Abstract
Plankton includes the primary producers and consumers that are critical for healthy ecosystem function in the marine realm. My objective was to identify the major contributions of the Leigh Marine Laboratory to our broader understanding of planktonic assemblages. Significant contributions were made prior to 1990 on the ecology of larval fishes. From the 1990s the focus changed to the sensory biology of larval invertebrates and fishes, with a strong emphasis given to the role of reef sound in attracting potential settlers. Both early and post 1980s research has been highly influential in a paradigm shift away from passively drifting larvae that have little control over their fate. Important contributions have also been made on the dynamics of nutrient–phytoplankton interactions, larval development and aquaculture. Opportunities abound for future research on the dynamics of planktonic assemblages in shelf waters and in changing seas.
Introduction
Plankton is the trophic base of pelagic ecosystems and has a critical role in the cycling of atmospheric CO2 and important nutrients (Chisholm et al. Citation2001). Macronutrients and trace elements support autotrophs that range in size from picoplankton to large diatoms and dinoflagellates (e.g. Finkel et al. Citation2010). There is considerable recycling of nutrients through the microbial loop (Fenchel Citation1988). Heterotrophs depend on autotrophs where energy and nutrients are passed through multiple levels of consumers from tiny organisms, that are part autotroph/part heterotroph, to obligate heterotrophs that include microplankton, meso and macroplankton as well as larger nekton (i.e. planktivorous fishes and invertebrates to macrofauna such as tuna, sharks and whales). Some plankton spend their entire life history in the plankton (sensu holoplankton), while some are there only as larvae (sensu meroplankton). Most organisms that inhabit soft and hard benthic environments as juveniles and adults have planktonic life stages. In temperate reef environments, for example, ‘habitat formers’ such as kelp plants have spores which disperse in the plankton (habitats classification, sensu Jones & Andrew Citation1993), and ‘habitat determiners’ or ‘habitat responders’ such as sea urchins, ascidians and many fishes have larvae and spend hours to weeks in the plankton as larvae. Knowledge of life cycles is critical to understanding population dynamics, and this is exemplified by jellyfishes that have benthic and pelagic components to their life history with polypoid benthic stages and pelagic medusoid stages. Moreover, they have sexual reproduction and multiple modes of sexual reproduction (Arai Citation1997). A major paradigm in plankton biology is that most larvae die before they have completed a larval phase, and estimates of 5%–10% mortality per day are common (Cushing Citation1975); 50% of larvae of some species are lost before the yolk is absorbed (Fortier & Leggett Citation1984, Citation1985). Historically, key questions in larval studies are: how do larvae disperse; how are they distributed both horizontally and vertically; and are they just passive particles?
My objective is to review research on planktonic assemblages carried out at the Leigh Marine Laboratory (LML) in the broad context of the acquisition of knowledge and contributions to paradigm change.
It was not my intention to list all pelagic paradigms, as that would be beyond the scope of a short review. I have, therefore, focused on areas that have been addressed at least in part at the LML, and for some there are clear and significant contributions to paradigm shifts. The approach I have taken is to give a brief background to the initial building blocks of plankton research at the LML; here, I reflect on the strong influence of quantitative benthic ecology at Leigh. A summary of papers of high impact follows. My measure of the LML's impact is based on citations by publication and the contributions of clusters of papers toward major paradigms of planktonic assemblages in pelagic ecosystems. Focus is then given to LML and related international contributions in the following areas:
| • | innovative methods and sampling designs for studying plankton, and an increasing appreciation of the physical drivers of plankton distributions with an emphasis on fine-scale features (e.g. internal waves); | ||||
| • | descriptions of the development of larvae, and their condition and growth, both from rearing and field-related studies. Some of this work was carried out in the context of aquaculture and overlaps with the following area of research: | ||||
| • | an enhanced appreciation that fish larvae, in particular, are not passive particles has come from a greater knowledge of larval sensory capabilities and swimming abilities. Daily increments in otoliths have also allowed estimates of the time spent in the plankton; and | ||||
| • | a broader understanding of bottom-up and top-down phenomena based on the concurrent studies of the continental shelf of Northland, New Zealand and adjacent to the LML. Here, I comment on contemporaneous studies by other research groups in Northland (e.g. the National Institute for Water and Atmospheric Research [NIWA]). | ||||
Finally, I comment on growing contributions and opportunities for future research on plankton, focusing on developing areas such as: ecophysiological traits of phytoplankton and our understanding and predictions of pelagic food chains in warming and increasingly acid seas; expanding on historical platforms; collaboration with other institutions (e.g. NIWA) and the vulnerability of losing specialist skills required for the study of plankton, especially taxonomy.
Background
The LML was built in 1962, located on the northeastern side of New Zealand in temperate to subtropical waters (36°16 S; 174°47 E). Located at the boundary of the inner and outer Hauraki Gulf, Leigh is about 70 km from the shelf break (). Early research on plankton at Leigh, and by Fisheries research, initially focused on the identification of holoplankton (phytoplankton; e.g. Taylor Citation1974a, Citationb) and meroplankton (e.g. Cassie Citation1956) (). Jillett (Citation1971) published a memoir on seasonal variation in plankton abundance and physical conditions by season (e.g. temperature) in the Hauraki Gulf and Jellicoe Channel near Leigh.
Figure 1 Map of coastal waters and the shelf break within c. 100 km of LML on the east coast of New Zealand. The star indicates the Jellicoe Channel site used by Jillett (Citation1971) in his plenary work on seasonality of plankton. The majority of work on plankton took place near to the LML, some studies extended to Great Barrier Island and work on the interactions between planktivorous reef fishes and plankton was done at the Poor Knights Islands. Research by New Zealand Fisheries and NIWA (especially by J Zeldis) and others has encompassed the Hauraki Gulf and outer Hauraki Gulf (north of the star). Oceanography of the Gulf is dominated by wind effects and periodic flows of the East Auckland Current (Zeldis et al. Citation2004). The Okakari Point to Cape Rodney Leigh Marine Reserve extends ~2.5 km either side of LML.
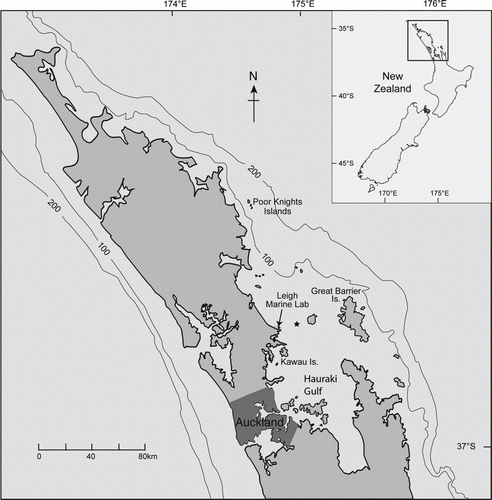
Table 1 Examples of studies on the identification and distribution of plankton with a brief explanation of the focus of the study, location and the sampling design.
The identification of many planktonic groups in New Zealand was based on publications from Australia (e.g. Dakin & Colefax Citation1940) and Europe (e.g. Rose Citation1970), with a small number of local descriptions on specific taxa (e.g. Robertson Citation1975; Bradford-Grieve & Jillett Citation1980; Bradford et al. Citation1983). A series of cruises by Baker (Citation1972) and Crossland (Citation1980, Citation1981, Citation1982) were completed in the inner and sometimes outer Hauraki Gulf. Samples were generally taken at grid stations, or more haphazardly located stations, and data were presented on the egg and larval abundance of multiple species of fish. In some cases, larval descriptions were made and these were augmented by a number of unpublished guides and theses (Kingsford Citation1985; review and illustrations in Kingsford Citation1988).
The focus of research and impact
The research reputation of the LML has generally focused on the ecology of rocky intertidal and subtidal reefs (e.g. Ayling Citation1981; Andrew & Choat Citation1982), and research that was unrelated to plankton predominated. This quantitative approach to ecology influenced the approach that was taken to study plankton and the types of questions that were asked. For example, while Choat and Kingett (Citation1982) estimated the influence of predation by small snapper (Pagrus auratus) on the algal invertebrate fauna a similar type of study was done on planktivorous fishes and plankton by Kingsford and MacDiarmid (Citation1988). In this case it was demonstrated that large aggregations of planktivorous fishes at the Poor Knights Islands (, ) had a measurable effect on the abundance and composition of mesoplankton. Plankton was more abundant upcurrent of aggregations of fish than within or downstream of aggregations. Sampling at the same locations at night did not detect low concentrations of zooplankton where diurnal planktivores had earlier fed.
Figure 2 Planktivorous fish, Caprodon longimanus, feeding in Northern Arch, Poor Knights Islands. Kingsford and MacDiarmid (Citation1988) demonstrated that large aggregations of fish could reduce densities of plankton by up to 5×that of areas without fish. Photo: MJ Kingsford.

The ability to work on plankton was partly limited by the size of vessels at the disposal of staff and students. Unless collaborations were done with Fisheries, or the Oceanographic Institute of New Zealand (e.g. Foster & Battaerd Citation1985), sampling was undertaken from the shore (e.g. Martin & Foster Citation1986) or from small boats (Taylor & Taylor Citation1985). During the late 1970s the only boats were 3.7 m dinghies (e.g. Roper Citation1986). The capability to study plankton was enhanced with a 5 m Avon inflatable that was used to deploy a plankton mesh purse seine net (Kingsford & Choat Citation1985). Although modest in size it gave researchers an opportunity for a ‘between the grid lines’ approach (; e.g. the study of drifting algae [Kingsford Citation1992] and how it can affect the distribution of meroplankton). The Avon was also used to investigate the influence of small-scale oceanographic features (e.g. the slicks of internal waves) on the distribution and movements of plankton (Kingsford & Choat Citation1986). In 1982 the 12 m R. V. Proteus was launched and this allowed larger ichthyoplankton nets to be towed to the side of the vessel (Kingsford Citation1988; Tricklebank Citation2000) or oblique tows to depths of up to 60 m (Kingsford & Choat Citation1989). The most recent vessel, the 14.9 m R. V. Hawere, was launched in 2001 as a multi-task platform that could give quick access to the offshore islands.
Table 2 Examples of between the grid lines research from LML.
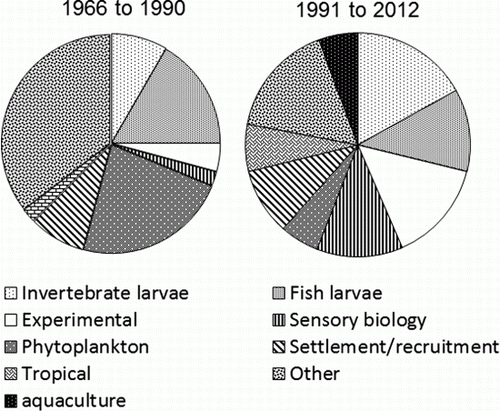
The output of publications and the type of research at LML has greatly changed with time. From 1966 to 1970 only 19 papers were published; but, by the early 2000s, there were over 100 papers per 5-year period (). Publications on plankton made up 6%–23% of total publications in a 5-year period. The greatest contributions have been made in the areas of the ecology of larval fishes, and the sensory biology and development of fishes (). A classification of publications into broad categories before and after 1990 demonstrated great changes in the type of plankton research that was done over the 50 year history of the LML. The proportion of papers on the ecology of fish larvae dropped, and research on the sensory biology of invertebrate and vertebrate larvae, and on aquaculture increased after 1990. Research on invertebrate larvae (e.g. crab megalopae) and experimental work increased with time, and there was greater engagement of staff and students in tropical research that was seeded from LML, especially on the Great Barrier Reef (GBR). This phase-shift in emphasis created new opportunities but, in some cases, bypassed the original research building blocks. The substantial ‘other’ category in 1966 to 1990 included general plankton studies, taxonomy and other minor contributions.
Figure 3 The contribution of plankton-related publications to the total LML publications from 1966 to 2012 (5 year bins).
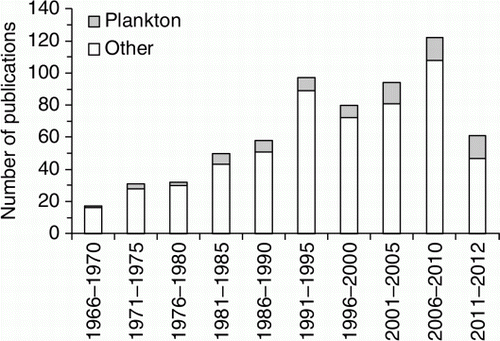
Figure 4 Research categories covered by plankton publications 1966 to 1990, and 1991 to 2012; a single paper could score more than once by category.
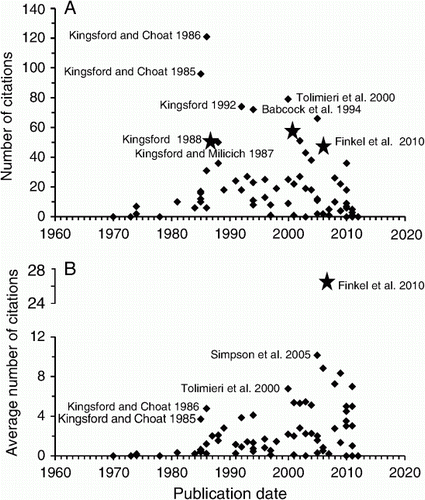
The impact of papers has varied greatly (). Not unexpectedly, taxonomy papers were generally poorly cited. The two most cited papers on plankton are those of Kingsford and Choat (Citation1985, Citation1986); the first, an experimental study on drift algae, presettlement fishes and invertebrates, and the second on the influence of the surface slicks of internal waves on the distribution and transport of presettlement fishes and plankton. With the exception of Babcock's work on coral spawning in Western Australia, the papers fall into three general categories: reviews; presettlement fishes; and invertebrate distributions as influenced by drift algae and oceanography and sensory biology of larvae. Kingsford (Citation1988) reviewed research on the early life history of fishes, Montgomery et al. (Citation2001, Citation2006) on active habitat selection by presettlement fishes and Finkel et al. (2010) on phytoplankton in a changing world. Papers of high impact for sensory biology were Tolimieri et al. (Citation2000) with their ground-breaking work on sound as a cue for navigation. Simpson et al. (Citation2005) consolidated this position of leadership with similar work on a tropical setting – Lizard Island (GBR). Although some of the old papers are still getting four to five citations per year, some more recent papers (date, cites per year) greatly exceed this rate as follows: Finkel et al. (Citation2010; 26.5); Simpson et al. (Citation2005; 10.1).
Figure 5 A, Total citations by year for LML plankton publications. B, Average number of citations by year for LML plankton-related publications. Source: Web of Knowledge, November 2012.
External benchmarking with the other major marine laboratory in New Zealand, Portobello Marine Laboratory (University of Otago) indicated that LML has been competitive with respect to publications on plankton. Portobello had maximum plankton-related outputs of about 15 papers within a 5-year period (especially relationships between oceanography and plankton and aquaculture/developmental biology). Most cited papers include (date, total citations); Wing et al. Citation1995a; 138, Citation1995b; 145) on the transport and settlement of benthic invertebrates, Zeldis and Jillett's (Citation1982; 89) aerial views of Munida gregaria aggregation in fronts and internal waves, McGehee (Citation1998; 78) on acoustic scattering layers; Byrne and Barker (Citation1991; 52), Lamare and Barker (Citation1999; 57) on larval development. In contrast to the LML metrics I could not find any high impact reviews from Portobello.
Innovation in approaches to sampling plankton and oceanographic features
Innovation in methodology and approaches to the study of plankton has resulted in paradigm shifts worldwide. Direct observations of gelatinous zooplankton (Hamner et al. Citation1975) and aggregate material (e.g. Silver et al. Citation1978; Alldredge & Gotschalk Citation1989) greatly increased our understanding of the role of delicate zooplankton and aggregate material that would have previously gathered as an amorphous mass in the cod-end of plankton nets. Towed particle counters (e.g. Suthers et al. Citation2004) and video-based identification have allowed continuous data (rather than discrete data from nets) to be collected among water masses and clear oceanographic features such as gyres. Some methods of antiquity prevail. For example, continuous plankton records from Longhurst Hardy Plankton Recorders (LHPR) are towed on ships of opportunity and have been since the 1930s. As a result they have been the sentinels of climate change (e.g. Richardson & Schoeman Citation2004).
New approaches from LML contributed to changes in our view of larval distributions and behaviour. For example, the presence of larval aggregations in complex near-shore environments, inaccessible to nets, were identified by direct observation on snorkel or scuba (Kingsford & Choat Citation1989; also see Brietburg Citation1991). These studies suggested that some taxa may not disperse far from natal reefs and/or could have a prolonged phase as a ‘potential settler’. Avoidance of plankton nets by agile plankton such as presettlement fishes blinkered our view of what was happening in the plankton. Light traps (Doherty Citation1987), purse seines made of plankton mesh (Leigh; Kingsford & Choat Citation1986) and very large multiple opening closing nets (MOCNESS; Huebert et al. Citation2011) allowed a greater level of accuracy in determining temporal and spatial variation in the abundance of late stage presettlement reef fishes.
Spatial scales of variation in plankton abundance have been the essence of pelagic ecology, and there are strong relationships between oceanographic features of different spatial scales and the distribution of plankton (reviews in Haury et al. Citation1978; Kingsford & Murdoch Citation1998; Mann & Lazier Citation2003). A hierarchical understanding of oceanographic features and their relative importance for influencing plankton distributions is essential. Small-scale phenomena such as internal waves, wind driven circulations and topographic effects can be of great biological importance for determining distributions and for transport (Zeldis & Jillett Citation1982; Pineda Citation1991; Pineda et al. Citation2007). Stratified sampling in and out of the slicks of internal waves near Leigh demonstrated aggregation and the movement of larvae toward coastal environments (Kingsford & Choat Citation1986). Further, variation in the accumulation of drifting macroalgae and associated fishes near shore was attributed to wind and transport by internal waves (Kingsford Citation1992). Data on the velocity of internal waves was presented by Kingsford and Choat (Citation1986) and further data on internal tide activity was provided by Sharples et al. (Citation2001).
Larval development and aquaculture
There have been significant contributions from LML in the areas of aquaculture and larval development (). Research on Pagrus (=Chrysophrys) auratus (Sparidae) has included larval identification, larval development (including senses) and early growth (Pankhurst et al. Citation1991; Pankhurst Citation1994). Important work has also been done on the factors influencing first feeding in kingfish larvae (e.g. Carton Citation2005; Carton & Vaughan Citation2010), digestion and growth in lobster larvae (Simon & Jeffs Citation2011) as well as the larvae of green lipped mussels and patterns of settlement (e.g. Fitzpatrick et al. Citation2011) ().
Table 3 Examples of research on aquaculture from the LML aquaculture; measures of impact are also provided.
Assessments of holoplankton and meroplankton quality have had significant impacts on the questions that could be addressed concerning the feeding environments that plankton encounter. In short, the health of larvae will influence survival rates (Purcell Citation1985). The condition of larvae and relative growth rates have been measured using morphometrics, total lipid and classes of lipid (e.g. Fraser Citation1989; Westerman & Holt Citation1994) as well as RNA:DNA ratios and the widths of daily increments in otoliths (Suthers et al. Citation1989); the latter is used as a proxy for growth. There are few studies from LML that have focused on quality-related assessments of larvae. The spacing of increments was assumed to be a proxy for growth in presettlement monacanthids by Kingsford and Milicich (Citation1987) and this assumption was tested experimentally by Milicich and Choat (Citation1992). Sim-Smith et al. (Citation2012) used increment spacing as a proxy of growth and found that snapper that grew fast as larvae and had a short planktonic larval duration (PLD) of ≤20 d generally grew faster as settled juveniles. Kingsford and Atkinson (Citation1994) suggested that circuli in the scales of recently settled snapper could provide information on growth late in the presettlement phase and be a predictor of behavioural changes of larvae. Lipids (especially triglyceride) were used as a measure of condition of snapper larvae in the unpublished thesis of Atkinson (Citation1987).
How long are fish larvae in the plankton?
The end of the larval stage for pelagic species is when they metamorphose into the form of a juvenile. In contrast, in the case of taxa that are benthic as juveniles and adults, the event of ‘settlement’ (adapted from the study of invertebrate larvae, e.g. Rodriguez et al. Citation1993) marks the end of the presettlement (sensu Kingsford & Milicich Citation1987) or planktonic phase. The duration of this phase has been referred to as the PLD (Victor Citation1991). In the case of fish, this can be estimated from daily increments in otoliths and a change in the spacing of increments that coincides with the time of settlement. Validation that increments are deposited daily in otoliths has been demonstrated for pelagic juvenile monacanthids (Meuschenia (=Parika) scaber; Kingsford & Milicich Citation1987) and larval and juvenile sparids (P. auratus; Kingsford & Atkinson Citation1994). PLDs were estimated as c. 22–44 d in M. scaber and c. 20–30 d in P. auratus (Sim-Smith et al. Citation2012).
Larvae are not passive particles: major international paradigm shift
The primary contributions to global paradigm shifts have been in the areas of larval reef fish biology and ecology, and the sensory biology of fish larvae and invertebrates. Historically, reefs were considered to be ‘open systems’ where larvae were thought to disperse widely and there was little likelihood of larvae returning to the reef from which they were spawned (=natal reef). This view was partly based on a lack of research on the larvae of reef fishes during the 1960s and 70s. Comprehensive work had been done on high latitude larvae by Blaxter (Citation1986) and others. The larvae of clupeids, gadiformes and pleuronectids generally developed slowly and took many days to gain notable swimming abilities, to see further than their own body length, and to gain resilience to starvation. Small wonder that, despite early contributions on the distributions of tropical larval reef fishes (e.g. Leis & Goldman Citation1984), models that predicted dispersal and likely locations for settlement treated larvae as passive particles in mainstream currents (e.g. Williams et al. Citation1984). The mismatch between what was caught in nets, and the size and form of new settlers arriving on reefs was clear.
The term ‘larvae’ implied poorly-formed fish of limited swimming abilities. This did not match what was caught off Leigh (i.e. large pelagic juveniles of fishes such as monacanthids, tripterygiids and mullids that looked very similar to the fishes caught on reefs). The use of plankton-meshed purse nets (Kingsford & Choat Citation1985; Kingsford & Murdoch Citation1998) to encircle drift algae, and stratified sampling in and out of convergence zones resulted in catches of these fishes. In parallel with this work, light traps that were pioneered in Australia (Doherty Citation1987) caught pelagic juveniles of many taxa that resembled the form of newly-settled juveniles.
It was also clear from underwater observations that some larvae, such as tripterygiids, may spend a considerable part of their presettlement phase near suitable settlement habitat (Kingsford & Choat Citation1989). High catches of tripterygiid larvae have been obtained kilometres from shore (e.g. Kingsford & Choat Citation1986; Tricklebank et al. Citation1992). These larvae have a PLD of about three months and if they encounter suitable habitat during this period then they have the swimming abilities to stay close to shore. Larvae of the Gobiesocidae were also found very near shore (in less than 2 m of water) and given the small size difference between the size at which they hatch and settlement it is likely they never move far from their natal reefs (Kingsford & Choat Citation1989).
The larvae of different species were clearly doing different things (Kingsford Citation1988). The monacanthid Meuschenia scaber was found throughout the water column as larvae. Once they reached a juvenile form they were attracted to floating projects such as drift algae in surface waters. Mullids (Upenichthys lineatus) were found in surface waters as larvae and pelagic juveniles. Moreover, aggregations of juveniles were often found around drift algae. In contrast, snapper (Pagrus auratus) were found throughout the water column through development and may be found in concentrations where there are clear thermoclines, while the aforementioned gobiesocids probably don't move far from natal reefs. Although swimming speeds of these taxa were not estimated it was clear from observing larvae swimming near reefs or in purse seines that they have strong swimming abilities.
A series of papers on tropical reef fishes in Australia (Stobutzki & Bellwood Citation1994, Citation1997; Fisher & Bellwood Citation2001; Fisher & Wilson Citation2004) demonstrated that presettlement fishes could swim for long periods of time in waters that were flowing at 35 cm/s. Great differences in performance were found among taxa, and swimming speed increased greatly with size of fish, especially after flexion of the notochord to form a caudal fin and while fish were in the form of a pelagic juvenile. Swimming was one part of the discovery process, but what senses determine the direction of swimming? Directional swimming had been demonstrated in salmon and other taxa, but the literature had historically focused on juveniles and adults (review in Kingsford et al. Citation2002). A range of senses had been suggested including the detection of smell, sound, vibration, visual cues, magnetism, acceleration, electricity and sun compass; many of which influence the orientation of birds and other taxa.
Contributions from LML have been pivotal in terms of demonstrating how presettlement fishes and invertebrates can respond to environmental stimuli, particularly sound (). Tolimieri et al. (Citation2000) used light traps, some that emitted sounds (recordings of reef noises), while others were controls. Tripterygiids, in particular, were about four times as likely to be attracted to traps with sound compared to those without. This discovery was then tested in the tropics by Simpson et al. (Citation2005), who demonstrated that damselfishes (Pomacentridae) and cardinalfishes (Apogonidae) were more likely to be attracted to high frequency or low frequency noises than to controls. In situ measurements of the distances that reef sounds could potentially attract presettlement organisms to reefs were calculated (Radford et al. Citation2011); estimates of up to 4 km can allow modellers to include larval behaviour in their models.
Table 4 Examples of investigations on the sensory biology and behaviour of larvae.
Jeffs et al. (Citation2003) and Stanley et al. (Citation2010, Citation2011) demonstrated that crab megalopae are also attracted to sound, and that favourable sound can speed up metamorphosis into the form of a juvenile crab. Most recently, sound has been shown to influence the settlement of mussels (Wilkens et al. Citation2012). Other significant contributions from LML have been descriptions of changes in the visual capabilities of both invertebrate larvae (Mishra et al. Citation2007) and fish larvae (Pankhurst Citation1994). Furthermore, Radford et al. (2012) have demonstrated that larval snapper can use olfactory cues to orient to suitable settlement habitats such as sea grass beds. The next important step in this research will be to determine the importance of sound relative to other cues such as smell, sun compass and others. Furthermore, how does the importance of different senses change with distance from the reef? For example, presettlement forms may be attracted to reefs by sound and olfaction, but the choice of settlement site can depend on the olfactory cues from conspecifics and habitat (Kingsford et al. Citation2002).
Oceanography and ‘bottom-up’ and ‘top-down’ phenomena on the continental shelf
Pelagic ecosystems are often viewed as ‘bottom-up’ in that the supply of nutrients influences biomass of primary producers, which in turn determines biomass of primary, secondary and tertiary consumers (i.e. from microscopic grazers to the charismatic megafauna such as whales). However, there can also be significant ‘top-down effects’. For example, high densities of predatory cnidarian jellyfishes can have a great effect on small grazers, which in turn affects abundance of producers (e.g. Pitt et al. Citation2007). The grazing activity of salps can also have a great influence on the abundance of microscopic prey (Zeldis et al. Citation1995). Gelatinous zooplankton are considered to be a major causal factor in ‘phase shifts’ of pelagic ecosystems in many parts of the world (e.g. west coast of North America, Brodeur et al. Citation2002; Africa, Lynam et al. Citation2006).
Production in the photic zone has a great impact on the benthos because dead plankton, faeces and aggregations of material (sensu ‘marine snow’; Silver et al. Citation1978) sink to the substratum. This component of ‘benthic-pelagic coupling’ provides deposit feeders with nutrition, in turn supporting a range of other consumers (Graf Citation1989). Other important aspects of benthic–pelagic coupling include biochemical coupling of nutrients from the water column (Marcus & Boero Citation1998; Zeldis Citation2004), resting phases of planktonic organisms that spend part of their life on the substratum and mortality events on the benthos from plankton blooms (Taylor et al. Citation1985). Furthermore, benthic organisms also have links with the water column through suspension feeding and/or larvae that are planktonic. The survival of larvae can be highly dependent on pelagic processes be they bottom-up or top-down. For example, variation in recruitment and subsequent population size may be determined by upwelling-related variation in feeding conditions for larvae (e.g. Navarette et al. Citation2005). In contrast, direct predation on larvae (Moller Citation1984), or competition for suitable larval food (Zeldis et al. Citation1995, Citation2005; Lynam et al. Citation2005), are top-down planktonic processes that can influence survival to settlement out of the plankton.
There have been few contributions from LML on shelf-wide oceanographic processes and variation in plankton dynamics as well as benthic–pelagic coupling; this has been the domain of NIWA. In a series of papers it was demonstrated that warm incursions by the East Auckland Current (EAC) and upwelling events occur on the shelf late in spring and leading into summer (Sharples Citation1997; Sharples & Greig Citation1998). Between early and late summer upwelling generally ceases, and there was a transition to downwelling, due to a combination of forcing by local winds and the influence of the EAC (Zeldis et al. Citation2004). Upwelling caused extensive loading of the inner shelf and Hauraki Gulf bottom waters with nitrate. However, the system moved towards a net heterotrophic system where it was concluded that remineralisation on the shelf and regenerated nutrient supply was important for sustaining stocks of chlorophyll a (Zeldis Citation2004; Bury et al. Citation2012). Furthermore, calculations on the percentage of primary production that was remineralised in sediments of the northern shelf-slope region, Hauraki Gulf and Firth of Thames (13%–34%) were provided by Giles et al. (Citation2007).
Zeldis also published a number of important papers that demonstrated the importance of upwelling and abundance of gelatinous zooplankters on the abundance of snapper larvae (P. auratus). The larvae of this important species suffered highest mortality rates when salps were abundant. They altered the distribution of potential prey or competed for suitable larval food (Zeldis et al. Citation1995, Citation2005). Zeldis and Francis (Citation1998) also used plankton surveys (i.e. the daily egg production method) to estimate the biomass of snapper in the Hauraki Gulf.
Phytoplankton: an area of growth in research
Research on phytoplankton moved from identification to nutrient uptake kinetics (Rees & Allison Citation2006; Rees Citation2007; Beardall et al. Citation2009; Flynn et al. Citation2010). This has received increased attention due to potential climate-induced changes to marine food chains (Finkel et al. Citation2010). In this well-cited review, it was stated that ecophysiological traits in phytoplankton can be used to predict phytoplankton growth rates as well as chemical recycling and changed interactions between producers and consumers and loss processes. This in turn can vary with changes in key environmental factors. There is great potential for the LML to contribute to paradigm shifts in this area.
Other contributions and opportunities
With a focus on sensory biology over the last 10 to 15 years, many larger-scale questions have not been addressed. For example, there are great differences in the distribution and abundance of planktivorous fishes across the continental shelf (Kingsford Citation1989). Offshore islands have a fauna dominated by Chromis dispilus and other taxa such as Caprodon longimanus and Callanthias allporti, whereas Scorpis lineolata is abundant near the mainland. These patterns are reminiscent of debates that have been had in Australia on the factors determining cross-shelf distributions of fish such as: oceanography; plankton abundance and composition; turbidity; and larval connectivity. Benthic feeding fishes also show great variation in abundance across the shelf (e.g. Notolabrus celidotus, Pseudolabrus luculentus, Parma alboscapularis) and new tools for addressing issues, such as connectivity, are available and can provide invaluable information to marine managers (Jones et al. Citation2009; Jones Citation2013).
Marine laboratories generally have many theses with valuable unpublished information; LML is no exception. Cole (Citation1987) worked on the depth stratification of pilchard larvae while Coupe (Citation1993) studied plankton in the Leigh Marine Reserve and Parr (Citation1994) worked on bivalve larvae in Whangateau Harbour. Despite Jillett's (Citation1971) major contribution on the seasonality of plankton there are few other published data. The theses of Kingsford (Citation1980) and MacDiarmid (Citation1981) contain 18 months of valuable plankton data (their seasonal data on Penilia avirostris are graphed in Kingsford & Murdoch Citation1998). Thompson (Citation1983) described seasonal variation in fish larvae near Goat Island. Although seasonal plankton data appear pedestrian, they are critical for understanding plankton trophodynamics at different times of the year. The importance of baselines of plankton composition and abundance is now considered critical in times of changing water temperatures, currents, ocean pH and oxygen profiles (Richardson & Schoeman Citation2004; Brierley & Kingsford Citation2009).
In conclusion, research at the LML has contributed to paradigm shifts in a few discrete areas, but opportunities abound. Aquaculture and research on uptake dynamics of phytoplankton shows promise. The impact of climate change on phytoplankton and the cascade effects that cause broader impacts on marine food chains is a sensible area to progress (e.g. Finkel et al. Citation2010). Patterns that were found pre-1990s have not been pursued. Our knowledge of the senses that larvae use prior to settlement has increased greatly. However, multifactorial experiments are required for a more integrated understanding of the spatial and temporal hierarchy of senses used. New tools (e.g. intergenerational tagging and improved genetics) can allow old questions, such as larval connectivity, to be addressed. Research on cross-shelf phenomena have been in the NIWA domain and value could have been added with more collaborative ventures; the Auckland/NIWA Joint Graduate School in Coastal and Marine Science should help. The rich broth of deep-sea plankton that rises from the shelf break beckons for both taxonomic and ecological investigation.
Acknowledgements
I would like to thank Mark O'Callaghan for assisting in the data mining required to write this review. Furthermore, thanks to Kendall Clements and John Montgomery for organising the collection of reviews in this special issue. I apologise to the authors of any papers that have been missed, but I hope you agree with the thesis of this review. Thanks to John Zeldis and an anonymous referee for helpful comments on the manuscript. I believe telling the stories as we have done reminds us of how far we have come and will realise new opportunities for research. The LML has been a special place for inspiration, camaraderie, unique characters, life-long friendships and opportunity; it has been a privilege to be a part of the Leigh story.
References
- Alfaro AC , Jeffs AG , Creese RG 2004 . Bottom-drifting algal/mussel spat associations along a sandy coastal region in northern New Zealand . Aquaculture 241 : 269 – 290 . doi: 10.1016/j.aquaculture.2004.07.029
- Alfaro AC , McArdle B , Jeffs AG 2010 . Temporal patterns of arrival of beachcast green-lipped mussel (Perna canaliculus) spat harvested for aquaculture in New Zealand and its relationship with hydrodynamic and meteorological conditions . Aquaculture 302 : 208 – 218 . doi: 10.1016/j.aquaculture.2010.02.028
- Alldredge AL , Gotschalk CC 1989 . Direct observations of the mass flocculations of diatom blooms: characteristics, settling velocities and formation of diatom aggregates . Deep Sea Research 36 : 159 – 171 . doi: 10.1016/0198-0149(89)90131-3
- Andrew NL , Choat JH 1982 . The influence of predation and conspecific adults on the abundance of juvenile Evechinus chloroticus (Echinoidea; Echinometridae) . Oecologia 54 : 80 – 87 . doi: 10.1007/BF00541113
- Arai MN 1997 . A functional biology of Scyphozoa . London , Chapman & Hall . 316
- Atkinson MH 1987 . Ontogenetic patterns in presettlement: Chrysophrys auratus (Sparidae) . Unpublished thesis , University of Auckland , New Zealand . 72
- Ayling AM 1981 . The role of biological disturbance in temperate subtidal encrusting communities . Ecology 62 : 830 – 847 . doi: 10.2307/1937749
- Baker AN 1972 . Reproduction, early life history and age-growth relationships of the New Zealand pilchard, Sardinops neopilchardus (Steindachner) . Fisheries Research Bulletin 5 . 64
- Baker CF , Montgomery JC 2001 . Species-specific attraction of migratory banded kokopu juveniles to adult pheromones . Journal of Fish Biology 58 : 1221 – 1229 . doi: 10.1111/j.1095-8649.2001.tb02281.x
- Beardall J , Allen D , Bragg J , Finkel ZV , Flynn KJ , Quigg A , et al. 2009 . Allometry and stoichiometry of unicellular, colonial and multicellular phytoplankton . New Phytologist 181 : 295 – 309 . doi: 10.1111/j.1469-8137.2008.02660.x
- Blaxter JHS 1986 . Development of sense organs and behaviour of teleost larvae with special reference to feeding and predation avoidance . Transactions of the American Fisheries Society 115 : 98 – 114 . doi: 10.1577/1548-8659(1986)115<98:NLFCDO>2.0.CO;2
- Bradford JM , Haakonssen L , Jillett JB 1983 . The marine fauna of New Zealand: pelagic calanoid copepods: families Eucaetidae, Phaennidae, Scolecithricidae, Diaixidae, and Tharybidae . Wellington , New Zealand Oceanographic Institute . 149 .
- Bradford-Grieve JM , Jillett JB 1980 . The marine fauna of New Zealand: pelagic calanoid copepods: family Aetideidae . Wellington , New Zealand, Department of Scientific and Industrial Research . 101
- Breitburg DL 1991 . Settlement patterns and presettlement behavor of the naked goby, Gobiosoma bosci, a temperate oyster reef fish . Marine Biology 109 : 213 – 221 . doi: 10.1007/BF01319389
- Brierley AS , Kingsford MJ 2009 . Impacts of climate change on marine organisms and ecosystems . Current Biology 19 : R602 – R614 . doi: 10.1016/j.cub.2009.05.046
- Brodeur RD , Sugisaki H , Hunt JGL 2002 . Increases in jellyfish biomass in the Bering Sea: implications for the ecosystem . Marine Ecology Progress Series 233 : 89 – 103 . doi: 10.3354/meps233089
- Bury SJ , Zeldis JR , Nodder SD , Gall M 2012 . Regenerated primary production dominates in a periodically upwelling shelf ecosystem, northeast New Zealand . Continental Shelf Research 32 : 1 – 21 . doi: 10.1016/j.csr.2011.09.008
- Byrne M , Barker MF 1991 . Embryogenesis and larval development of the asteroid Patiriella regularis viewed by light and scanning electron-microscopy . Biological Bulletin 180 : 332 – 345 . doi: 10.2307/1542335
- Carton AG 2005 . The impact of light intensity and algal-induced turbidity on first-feeding Seriola lalandi larvae . Aquaculture Research 36 : 1588 – 1594 . doi: 10.1111/j.1365-2109.2005.01383.x
- Carton AG , Vaughan MR 2010 . Behavioural and anatomical measures of visual acuity in first-feeding yellowtail kingfish (Seriola lalandi) larvae . Environmental Biology of Fishes 89 : 3 – 10 . doi: 10.1007/s10641-010-9657-2
- Cassie RM 1956 . Early development of the snapper, Chrysophrys auratus Forster . Transactions of the Royal Society of New Zealand 83 : 705 – 713 .
- Chisholm SW , Falkowski PG , Cullen JJ 2001 . Dis-crediting ocean fertilization . Science 294 : 309 – 310 . doi: 10.1126/science.1065349
- Choat JH , Kingett PD 1982 . The influence of fish predation on the abundance cycles of an algal turf invertebrate fauna . Oecologia 54 : 88 – 95 . doi: 10.1007/BF00541114
- Cole RG 1987 . Distribution and abundance of clupeoid larvae in the Hauraki Gulf . Unpublished PhD thesis , University of Auckland , New Zealand . 67
- Coupe NM 1993 . The variability of zooplankton, at the Leigh Marine Reserve . Unpublished MSc thesis . University of Auckland , New Zealand . 161 p.
- Crossland L 1980 . The number of snapper, Chrysophrys auratus (Foster) in the Hauraki Gulf, New Zealand, based on egg surveys in 1974–75 and 1975–76 . Fisheries Research Bulletin 22 . 39
- Crossland L 1981 . Fish eggs and larvae of the Hauraki Gulf, New Zealand . Fisheries Research Bulletin 23 . 62
- Crossland J 1982 . Movements of tagged snapper in the Hauraki Gulf . Fisheries Research Division Occasional Publication 35 : 1 – 15 .
- Cushing D 1975 . Marine ecology and fisheries . Cambridge , , UK , Cambridge University Press . 278
- Dakin WJ , Colefax AN 1940 . The plankton of the Australian coastal waters off New South Wales . Part I . Sydney , Australian Publishing Company Ltd . 215
- Doherty PJ 1987 . Light-traps: selective but useful devices for quantifying the distributions and abundance of larval fishes . Bulletin of Marine Science 41 : 423 – 431 .
- Fenchel T 1988 . Marine plankton and food chains . Annual Review of Ecology and Systematics 19 : 19 – 38 . doi: 10.1146/annurev.es.19.110188.000315
- Finkel ZV , Beardall J , Flynn KJ , Quigg A , Rees TAV , Raven JA 2010 . Phytoplankton in a changing world: cell size and elemental stoichiometry . Journal of Plankton Research 32 : 119 – 137 . doi: 10.1093/plankt/fbp098
- Fisher R , Bellwood DR 2001 . Effects of feeding on the sustained swimming abilities of late-stage larval Amphiprion melanopus . Coral Reefs 20 : 151 – 154 . doi: 10.1007/s003380100149
- Fisher R , Wilson SK 2004 . Maximum sustainable swimming speeds of late stage larvae of nine species of reef fishes . Journal of Experimental Marine Biology and Ecology 312 : 171 – 186 . doi: 10.1016/j.jembe.2004.06.009
- Fitzpatrick MP , Jeffs AG , Dunphy BJ 2011 . Efficacy of calcein as a chemical marker of green-lipped mussel (Perna canaliculus) larvae and its potential use for tracking larval dispersal . Aquaculture Research . doi: 10.1111/j.1365–2109.2011.03034.x
- Flynn KJ , Raven JA , Rees TAV , Finkel Z , Quigg A , Beardall J 2010 . Is the growth rate hypothesis applicable to microalgae? Journal of Phycology 46 : 1 – 12 . doi: 10.1111/j.1529-8817.2009.00756.x
- Fortier L , Leggett WC 1984 . Small-scale covariability in the abundance of fish larvae and their prey . Canadian Journal of Fisheries and Aquatic Sciences 41 : 502 – 512 . doi: 10.1139/f84-060
- Fortier L , Leggett WC 1985 . A drift study of larval fish survival . Marine Ecology Progress Series 25 : 245 – 257 . doi: 10.3354/meps025245
- Foster BA , Battaerd WR 1985 . Distribution of zooplankton in a coastal upwelling in New Zealand . New Zealand Journal of Marine and Freshwater Research 19 : 213 – 226 . doi: 10.1080/00288330.1985.9516088
- Fraser A 1989 . Triacylglycerol content as a condition index for fish, bivalve and crustacean larvae . Canadian Journal of Fisheries and Aquatic Science 46 : 1868 – 1873 . doi: 10.1139/f89-235
- Giles H , Pilditch CA , Nodder SD , Zeldis JR , Currie K 2007 . Benthic oxygen fluxes and sediment properties on the northeastern New Zealand continental shelf . Continental Shelf Research 27 : 2373 – 2388 . doi: 10.1016/j.csr.2007.06.007
- Graf G 1989 . Benthic-pelagic coupling in a deep-sea benthic community . Nature 341 : 437 – 439 . doi: 10.1038/341437a0
- Gribben PE 2005 . Gametogenic development and spawning of the razor clam, Zenatia acinaces in northeastern New Zealand . New Zealand Journal of Marine and Freshwater Research 39 : 1287 – 1296 . doi: 10.1080/00288330.2005.9517393
- Gribben PE , Jeffs AG , de Nys R , Steinberg PD 2011 . Relative importance of natural cues and substrate morphology for settlement of the New Zealand Greenshell™ mussel, Perna canaliculus . Aquaculture 319 : 240 – 246 . doi: 10.1016/j.aquaculture.2011.06.026
- Hamner WM , Madin LP , Alldredge AL , Gilmer RW , Hamner PP 1975 . Underwater observations of gelatinous zooplankton: sampling problems, feeding biology, and behaviour . Limnology and Oceanography 20 : 907 – 917 . doi: 10.4319/lo.1975.20.6.0907
- Haury IR , McGowan JA , Wiebe PH 1978 . Patterns and processes in the time-space scales of plankton distributions . In : Steele J . Spatial pattern plankton communities . New York , Plenum Press . 277 – 337 .
- Huebert KB , Cowen RK , Sponaugle S 2011 . Vertical migrations of reef fish larvae in the Straits of Florida and effects on larval transport . Limnology and Oceanography 56 : 1653 – 1666 . doi: 10.4319/lo.2011.56.5.1653
- Jeffs A , Tolimieri N , Montgomery JC 2003 . Crabs on cue for the coast: the use of underwater sound for orientation by pelagic crab stages . Marine and Freshwater Research 54 : 841 – 845 . doi: 10.1071/MF03007
- Jillet JB 1971 . Zooplankton and hydrology of Hauraki Gulf, New Zealand . Memoirs of the New Zealand Oceanographic Institute 53 : 1 – 103 .
- Jones GP 2013 . Ecology of rocky reef fish of northeastern New Zealand: 50 years on . New Zealand Journal of Marine and Freshwater Research 47 . doi: 10.1080/00288330.2013.812569
- Jones GP , Almany GR , Russ GR , Sale PF , Steneck RS , van Oppen MJH , et al. 2009 . Larval retention and connectivity among populations of corals and reef fishes: history, advances and challenges . Coral Reefs 28 : 307 – 325 . doi: 10.1007/s00338-009-0469-9
- Jones GP , Andrew NL 1993 . Temperate reefs and the scope of seascape ecology . In : Battershill CN , Schiel DR , Jones GP , Creese RG , MacDiarmid AB . Proceedings of the Second International Temperate Reef Symposium, 7–10 January 1992, Auckland . Wellington , NIWA Marine . 252 .
- Kingsford MJ 1980 . Interrelationships between spawning and recruitment of Chromis dispilus (Pices: Pomacentridae) . Unpublished MSc thesis , University of Auckland , New Zealand . 84
- Kingsford MJ 1985 . The demersal eggs and planktonic larvae of Chromis dispilus (Teleostei: Pomacentridae) in north-eastern New Zealand coastal waters . New Zealand Journal of Marine and Freshwater Research 19 : 429 – 438 . doi: 10.1080/00288330.1985.9516107
- Kingsford MJ 1988 . The early life history of fish in coastal waters of northern New Zealand: a review . New Zealand Journal of Marine and Freshwater Research 22 : 463 – 479 . doi: 10.1080/00288330.1988.9516316
- Kingsford MJ 1989 . Distribution patterns of planktivorous reef fish along the coast of northeastern New Zealand . Marine Ecology Progress Series 54 : 13 – 24 . doi: 10.3354/meps054013
- Kingsford MJ 1992 . Drift algae and small fish in coastal waters of northeastern New Zealand . Marine Ecology Progress Series 80 : 41 – 55 . doi: 10.3354/meps080041
- Kingsford MJ , Atkinson MH 1994 . Increments in otoliths and scales: how they relate to the age and early development of reared and wild larval and juvenile snapper Pagrus auratus (Sparidae) . Australian Journal of Marine and Freshwater Research 45 : 1007 – 1021 . doi: 10.1071/MF9941007
- Kingsford MJ , Choat JH 1985 . The fauna associated with drift algae captured with a plankton-mesh purse-seine net . Limnology and Oceanography 30 : 618 – 630 . doi: 10.4319/lo.1985.30.3.0618
- Kingsford MJ , Choat JH 1986 . Influence of surface slicks on the distribution and onshore movements of small fish . Marine Biology 91 : 161 – 171 . doi: 10.1007/BF00569432
- Kingsford MJ , Choat JH 1989 . Horizontal distribution patterns of presettlement reef fish: are they influenced by the proximity of reefs? Marine Biology 100 : 285 – 297 . doi: 10.1007/BF00428124
- Kingsford MJ , Leis J , Shanks A , Lindeman K , Morgan S , Pineda J 2002 . Sensory environments, larval abilities and local self-recruitment . Bulletin of Marine Science . 70(Supplement 1) : 309 – 340 .
- Kingsford MJ , MacDiarmid AB 1988 . Interrelations between planktivorous reef fish and zooplankton in temperate waters . Marine Ecology Progress Series 48 : 103 – 117 . doi: 10.3354/meps048103
- Kingsford MJ , Milicich MJ 1987 . Presettlement phase of Parika scaber (Pisces: Monacanthidae): a temperate reef fish . Marine Ecology Progress Series 36 : 65 – 79 . doi: 10.3354/meps036065
- Kingsford MJ , Murdoch R 1998 . Planktonic assemblages . In : Kingsford MJ , Battershill CN . Studying temperate marine environments: a handbook for ecologists . Christchurch , , New Zealand , Canterbury University Press . 227 – 268 .
- Kingsford MJ , Tricklebank KA 1991 . Ontogeny and behavior of (Aldrichetta forsteri) (Teleostei: Mugilidae) . Copeia 1 : 9 – 16 . doi: 10.2307/1446243
- Lamare MD , Barker MF 1999 . In situ estimates of larval development and mortality in the New Zealand sea urchin Evechinus chloroticus (Echinodermata: Echinoidea) . Marine Ecology Progress Series 180 : 197 – 211 . doi: 10.3354/meps180197
- Leis JM , Goldman B 1984 . A preliminary distributional study of fish larvae near a ribbon coral reef in the Great Barrier Reef . Coral Reefs 2 : 197 – 203 .
- Lynam CP , Gibbons MJ , Axelsen BE , Sparks CAJ , Coetzee J , Heywood BG , et al. 2006 . Jellyfish overtake fish in a heavily fished ecosystem . Current Biology 16 : 492 – 493 . doi: 10.1016/j.cub.2006.06.018
- Lynam CP , Heath MR , Hay SJ , Brierley AS 2005 . Evidence for impacts by jellyfish on North Sea herring recruitment . Marine Ecology Progress Series 298 : 157 – 167 . doi: 10.3354/meps298157
- MacDiarmid AB 1981 . Factors influencing the distribution and abundance of two temperate planktivorous reef fish, Pempheris adspersa and Scorpis violacenus . Unpublished PhD thesis , University of Auckland . 101
- MacDiarmid AB 1985 . Sunrise release of larvae from the palinurid rock lobster Jasus edwardsii . Marine Ecology Progress Series 21 : 313 – 315 . doi: 10.3354/meps021313
- Maddock L , Taylor FJ 1984 . The application of multivariate statistical methods to phytoplankton counts from the Hauraki Gulf, New Zealand . New Zealand Journal of Marine and Freshwater Research 18 : 309 – 322 . doi: 10.1080/00288330.1984.9516053
- Mann KH , Lazier JRN 2003 . Dynamics of marine ecosystems: biological-physical interactions in the ocean. , 3rd edn. Oxford , UK , Blackwell . 486
- Marcus NH , Boero F 1998 . Minireview: the importance of benthic-pelagic coupling and the forgotten role of life cycles in coastal aquatic systems . Limnology and Oceanography 43 : 76 . doi: 10.4319/lo.1998.43.5.0763
- Martin AT , Foster BA 1986 . Distribution of barnacle larvae in Mahurangi Harbour, North Auckland . New Zealand Journal of Marine and Freshwater Research 20 : 67 – 76 . doi: 10.1080/00288330.1986.9516130
- McGehee DE , O'Driscoll RL , Traykovski LVM 1998 . Effects of orientation on acoustic scattering from Antarctic krill at 120 kHz . Deep-Sea Research Part II – Topical Studies in Oceanography 45 : 1273 – 1294 . doi: 10.1016/S0967-0645(98)00036-8
- Milicich MJ , Choat JH 1992 . Do otoliths record changes in somatic growth rate? Conflicting evidence from a laboratory and field study of a temperate reef fish . Australian Journal of Marine and Freshwater Research 43 : 331 – 342 . doi: 10.1071/MF9921203
- Mishra M , Jeffs AG , Meyer-Rochow VB 2007 . Eye structure of the phyllosoma larva of the rock lobster Jasus edwardsii: how does it differ from that of the adult? Invertebrate Reproduction and Development 49 : 213 – 222 . doi: 10.1080/07924259.2006.9652209
- Moller H 1984 . Reduction of a larval herring population by a jellyfish predator . Science 224 : 621 – 622 . doi: 10.1126/science.224.4649.621
- Montgomery JC , Tolimieri N , Haine OS 2001 . Active habitat selection by pre-settlement reef fishes . Fish and Fisheries 2 : 261 – 277 . doi: 10.1046/j.1467-2960.2001.00053.x
- Montgomery JC , Jeffs A , Simpson SD , Meekan M , Tindle C 2006 . Sound as an orientation cue for the pelagic larvae of reef fishes and decapod crustaceans . In : Southward AJ , Sims DW . Advances in marine biology , 51 . London , Academic Press Ltd–Elsevier Science Ltd . 143 – 196 .
- Navarrete SA , Wieters EA , Broitman BR , Castilla JC 2005 . Scales of benthic-pelagic and the intensity of species interactions: from recruitment limitation to top-down control . Proceedings of the National Academy of Sciences of the USA 102 : 18046 – 18051 . doi: 10.1073/pnas.0509119102
- Pankhurst PM 1994 . Age-related-changes in the visual-acuity of larvae of New Zealand snapper, Pagrus auratus . Journal of the Marine Biological Association of the United Kingdom 74 : 337 – 349 . doi: 10.1017/S0025315400039370
- Pankhurst PM , Montgomery JC , Pankhurst NW 1991 . Growth, development and behavior of artificially reared larval Pagrus auratus (Bloch and Schneider, 1801) (Sparidae) Australian Journal of Marine and Freshwater Research 42 : 391 – 398 . doi: 10.1071/MF9910391
- Parr SW 1994 . Bivalve larvae in the Whangateau Harbour . Unpublished MSc thesis . University of Auckland , New Zealand . 102 p.
- Pineda J 1991 . Predictable upwelling and the shoreward transport of planktonic larvae by internal bores . Science 253 : 548 – 551 . doi: 10.1126/science.253.5019.548
- Pineda J , Hare JA , Sponaugle S 2007 . Larval transport and dispersal in the coastal ocean and consequences for population connectivity . Oceanography 20 : 22 – 39 . doi: 10.5670/oceanog.2007.27
- Pitt KA , Kingsford MJ , Rissik D , Koop K 2007 . Jellyfish modify the response of planktonic assemblages to nutrient pulses . Marine Ecology Progress Series 351 : 1 – 13 . doi: 10.3354/meps07298
- Purcell JE 1985 . Predation on fish eggs and larvae by pelagic cnidarians and ctenophores . Bulletin of Marine Science 37 : 739 – 755 .
- Radford CA , Stanley JA , Simpson SD , Jeffs AG 2011 . Juvenile coral reef fish use sound to locate habitats . Coral Reefs 30 : 295 – 305 . doi: 10.1007/s00338-010-0710-6
- Rees TAV 2007 . Metabolic and ecological constraints imposed by similar rates of ammonium and nitrate uptake per unit surface area at low substrate concentrations in marine phytoplankton and macroalgae . Journal of Phycology 43 : 197 – 207 . doi: 10.1111/j.1529-8817.2007.00321.x
- Rees TAV , Allison VJ 2006 . Evidence for an extracellular L-amino acid oxidase in nitrogen-deprived Phaeodactylum tricornutum (Bacillariophyceae) and inhibition of enzyme activity by dissolved inorganic carbon . Phycologia 45 : 337 – 342 . doi: 10.2216/04-92.1
- Richardson AJ , Schoeman DS 2004 . Climate impact on plankton ecosystems in the northeast Atlantic . Science 305 : 1609 – 1611 . doi: 10.1126/science.1100958
- Robertson DA 1975 . A key to the planktonic eggs of some New Zealand marine teleosts . Fisheries Research Division Occasional Publication 9 . 19
- Rodriguez SR , Ojeda FP , Inestrosa NC 1993 . Settlement of benthic marine invertebrates . Marine Ecology Progress Series 97 : 193 – 207 . doi: 10.3354/meps097193
- Roper DS 1986 . Occurrence and recruitment of fish larvae in a northern New Zealand estuary . Estuarine, Coastal and Shelf Science 22 : 705 – 717 . doi: 10.1016/0272-7714(86)90094-6
- Rose M 1970 . Copépodes pélagiques . In: Faune de France 26 (Paris, Librairie de la Faculté des Sciences, 1933). In Faune de France 26. Liechtenstein, Nendeln, Kraus Reprint. 374 p .
- Sharples J 1997 . Cross-shelf intrusion of subtropical water into the coastal zone of northeast New Zealand . Continental Shelf Research 17 : 835 – 857 . doi: 10.1016/S0278-4343(96)00060-X
- Sharples J , Greig MJN 1998 . Tidal currents, mean flows, and upwelling on the north-east shelf of New Zealand . New Zealand Journal of Marine and Freshwater Research 32 : 215 – 231 . doi: 10.1080/00288330.1998.9516821
- Sharples J , Moore CM , Abraham ER 2001 . Internal tide dissipation, mixing, and vertical nitrate flux at the shelf edge of NE New Zealand . Journal of Geophysical Research – Oceans 106 : 14069 – 14081 . doi: 10.1029/2000JC000604
- Silver MW , Shanks AL , Trent JD 1978 . Marine snow: microplankton habitat and source of small-scale patchiness in pelagic populations . Science 201 : 371 – 373 . doi: 10.1126/science.201.4353.371
- Simon CJ , Jeffs A 2011 . The effect of dietary carbohydrates on the growth response, digestive gland glycogen and digestive enzyme activities of early spiny lobster juveniles, Jasus edwardsii . Aquaculture Nutrition 17 : 613 – 626 . doi: 10.1111/j.1365-2095.2011.00861.x
- Simpson SD , Meekan M , Montgomery J , McCauley R , Jeffs A 2005 . Homeward Sound . Science 308 : 221 . doi: 10.1126/science.1107406
- Sim-Smith CJ , Jeffs AG , Radford CA 2012 . Variation in the growth of larval and juvenile snapper, Chrysophrys auratus (Sparidae) . Marine and Freshwater Research 63 : 1231 – 1243 . doi: 10.1071/MF12176
- Stanley JA , Radford CA , Jeffs , AG 2010 . Induction of settlement in crab megalopae by ambient underwater reef sound . Behavioral Ecology 21 : 113 – 120 . doi: 10.1093/beheco/arp159
- Stanley JA , Radford CA , Jeffs AG 2011 . Behavioural response thresholds in New Zealand crab megalopae to ambient underwater sound . PLoS ONE 6 ( 12 ): e28572 . doi: 10.1371/journal.pone.0028572
- Stobutzki IC , Bellwood DR 1994 . An analysis of sustained swimming abilities of pre- and post-settlement coral reef fishes . Journal of Experimental Marine Biology and Ecology 175 : 275 – 286 . doi: 10.1016/0022-0981(94)90031-0
- Stobutzki I , Bellwood DR 1997 . Sustained swimming abilities of the late pelagic stages of coral reef fishes . Marine Ecology Progress Series 149 : 35 – 41 . doi: 10.3354/meps149035
- Suthers IM , Frank KT , Campana SC 1989 . Spatial comparison of recent growth in post-larval cod (Gadus morhua) off southwestern Nova Scotia: inferior growth in a presumed nursery area . Canadian Journal of Fisheries and Aquatic Science 46 : 113 – 124 . doi: 10.1139/f89-283
- Suthers IM , Taggart CT , Kelley D , Rissik D , Middleton JH 2004 . Entrainment and advection in an island's tidal wake, as revealed by light attenuance, zooplankton, and ichthyoplankton . Limnology and Oceanography 49 : 283 – 296 . doi: 10.4319/lo.2004.49.1.0283
- Taylor FJ 1970 . A preliminary annotated check list of diatoms from New Zealand coastal waters . Transactions of the Royal Society of New Zealand: Biological Sciences 12 : 153 – 174 .
- Taylor FJ 1973 . Phytoplankton and nutrients in the Hauraki Gulf approaches . In : Fraser R . Oceanography of the South Pacific 1972: Proceedings of the International Symposium on the Oceanography of the South Pacific, Wellington, 9–15 February 1972 . Wellington , , New Zealand , National Commission for UNESCO . 485 – 492 .
- Taylor FJ 1974a . A preliminary annotated check list of dinoflagellates from New Zealand coastal waters . Journal of the Royal Society of New Zealand 4 : 193 – 202 . doi: 10.1080/03036758.1974.10426439
- Taylor FJ 1974b . A preliminary annotated check list of micro-algae other than diatoms and dinoflagellates from New Zealand coastal waters . Journal of the Royal Society of New Zealand 4 : 395 – 400 . doi: 10.1080/03036758.1974.10419383
- Taylor FJ 1978 . Records of marine algae from the Leigh area. Part II. Records of phytoplankton from Goat Island Bay . Tane 24 : 213 – 218 .
- Taylor FJ 1981 . Phytoplankton and nutrients in Goat Island Bay, New Zealand . Internationale Revue der Gesamten Hydrobiologie 66 : 377 – 406 . doi: 10.1002/iroh.19810660310
- Taylor FJ , Durbin EG 1978 . Records of marine algae from the Leigh area. Part III. Phytoplankton from the Whangateau Harbour . Tane 24 : 219 – 222 .
- Taylor NJ , Taylor FJ 1985 . Hydrology and changes in the nutrients and phytoplankton levels in Goat Island Bay, northern New Zealand . Internationale Revue der Gesamten Hydrobiologie 70 : 173 – 186 . doi: 10.1002/iroh.19850700202
- Taylor BE , Taylor NJ , Walsby JR 1985 . A bloom of the planktonic diatom, Ceratualina pelagica, off the coast of northeastern New Zealand in 1982–83, and its contribution to an associated mortality of fish and benthic fauna . Internationale Revue der Gesamten Hydrobiologie 70 : 773 – 795 . doi: 10.1002/iroh.19850700602
- Thompson BA 1983 . The distribution and abundance of ichthyoplankton in the Leigh Marine Reserve, New Zealand . Unpublished PhD thesis , University of Auckland , New Zealand . 82
- Tolimieri N , Haine O , Montgomery J , Jeffs A 2002 . Ambient sound as a navigational cue for larval reef fish . Journal of Bioacoustics 12 : 214 – 217 . doi: 10.1080/09524622.2002.9753700
- Tolimieri N , Jeffs A , Montgomery JC 2000 . Ambient sound as a cue for navigation by pelagic larvae of reef fishes . Marine Ecology Progress Series 207 : 219 – 224 . doi: 10.3354/meps207219
- Tricklebank KA 2000 . Effects of organochlorines on the ultrastructure of the liver of the damselfish Parma microlepis from reefs in New South Wales, Australia . Marine Biology 136 : 337 – 348 . doi: 10.1007/s002270050692
- Tricklebank KA , Jacoby CA , Montgomery JC 1992 . Composition, distribution and abundance of neustonic ichthyoplankton off northeastern New Zealand . Estuarine, Coastal and Shelf Science 34 : 263 – 275 . doi: 10.1016/S0272-7714(05)80083-6
- Victor BC 1991 . Settlement strategies and biogeography of reef fishes . In : Sale PF . The ecology of fishes on coral reefs . San Diego , CA , Academic Press . 231 – 260 .
- Westerman M , Holt GJ 1994 . RNA:DNA ratio during the critical period and early larval growth of the red drum Sciaenops ocellatus . Marine Biology 121 : 1 – 10 . doi: 10.1007/BF00349468
- Wilkens SL , Stanley JA , Jeffs AG 2012 . Induction of settlement in mussel (Perna canaliculus) larvae by vessel noise . Biofouling 28 : 65 – 72 . doi: 10.1080/08927014.2011.651717
- Williams DM , Wolanski E , Andrews JC 1984 . Transport mechanisms and the potential movement of planktonic larvae in the central region of the Great Barrier Reef . Coral Reefs 3 : 229 – 236 . doi: 10.1007/BF00288259
- Wing SR , Botsford LW , Largier JL , Morgan LE 1995a . Spatial structure of relaxation events and crab settlement in the northern California upwelling system . Marine Ecology Progress Series 128 : 199 – 211 . doi: 10.3354/meps128199
- Wing SR , Largier JL , Botsford LW , Quinn JF 1995b . Settlement and transport of benthic invertebrates in an intermittent upwelling region . Limnology and Oceanography 40 : 316 – 329 . doi: 10.4319/lo.1995.40.2.0316
- Zeldis JR 2004 . New and remineralised nutrient supply and ecosystem metabolism on the northeastern New Zealand continental shelf . Continental Shelf Research 24 : 563 – 581 . doi: 10.1016/j.csr.2003.11.008
- Zeldis JR , Davis CS , James MR , Ballara SL , Booth WE , Chang FH 1995 . Salp grazing – effects on phytoplankton abundance, vertical-distribution and taxonomic composition in a coastal habitat . Marine Ecology Progress Series 126 : 267 – 283 . doi: 10.3354/meps126267
- Zeldis JR , Francis R 1998 . A daily egg production method estimate of snapper biomass in Hauraki Gulf, New Zealand . ICES Journal of Marine Science 55 : 522 – 534 . doi: 10.1006/jmsc.1997.0277
- Zeldis JR , Jillett JB 1982 . Aggregation of pelagic Munida gregaria (Fabricius) (Decapoda, Anomura) by coastal fronts and internal waves . Journal of Plankton Research 4 : 839 – 857 . doi: 10.1093/plankt/4.4.839
- Zeldis JR , Oldman J , Ballara SL , Richards LA 2005 . Physical fluxes, pelagic ecosystem structure, and larval fish survival in Hauraki Gulf, New Zealand . Canadian Journal of Fisheries and Aquatic Sciences 62 : 593 – 610 .
- Zeldis JR , Walters RA , Greig MJN , Image K 2004 . Circulation over the northeastern New Zealand continental slope, shelf and adjacent Hauraki Gulf, during spring and summer . Continental Shelf Research 24 : 543 – 561 .