage, channelisation and groundwater extraction) can cause to riverine ecosystems is well documented (Poff et al. Citation1997 and references therein). Despite this, human desire to ensure that freshwater supply and movement is as predictable and as controllable as possible has dominated our relationship with the freshwater ecosystems we live amongst, and anthropogenic alteration of natural flow regimes through structures such as dams has had negative effects on aquatic biota (Petts Citation1984; Hunt Citation1988; Hunter Citation1992; Barinaga Citation1996; Power et al. Citation1996; Graf Citation1999; Kingsford Citation2000; Holland Citation2001). Some organisms have developed or evolved physical, physiological and behavioural traits that allow them to survive and even depend on disturbances (Lytle & Poff Citation2004). For example, numerous freshwater fishes spawn following floods to safeguard offspring from drought conditions (John Citation1963; Nesler et al. Citation1988; Fausch & Bestgen Citation1997; King et al. Citation2003). Many freshwater fish possess morphological, physiological and life history traits that enable them to adapt to or take advantage of periods of high discharge (Lytle & Poff Citation2004 and references therein). However, descriptions of behavioural adaptations are rare in the literature, probably due to the logistical difficulty associated with observing fish during flood conditions (Matthews Citation1998). Nevertheless, understanding the relative importance of high flow events for particular riverine ecosystems and for the biota they support is crucial for effective water resource management. In New Zealand, hydroelectricity generation is common and many major rivers have a dam at some point along their length. Dams alter physical habitat and flow regimes, including the conversion of lotic habitat to lentic, cause erratic flow patterns downstream (Bunn & Arthington Citation2002) and reduce frequency and magnitude of floods (Lytle & Poff Citation2004). The highly endemic native freshwater fish fauna of New Zealand is in a general state of ongoing decline (McDowall Citation1990; Allibone et al. Citation2010) and greater understanding of the relationships between freshwater fish and flow regimes is essential if the fauna is to be preserved.
Most research on freshwater fish responses to flooding has been conducted in a before and after framework. Such studies report that the immediate effects of high discharge events on freshwater fish show that such effects can be species specific (Matthews Citation1986; Jowett & Richardson Citation1989; Dudley & Matter Citation1999), size specific (Allen Citation1951; Elwood & Waters Citation1969) and habitat specific (Jowett & Richardson Citation1989; Godlewska et al. Citation2003). Studies that conduct follow up sampling two or more weeks after a flood (Matthews Citation1986 and references therein; Chapman & Warburton Citation2006) show that most fish community parameters return to a pre-flood state providing significant habitat alteration does not occur. There are some studies available that report on fish behaviour during flood events: Meffe (Citation1984) used a stream simulator to show that Sonoran topminnows (Poeciliopsis occidentalis (Baird & Girard, 1853)) oriented themselves into the current and sought out areas where the current was slowest. In addition, recent advances in radio telemetry have allowed fish behaviour during floods to be documented in the field, e.g., studies have shown that northern hog suckers Hypentelium nigricans (Lesuer, 1817) in the USA (Matheney & Rabeni Citation1995), giant kōkopu Galaxias argenteus (Gmelin, 1789) in New Zealand (David & Closs Citation2002) and river blackfish Gadopsis marmoratus (Richardson 1848) in Australia (Koster & Crook Citation2008) move towards the water's edge during episodes of high flow.
The importance of the ecological elements of natural flow regimes such as magnitude, frequency, seasonal timing, predictability, duration and rate of change (Lytle & Poff Citation2004) to the New Zealand native freshwater fish fauna is likely high due to the unstable nature of many New Zealand streams (Winterbourne et al. Citation1981). Some species of Galaxias use high discharge events to spawn in temporarily inundated riparian margins (McDowall Citation1990; Allibone & Caskey Citation2000; Charteris et al. Citation2003), while accounts also exist of fishes using newly inundated terrain to exploit new food resources that become available during floods (Ross & Baker Citation1983; Turner et al. Citation1994). This paper aims to contribute to existing knowledge regarding the response of freshwater fishes to flooding, particularly behaviour during flood events. We present findings generated from opportunistic data collection associated with a long term instream passive integrated transponder (PIT) tag monitoring study (McEwan & Joy Citation2011; McEwan & Joy Citationin press), during which three significant floods occurred.
Methods
Study site
The Mangaore Stream, in the lower North Island of New Zealand is a third order tributary of the Manawatu River with a steep gradient catchment located in the foothills of the Tararua mountain ranges (40°35'24.69”S; 175° 26'11.61”E). The average base flow ranges from approximately 0.5 to 1.0 cumecs and the wetted environment is mostly less than 5 m wide and less than 1 m deep and contains predominantly cobble and boulder substrates. The Mangaore Stream frequently experiences brief yet intense periods of high flow. This stream also possesses a diverse and abundant native fish community (McEwan & Joy Citation2011), providing a suitable situation to examine behavioural responses in a varied hydrological environment. A small hydroelectricity station is located on the stream just east of the Shannon township (below the study reach). The dam itself is not a physical barrier on the Mangaore Stream (water is collected and stored from the neighbouring Mangahao catchment), although during peak generation discharges, high flows may present a velocity barrier to upstream migrating fish. The methodology described below formed part of a larger project involving repeated surveying of PIT-tagged fishes within an open, 100-m reach of the stream from January to July 2008 (McEwan & Joy Citation2011). Three floods occurred during this period, which facilitated the collection of data regarding the locations of tagged fish associated with flood events.
Habitat mapping
Stream wetted width and physical features (e.g., boulders and logs) over the 100-m reach were mapped by point triangulation using a laser distance meter at a 0.25-m2 grid scale. These measurements were then transposed onto paper using a ruler and compass at a scale of 1 m = 1 cm. Stream bed, wetted edge and any physical habitat features (e.g., logs) were sketched and transposed to the map using a 1-cm2 grid onto which the spatial positions of PIT-tagged fish were recorded. Substrate size was assessed by first recording the proportion of the benthos of each grid square that was covered in each of eight substrate types: boulders, large cobbles, small cobbles, large gravel, small gravel, sand, silt and bedrock (using particle size classes from Wentworth Citation1922). These substrate composition variables were then condensed for each grid square into a single variable by summing the products of the minimum axis length of each particle size class (with sand, silt and bedrock being assigned lengths of 0.5, 0.1 and 0 mm, respectively) multiplied by the proportion of the benthos covered by that particle size class (see for example calculation). The resulting substrate size scores ranged from 0 (smallest) to 256 (largest). Habitat mapping was completed for the wetted area of the reach at base flow and was carried out in February (prior to the April flood) and again directly following the April flood. The study reach remained open so PIT-tagged fish (and fish without PIT tags, for which we did not collect information) could move freely in and out of the reach.
Table 1 An example showing how a substrate size score was calculated for each grid square.
PIT-tagging and monitoring
Three native diadromous fish species were PIT-tagged and monitored; 67 redfin bully (Gobiomorphus huttoni (Ogilby, 1894)), 22 shortjaw kōkopu (Galaxias postvectis Clarke, 1899) and 29 kōaro (Galaxias brevipinnis Günther, 1866; ; common names used hereafter). All three species are classified as ‘At Risk: Declining’ by the New Zealand Department of Conservation (Allibone et al. Citation2010). Fish were collected and tagged in the study reach during two periods: 01–03 January 2008 and 21–24 March 2008. They were collected using a combination of multiple pass electro-fishing and repeated spotlighting (locating fish at night with a spotlight, then dip-netting) and tagged according to protocols detailed in McEwan & Joy (Citation2011). Once implanted, fish were allowed to recover in in-stream cages for approximately half an hour before being released back into the approximate region of the study reach where they were collected.
Table 2 Summary information for 120 native freshwater fish that were implanted with 12.5-mm passive integrated transponder (PIT) tags in the Mangaore Stream, New Zealand in 2008.
The PIT tag antenna (BiomarkTM racquet antenna with waterproof coating) was affixed to a wooden pole and the PIT tag reader (FS2001FR-ISO) was attached to a harness so the screen could be viewed by a mobile researcher at any time. Monitoring of the study reach was conducted by a single researcher systematically moving upstream and slowly scanning the entire bed. A copy of the stream map was carried on each sampling occasion and the location of each tag entered in the corresponding grid square on the map. During sampling, care was taken to cause as little physical disturbance as possible and the presence of large boulders meant that the researcher was usually able to avoid stepping in wetted areas. Each sample took approximately 2 h. Daytime surveys were carried out any time between 09:00 and 15:00 h and night-time surveys were carried out between 1900 h and midnight, commencing at least 1 h after sunset. The section was sampled on 25 occasions during the day and on 15 occasions at night (assisted by spotlight) over March to July 2008. During this time three floods (using the definition of ‘flood’ from David & Closs [Citation2002] as greater than five times base flow discharge) occurred, allowing us to not only survey for PIT-tagged fish during flood conditions but also to examine community parameters in a before/after context. Base flow and flood flow figures were obtained from the peaking station located approximately 5 km downstream. While these figures cannot be applied directly to the study site, they serve to provide a comparison between flood flows and base flows in the Mangaore Stream in general and thus a measure of flood magnitude. A depth-gauging station was used as a proxy measure of flow magnitude at the site. During flood conditions, comprehensive surveying of some sections of the stream was limited but reasonable coverage of all areas was still possible from large exposed boulders and outcrops scattered throughout the stream.
Floods
Before and after data related to individual fish presence/absence was obtained for the January flood and the April flood, and in-flood sampling was conducted for the April flood and the July flood. The first flood event occurred four days after the first group of fish were tagged in January and was of exceptional magnitude. The flood flow measured at the power station peaked at 99.87 cumecs, which was almost 200 times the average base flow of 0.5 cumecs. This flood caused significant habitat change, including the main flow within the study section being shifted from the true left to the true right of the riverbed. No surveying was able to be conducted during this flood due to its magnitude, however we were able to collect before/after data based on fish PIT-tagged before the flood. The April flood, during which the stream was surveyed twice, once during the day and once at night, reached a peak flow of 41.36 cumecs (c. 83 times average base flow of 0.5 cumecs). Before/after data associated with this flood was also collected. The July flood peaked at 25.39 cumecs (c. 25 times average base flows of c. 1.0 cumec) and produced the opportunity to survey twice on two consecutive days. As the survey regime ceased immediately following this flood, before/after comparisons were not able to be made. The ‘before’ group for the January flood was composed of all fish PIT-tagged prior to the flood and the ‘after’ group composed of all fish that were detected in the reach following this flood and prior to the April flood. The ‘before’ group for the April flood was composed of the ‘after’ group from the January flood as well as the second group of fish tagged in March. Finally, the ‘after’ group for the April flood consisted of all fish detected in the reach following this flood. The inclusion of fish that were detected a number of weeks prior to a flood in the ‘before’ group was justified by the high degree of site fidelity that was observed in tagged fish (McEwan Citation2009; McEwan unpubl. data).
Data analysis
Tags located during flood conditions were recorded and examined for position relative to the base flow stream bed (inside or outside the bed or close to the original wetted edge). Substrate size index scores in habitats where tags were found inside the stream bed were compared with scores from a randomly selected (using the RAND function in Microsoft Excel) subset of all habitats available using Wilcoxon–Mann–Whitney tests (R development Core Team Citation2011). This was also done for data from non-flood samples for the purposes of comparison. For fish that were detected more than once, substrate data were averaged in order to produce a single value per individual (to avoid within-group data dependence in analyses). To investigate movement of individuals and their frequency of detection at local scales, we developed a simple index of ‘familiarity’ whereby each individual tag detection would be assigned a score between zero and ten based on when that tag was previously detected. For example, if a tag was found during the previous 10 samples then that detection would score 10 (very familiar); if it hadn't been detected during the previous 10 samples then it would score 0 (very unfamiliar); if it had been detected during five of the previous 10 samples then it would score 5 (medium familiarity). All scores were then summed and divided by the total number of tags detected to produce an average score of ‘community familiarity’ for each survey. These scores were then compared with stage depth (using a Wilcoxon–Mann–Whitney test) to investigate whether PIT tag familiarity was related to stage height.
Results
Before and after floods
In total, 56% of all PIT-tagged fish present in the study section before the January flood were detected in the study reach after this event (), despite the significant physical changes that occurred. This figure was higher for the smaller April flood, with 63% of all fish that were detected in the study reach before the April flood also being detected after the April flood. The percentages of redetected shortjaw kōkopu were approximately the same for both flood events (57% and 61% respectively), but redfin bullies and kōaro showed differences: only 52% of tagged redfin bullies were detected in the study reach following the larger January flood, compared with 71% detected following the April flood. Sixty-on per cent of tagged kōaro were detected in the study reach following the January flood, compared with only 45% detected following the April flood.
Table 3 Numbers of passive integrated transponder (PIT)-tagged fish that were detected in a 100-m reach of the Mangaore Stream, Manawatu, New Zealand before and after two floods in 2008.
During floods
A total of 31 PIT-tagged fish (22 redfin bullies, six kōaro and three shortjaw kōkopu) were detected within the study section over four surveys conducted while the stream was in flood. Three redfin bullies and one kōaro were detected during two or more flood surveys. One redfin bully (RF14) was found in the same location (within 0.25 m2) during both the April and July floods, despite being found (nine redetections) occupying a different area during non-flood conditions. Another redfin bully (RF20) was found once during the April flood and twice during the July flood, in all cases within the same 1-m2 area. This area was distinct from the area (15 redetections ranging from around 5–15 m away) in which this fish was found during base flow conditions. Seventeen fish were detected in areas within the base flow stream bed that were able to be matched with substrate size data (multiple values for two fish were averaged to facilitate statistical comparison) and these habitats had significantly higher substrate size index scores (151±16; mean±1 SEM) than scores from a randomly selected set of all available habitats (97±12; W 17, 17=5.75, P=0.024). During non-flood samples, the habitats in which fish were detected also had significantly higher scores (152±4) than a group of randomly selected habitats (106±4; W 351,351=37612, P<0.001).
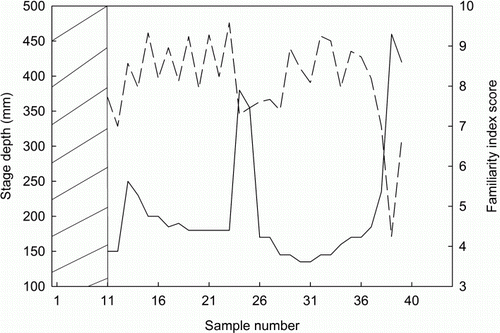
Both the April and July floods were accompanied by abrupt decreases in community familiarity (). The mean community familiarity score during flood surveys (n=4) was 6.4±0.7, compared with a higher mean score of 8.3±0.1 for non-flood surveys (n=27). When small increases in depth were taken into account however, no significant difference in familiarity was observed overall: for surveys when stage depth was at or below median (180 mm; n=20), mean familiarity was 8.4±0.2 and mean familiarity for surveys when stage depth was above median (n=11) was 7.4±0.4 (W=159; P=0.096).
Figure 1 Plots showing correlation between stage depth at each sampling occasion (solid line) and community familiarity score (dashed line). Scores were calculated for each sampling occasion based on the number of times each individual fish was found during the previous 10 sampling occasions. 0 = highly unfamiliar; 10 = highly familiar. Shaded area represents the first 10 sampling occasions, for which familiarity was not calculated. Samples 24 and 25 occurred during the April flood and samples 39 and 40 during the July flood.
Discussion
Changes in species' distributions following floods
David & Closs (Citation2002) found that individuals were less likely to return to pre-flood territories and Matthews (Citation1986) and Yoon et al. (Citation2011) found that communities were less likely to return to a pre-flood state if local habitat parameters were altered as a result of high flows. In the present study, changes in relative numbers of three fish species were observed following floods. These changes may have been due to visually assessed habitat changes that occurred as a result of flooding—the changes produced by the January flood may have favoured kōaro and the changes produced by the April flood may have favoured redfin bullies. Prior to the January flood, the study reach contained a high proportion of shallow, slow flowing pools containing large substrates and high amounts of interstitial refuge space—highly suitable conditions for redfin bullies and less suitable for kōaro (McDowall Citation1990). The January flood caused wide scale scouring, resulting in a greater proportion of run and riffle zones than previously, thereby reducing amounts of suitable habitat for redfin bullies and creating more habitat suited to kōaro.
The April flood resulted in the opposite (albeit smaller–scale) habitat changes. Material was washed downstream and deposited in the study section, which resulted in a general reduction in high velocity riffle zones. These changes rendered the section less suitable than previously for riffle dwelling kōaro. A number of the kōaro that were absent from the reach following the April flood were detected close to the reach (during separate monitoring not reported on here) in a small, riffle–filled side channel that was formed by the April flood. Interspecific competition may also have played a role in the changes that occurred although it is difficult to comment on this further as nothing is currently known regarding biotic interactions of these species. Redfin bullies were the smallest-bodied species included in the study, and appeared to be the most affected by flood magnitude. Of all the tagged redfin bullies that were found in the study section before the April flood, 30% were not found after the flood, whereas this figure was 52% following the more severe January flood. Studies have found that smaller trout are more likely to be affected by floods than larger trout (Allen Citation1951; Elwood & Waters Citation1969; Heggenes Citation1988; Jowett & Richardson Citation1989). Although it is possible that the fish that were not detected were actually still in the study reach, it is unlikely, given the repeated nature of the sampling regime that was underway. This is particularly true for the January flood, as the reach was surveyed on 40 separate occasions afterwards. Fifteen of these occurred after the April flood. For this reason, we are confident that the non-detections of PIT-tagged fish in this context do represent absence from the study reach.
Behaviour during floods
PIT-tagged fish were found in areas with large substrate particles during both flood surveys and non-flood surveys. Large floods have the potential to exert mortality on fish through substrate movement (Lytle & Poff Citation2004) and Heggenes (Citation1988) found that coarse substrates provided shelter for brown trout (Salmo trutta Linnaeus, 1758) and prevented downstream displacement during high flows in a small stream. During this study, two separate individuals were observed returning to the same area during multiple floods, which may indicate that some individuals habitually use specific localities during extreme hydrological conditions. The observed reductions in the familiarity of tags that occurred during floods could represent downstream displacement or more purposeful movements of the tagged fish. Some freshwater fish species are known to use high flows for reproduction (McDowall Citation1990; Allibone & Caskey Citation2000; Charteris et al. Citation2003) and feeding (Ross & Baker Citation1983; Turner et al. Citation1994), thus reductions in familiarity could represent greater mobility of the study population during elevated flows. This may cause temporary species’ redistribution—familiarity in the study reach returned to pre-flood levels following the April flood, indicating that species' redistribution that occurred as a result of this flood was only temporary. Matthews (Citation1986) found that fish communities in Piney Creek, Missouri, USA, were altered immediately following floods and these alterations were only temporary.
Overall, this study has shown that these three New Zealand native fish species appear to express adaptive behaviours, which allow them to avoid harm during episodes of high flow, although these behaviours may be less effective in especially severe floods for redfin bullies. While some habitat change did occur following floods, overall a remarkable level of persistence of individual fish was observed in the tagged population. Despite the severity of the January flood event and the large-scale visual changes to the study reach geomorphology, over half of the individuals that were found before this event were also found there afterwards. For the less severe April flood, close to two-thirds of the pre-flood population were also found in the reach following the flood. Previous studies have shown that fish communities tend to return to pre-flood conditions after time—the same numbers of the same species (Matthews Citation1986 and references therein; Chapman & Warburton Citation2006)—whereas the present study has shown that it is mostly the same individuals making up that community.
Implications for flow management in New Zealand
New Zealand has a highly variable maritime climate, steep, mountainous terrain and rivers are typically short and unstable (Winterbourn et al. Citation1981). These factors suggest that disturbance is common and New Zealand indigenous fish are adapted to living in disturbed environments. The consequences of loss or modification of disturbance events such as floods can include native habitat loss, recruitment failure, loss of native biodiversity and the proliferation of invasive species—populations of which would formerly have been controlled by flood events (Bunn & Arthington Citation2002). It is not known whether New Zealand species that spawn during floods actually require floods in order to spawn; however, it is likely, given that eggs are laid in riparian vegetation (McDowall Citation1990; Allibone & Caskey Citation2000; Charteris et al. Citation2003). Hydro-generated electricity is common in New Zealand and discussion regarding the effects of dams on aquatic biota is a regular feature of resource consent hearings associated with dam operations. Pulsed flow releases from dams such as peaking, flushing and recreational flows can cause stranding, downstream displacement of fishes and reduced spawning and rearing success (Young et al. Citation2011 and references therein), indicating that the character of such anthropogenic disturbance events may not provide appropriate cues for fish to use in order to react appropriately. In some cases, pulsed flow releases can be manipulated in order to minimise negative impacts downstream, for example, salmonids in Californian rivers are less susceptible to stranding when down-ramping rates (rates of flow reduction following a pulsed release) are reduced to produce a slow tail-off back to base flows instead of an abrupt cessation in flow (Young et al. Citation2011). The findings of this study indicate that these species of native fish have adopted behaviours that allowed communities and individuals to persist during and after flood events in a small stream. Better understanding of the behaviour of native fish during and after episodes of high flow in larger rivers is needed in order to recommend pulsed-flow management that avoids, minimises or reverses the impacts of altered flow regimes from human-made structures such as dams in New Zealand.
Acknowledgements
We gratefully acknowledge Patrick and Marlene Anderson for allowing access to the Mangaore Stream, to Alton Perrie for assistance with fieldwork and Dr Bruno David for helpful comments on the manuscript. The Department of Conservation, the Whanganui River Enhancement Trust, the family of the late Julie Alley and Greater Wellington Regional Council provided assistance with funding and equipment. Thanks to Peter Galpin and the Mangahao Power Station for stream flow data. This research was sanctioned by the Massey University Animal Ethics Committee (protocol No. 07/30 and 07/106).
References
- Allen KR 1951 . The Horokiwi Stream: a study of a trout population . New Zealand Marine Department . Fish Bulletin 10 . Wellington , , New Zealand . 231
- Allibone , RM and Caskey , D . 2000 . Timing and habitat of k(aro (Galaxias brevipinnis) spawning in streams draining Mt Taranaki, New Zealand . New Zealand Journal of Marine and Freshwater Research , 34 : 593 – 595 . doi: 10.1080/00288330.2000.9516960
- Allibone , R , David , B , Hitchmough , R , Jellyman , D , Ling , N Ravenscroft , P, . 2010 . Conservation status of New Zealand freshwater fish, 2009 . New Zealand Journal of Marine and Freshwater Research , 44 : 271 – 287 . doi: 10.1080/00288330.2010.514346
- Barinaga , M . 1996 . A recipe for river recovery? . Science , 273 : 1648 – 1650 . doi: 10.1126/science.273.5282.1648
- Bunn , SE and Arthington , AH . 2002 . Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity . Environmental Management , 30 : 492 – 507 . doi: 10.1007/s00267-002-2737-0
- Chapman , P and Warburton , K . 2006 . Postflood movements and population connectivity in gambusia (Gambusia holbrooki) . Ecology of Freshwater Fish , 15 : 357 – 365 . doi: 10.1111/j.1600-0633.2006.00145.x
- Charteris , SC , Allibone , RM and Death , RG . 2003 . Spawning site selection, egg development, and larval drift of Galaxias postvectis and G. fasciatus in a New Zealand stream . New Zealand Journal of Marine and Freshwater Research , 37 : 493 – 505 . doi: 10.1080/00288330.2003.9517184
- David , BO and Closs , GP . 2002 . Behaviour of a stream-dwelling fish before, during, and after high-discharge events . Transactions of the American Fisheries Society , 131 : 762 – 771 . doi: 10.1577/1548-8659(2002)131%3C0762:BOASDF%3E2.0.CO;2
- Dudley , RK and Matter , WJ . 1999 . Effects of a record flood on fishes in Sabino Creek, Arizona . Southwestern Naturalist , 44 : 218 – 221 .
- Elwood , JW and Waters , TF . 1969 . Effects of floods on food consumption and production rates in a stream brook trout population . Transactions of the American Fisheries Society , 98 : 253 – 262 . doi: 10.1577/1548-8659(1969)98[253:EOFOFC]2.0.CO;2
- Fausch KD , Bestgen KR 1997 . Ecology of fishes indigenous to the Central and Southwestern Great Plains . In : FL Knopf , FB Samson . Ecology and conservation of Great Plains vertebrates . New York , , Springer-Verlag . 131 – 166 .
- Godlewska , M , Mazurkiewicz-Boron , G , Pociecha , A , Wilk-Wozniak , E and Jelonek , M . 2003 . Effects of flood on the functioning of the Dobczyce reservoir ecosystem . Hydrobiologia , 504 : 305 – 313 . doi: 10.1023/B:HYDR.0000008530.31142.81
- Graf , WL . 1999 . Dam nation: a geographic census of American dams and their large-scale hydrologic impacts . Water Resources Research , 35 : 1305 – 1311 . doi: 10.1029/1999WR900016
- Heggenes , L . 1988 . Effects of short-term flow fluctuations on displacement of, and habitat use by, brown trout in a small stream . Transactions of the American Fisheries Society , 117 : 336 – 344 . doi: 10.1577/1548-8659(1988)117%3C0336:EOSFFO%3E2.3.CO;2
- Holland , E . 2001 . The state of California Rivers , 470 California : The Trust for Public Land .
- Hunt , CE . 1988 . Down by the river: the impact of federal water projects and policies on biological diversity , 256 Washington : Island Press .
- Hunter MA 1992 . Hydropower flow fluctuations and salmonids: a review of the biological effects, mechanical causes, and options for mitigation . Technical Report 119 . Washington Department of Fisheries , Olympia , , USA .
- John , KR . 1963 . The effect of torrential rain on the reproductive cycle of Rhinichthys osculus in the Chiricahua Mountains, Arizona . Copeia , 1 : 286 – 291 . doi: 10.2307/1441345
- Jowett , IG and Richardson , J . 1989 . Effects of a severe flood on instream habitat and trout populations in 7 New Zealand rivers . New Zealand Journal of Marine and Freshwater Research , 23 : 11 – 17 . doi: 10.1080/00288330.1989.9516335
- King , AJ , Humphries , P and Lake , PS . 2003 . Fish recruitment on flood plains: the roles of patterns of flooding and life history characteristics . Canadian Journal of Fisheries and Aquatic Sciences , 60 : 773 – 786 . doi: 10.1139/f03-057
- Kingsford , RT . 2000 . Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia . Austral Ecology , 25 : 109 – 127 . doi: 10.1046/j.1442-9993.2000.01036.x
- Koster , WM and Crook , DA . 2008 . Diurnal and nocturnal movements of river blackfish (Gadopsis marmoratus) in a south-eastern Australian upland stream . Ecology of Freshwater Fish , 17 : 146 – 154 . doi: 10.1111/j.1600-0633.2007.00269.x
- Lytle , DA and Poff , NL . 2004 . Adaptation to natural flow regimes . Trends in Ecology and Evolution , 19 : 94 – 100 . doi: 10.1016/j.tree.2003.10.002
- McDowall , RM . 1990 . New Zealand freshwater fishes: a natural history and guide , 553 Auckland : Heinemann Reed .
- McEwan AJ 2009 . Fine scale spatial behaviour of indigenous riverine fish in a small New Zealand stream . Unpublished MSc thesis . Palmerston North, New Zealand, Massey University . 89
- McEwan AJ , Joy MK in press . Habitat use of redfin bullies (Gobiomorphus huttoni) in a small upland stream in Manawatu, New Zealand . Environmental Biology of Fishes .
- McEwan , AJ and Joy , MK . 2011 . Monitoring a New Zealand freshwater fish community using passive integrated transponder (PIT) technology: lessons learned and recommendations for future use . New Zealand Journal of Marine and Freshwater Research , 45 : 137 – 149 . doi: 10.1080/00288330.2010.541925
- Matheney , MP and Rabeni , CF . 1995 . Patterns of movement and habitat use by hog suckers in an Ozark stream . Transactions of the American Fisheries Society , 124 : 886 – 897 . doi: 10.1577/1548-8659(1995)124%3C0886:POMAHU%3E2.3.CO;2
- Matthews , WJ . 1986 . Fish faunal structure in an Ozark stream: stability, persistence and a catastrophic flood . Copeia , 1986 : 388 – 397 . doi: 10.2307/1444997
- Matthews , WJ . 1998 . Patterns in freshwater fish ecology , 756 London : Chapman and Hall .
- Meffe , GK . 1984 . Effects of abiotic disturbance on coexistence of predator-prey fish species . Ecology , 65 : 1525 – 1534 . doi: 10.2307/1939132
- Nesler , TP , Muth , RT and Wasowicz , AF . 1988 . Evidence for baseline flow spikes as spawning cues for Colorado Squawfish in the Yampa River, Colorado . American Fisheries Society Symposium , 5 : 68 – 79 .
- Petts , GE . 1984 . Impounded rivers: perspectives for ecological management , 326 New York : John Wiley & Sons .
- Poff , NL , Allan , JD , Bain , MB , Karr , JR , Prestegaard , KL Richter , BD, . 1997 . The natural flow regime: a paradigm for river conservation and restoration . Bioscience , 47 : 769 – 784 . doi: 10.2307/1313099
- Power , ME , Dietrich , WE and Finlay , JC . 1996 . Dams and downstream aquatic biodiversity: potential food web consequences of hydrologic and geomorphic change . Environmental Management , 20 : 887 – 895 . doi: 10.1007/BF01205969
- R Development Core Team 2011 . R: A Language and Environment for Statistical Computing . R Foundation for Statistical Computing. http://www.R-project.org (accessed 3 March 2012) .
- Ross , ST and Baker , JA . 1983 . The response of fishes to periodic spring floods in a southeastern stream . American Midland Naturalist , 109 : 1 – 14 . doi: 10.2307/2425509
- Turner , TF , Trexler , JC , Miller , GL and Toyer , KE . 1994 . Temporal and spatial dynamics of larval and juvenile fish abundance in a temperate floodplain river . Copeia , 1994 : 174 – 183 . doi: 10.2307/1446683
- Wentworth , CK . 1922 . A scale of grade and class terms for clastic sediments . Journal of Geology , 30 : 377 – 392 . doi: 10.1086/622910
- Winterbourn , MJ , Rounick , JS and Cowie , B . 1981 . Food resources and ingestion patterns of insects along a West Coast, South Island, river system . New Zealand Journal of Marine and Freshwater Research , 18 : 379 – 388 . doi: 10.1080/00288330.1984.9516058
- Yoon , JD , Jang , MH and Joo , GJ . 2011 . Effect of flooding on fish assemblages in small streams in South Korea . Limnology , 12 : 197 – 203 . doi: 10.1007/s10201-010-0337-x
- Young , PS , Chech , JJ Jr and Thompson , LC . 2011 . Hydropower-related pulsed-flow impacts on stream fishes: a brief review, conceptual model, knowledge gaps, and research needs . Reviews in Fish Biology and Fisheries , 21 : 713 – 731 . doi: 10.1007/s11160-011-9211-0