Abstract
Internal loading as phosphorus (P) released from anoxic sediment surfaces often represents the main summer P load to lakes and reservoirs and can have an immense effect on their water quality. Many difficulties in internal load assessment exist, however, including ignoring internal load altogether, ambiguity about the origin of sediment released P and inexact definitions. Most of these problems are due to the difficulty in distinguishing internal from external P sources, which is particularly challenging in polymictic lakes. To prevent misconceptions and facilitate its evaluation, internal load in stratified and polymictic lakes should be expressed in a similar way to external loads: as annual, gross and areal load of total phosphorus (TP). Possible approaches to internal load quantification are: in situ determination from hypolimnetic P increases, mass balance approaches, and estimates from anoxic active area and P release. Further suggestions to facilitate the study of internal loading include: (a) the differentiation between polymictic and stratified lakes, sections of lakes, and time periods when evaluating indicators and impact of internal load; (b) the separation of internal load (upward flux) from sedimentation (downward flux) of external and internal loads, and (c) the consideration of the downward flux of both external (Lext, mg/m2/yr) and internal (Lint, mg/m2/yr) loads by a retention model (Rsed) when predicting lake TP averages in a mass balance model of the form (qs = annual areal water load in m/yr):
After more than 70 years (CitationEinsele 1936, CitationMortimer 1941) of knowledge about phosphorus (P) release from sediments, assessment of internal P load is still one of the most challenging subjects in lake and reservoir eutrophication and restoration. We know that internal loading as P released from sediment surfaces often represents the main summer P load to lakes. Because of its high biological availability, lack of dilution and timing, it can have an immense effect on the water quality of a lake, reservoir or pond; however, the following difficulties in internal load assessment still remain:
| 1. | Undetected internal load. | ||||
| 2. | Controversy about the ultimate origin and ambiguity about the form of sediment released P. | ||||
| 3. | Unclear definitions and inconsistent units and dimensions. | ||||
| 4. | Inadequate quantification and modeling of internal load by confusing downward with upward fluxes and net with gross estimates; and | ||||
| 5. | Inadequate determination of its contribution to lake P concentration. | ||||
Most of these problems are due to the difficulty in tracking lake water P, in particular distinguishing internal from external sources. Possible approaches have been developed including regression analysis, mass balance and time-dynamic modeling (e.g., CitationNürnberg 1998, Citation2005, CitationHåkanson 2004), but many pitfalls remain and are discussed here to help prevent further misconceptions and facilitate the evaluation of internal load in all lakes and reservoirs.
In this paper I first identify and discuss problems 1–3. Next, while addressing problems 4 and 5, I present three different methods for the quantification of internal load and describe the mass balance modeling of lake P concentration as affected by the concept of P retention. Examples from case studies are presented throughout to illuminate the problems and their possible solutions.
Unnoticed internal load
Internal P load is generally attributed to reductive dissolution of P adsorbed to iron oxyhydroxides in the sediments and subsequent release from the anoxic sediment surfaces according to the classic model of CitationMortimer (1941) and CitationEinsele (1936), or to release from organic compounds (CitationGächter and Meyer 1993) and poly-phosphates in very eutrophic sediments (CitationHupfer et al. 2007) according to more recent models. Controversies exist about the origin of P derived from sediments and its release mechanism (CitationPrairie et al. 2001, CitationHupfer and Lewandowski 2008), but these questions are less important for the quantification and management of internal load. It is generally accepted that most internal loading is released in the form of ortho-phosphate, which is fully biologically available and can potentially be used by phyto- and bacterio-plankton (CitationCooke et al. 2005).
While external P sources are generally recognized as contributors to lake P concentration, internal sources are often overlooked. In stratified lakes, the epilimnion can seem to be untouched by internal load during the summer, and a monitoring program could easily miss its occurrence if sampling occurs only in the mixed epilimnion. Nevertheless, certain seasonal and spatial patterns indicate the occurrence of internal load (). Typically, total P (TP) and dissolved reactive P (DRP, analysis for ortho-phosphate, often also called soluble reactive P or SRP) concentrations increase in the hypolimnion during summer stratification so that profiles show increasing concentrations below the thermocline toward the sediment (). If such elevated P concentrations are associated with anoxia and the prevalence of some reduced substances including ferrous iron, manganeous manganese, ammonium and gases of hydrogen sulfide and methane, the occurrence of internal P load is certain (). In addition to hypolimnetic P increases, epilimnetic increases due to entrainment and diffusion during the period of thermocline erosion later in the summer have been documented (CitationMataraza and Cooke 1997) and were included in a seasonal mass balance model (CitationAuer et al. 1997). Further, conspicuous increases during and after fall turnover are a definite sign of internal P load when it becomes mixed into the surface water (CitationNürnberg and Peters 1984b, CitationNürnberg 1985).
Table 1 Indicators of internal load in stratified (A) and polymictic lakes (B)
Figure 1 (A) P and DRP profiles at the dam of eutrophic Brownlee Reservoir, Snake River, CO, 11 August 1999; (B) P, dissolved oxygen (DO) and iron profiles (TFE = total; SFe = soluble; FE2 = ferrous) in oligotrophic Chub Lake, Ontario, Sep 13, 1982 (note lake characteristics in ).
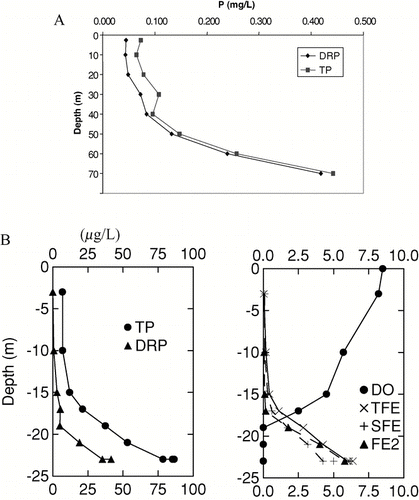
In shallow, polymictic water bodies it is more difficult to distinguish internal from external loads because their water is usually vertically mixed, in addition to the horizontal exchange as happens in all lakes (CitationSøndergaard et al. 2005); therefore, internal load indicators are different in polymictic lakes (). For example, some P is released from bottom sediments into the mixed overlaying water so that it is taken up by phytoplankton and may foster algal and cyanobacterial blooms in shallow lakes, while a large proportion remains as DRP in the stagnant summer hypolimnion of stratified lakes (). Consequently, internal load affects surface water quality in shallow polymictic lakes even in summer, but mostly during thermocline erosion and turnover in the fall in stratified lakes.
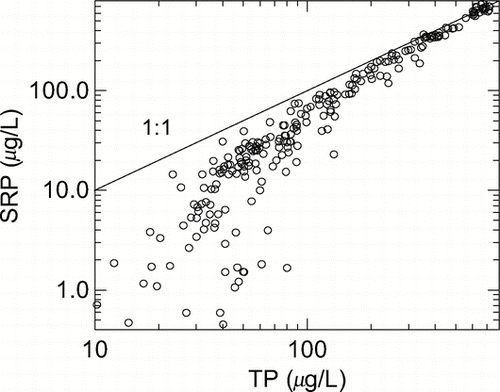
Figure 2 Comparison of DRP with TP from individual measurements in the anoxic hypolimnion of eight stratified Canadian lakes (with data from CitationNürnberg and Peters 1984b).
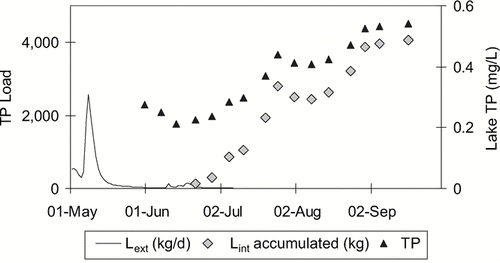
The effect of internal load is obvious when TP increases during a summer drought where all external inputs cease. In the shallow, polymictic reservoir Lake Mitchell, South Dakota (), TP concentrations typically increase throughout the summer. Because in summer 2001 the inflow, which on average contributes 92% of the annual external TP load, had ceased, all increases had to derive from internal sources and can be used to quantify internal load (after correction for changes in lake level; ). Similarly, evidence of internal load was observed when TP concentrations greatly exceeded inflow concentrations in western Washington lakes (CitationWelch and Jacoby 2001).
Table 2 Characteristics of lakes and reservoirs used as examples and case studies (Nürnberg, unpublished data; avg = average, max = maximum).
Figure 3 Lint_1 estimate from in situ TP increases in polymictic Lake Mitchell Reservoir, SD, for summer 2001.
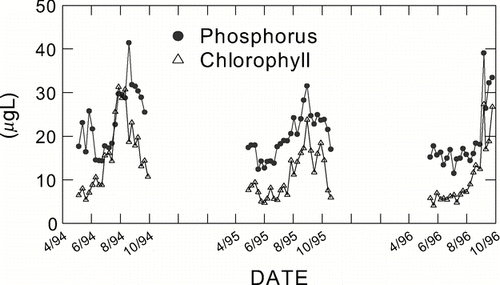
Differences due to the mixing state are less clear in lakes that are polymictic in some summers but stratified in others, as in Brome Lake, Quebec (). In this lake, TP increases were followed by chlorophyll increases throughout two growing seasons, while in one year (1996) increases happened only in late September (). Thus, the mixing state in polymictic lakes can be variable between years, sometimes more resembling the summer stratification of dimictic lakes, like in 1996 in Brome Lake.
Figure 4 TP and chlorophyll concentration for three growing seasons in polymictic Brome Lake, Quebec.
Differences are also less pronounced in water bodies with both stratified and polymictic areas. This is particularly important for deep, run-of-the river reservoirs that deepen gradually from the shallow and polymictic section at the inflow to a deep stratified section at the dam. Such differences also exist in lakes where a large shallow area and a deeper area exist in the same lake, as in Lake Pyhäjärvi, Finland () and Mona Lake, Michigan (CitationSteinman and Ogdahl 2008). In these lakes a high proportion of DRP in the stratified part indicates that internal loading is the P source, while a low proportion in the shallow riverine section may be the consequence of immediate transformation of orthophosphate into algae biomass and adsorption onto silt particles after its release from the sediments, in addition to sediment resuspension.
These examples reveal many clues to internal load in lakes and reservoirs despite its seeming invisibility. Such signs are especially obvious when the stratification regime is considered so that polymixis and stratification are differentiated with respect to the whole or partial sections of the lake or reservoir and different time periods.
Phosphorus release is not restricted to and does not require anoxia in the overlaying water. It suffices to have a mechanism that conveys the P released into the porewater from sediment iron-oxyhydroxides to the sediment surface. In addition to low redox potential at the sediment surfaces, P release can be enhanced by bioturbation, when Chironomids effectively pump the phosphate-rich porewater into the overlying water (CitationHoldren and Armstrong 1980). Persistent P release occurs especially from highly eutrophic sulfuric sediments where the formation of iron sulfides effectively removes iron from the P-Fe cycle and liberates P from vivianite. A result of such mechanisms is that artificial hypolimnetic oxygenation by pure oxygen in eutrophic Swiss lake Sempachersee did not decrease internal P loading during more than 15 years of this elaborate and expensive restoration treatment (CitationGächter and Müller 2003).
Chemical forms of sediment released P
Although internal load is typically ortho-phosphate release, P speciation, such as the analytical analysis of ortho-phosphate as DRP, does not necessarily lead to a reliable quantification of internal load. Only in anoxic hypolimnia with elevated TP concentrations can correctly conducted DRP determination be used to quantify internal load, because under these conditions most of the sediment-released P remains DRP, as discussed earlier ( and ). In anoxic P-rich hypolimnia, a large proportion of DRP was biologically available when mixed with epilimnetic plankton in short-term bioassays, so that DRP also quantifies the potential bioavailability of internal load in anoxic hypolimnia (CitationNürnberg 1984a; CitationNürnberg and Peters 1984a).
However, analytical errors are abundant when determining DRP in anoxic waters (CitationNürnberg 1984a), especially underestimation, for several reasons: (a) accidental aeration during the sampling and filtration procedure can lead to phosphate adsorption onto ferric iron; (b) conversely, neglecting to degas the sample by aeration before analysis implies interference by the hydrogen sulfide gas with the molybdenum blue analysis in more reduced hypolimnetic water; (c) delay in analysis immediately after sample collection allows P uptake by phytoplankton and bacteria. In addition to possible underestimation, overestimation can easily occur due to contamination when dealing with low concentrations of DRP.
More important, P compounds are altered after release in most circumstances except in anoxic hypolimnia. Generally, changes of the released P depend on conditions such as trophic state, chemistry and mixing regime of the lake water and abundance and nutrient state of the plankton (CitationSoslash;ndergaard et al. 2001). The following pathways have been described for sediment-released P (CitationNürnberg 1984a, CitationNürnberg 1985): (1) it remains ortho-phosphate (and is analyzed as DRP); (2) upon aeration it may become adsorbed onto ferric iron hydroxide particles that are larger than 0.45 μ m and are not analyzed as DRP; (3) it is analyzed as DRP in the presence of organic acids, in particular humic and fulvic acids, which can keep adsorbed P small (< 0.45 μ m) and suspended (in this case, DRP would overestimate phosphate); (4) it becomes incorporated into algae biomass that outcompetes aeration processes described in cases 2 and 3 due to high dilution during entrainment of hypolimnetic water and mixing events.
Because all these processes may have taken place after the release of sediment-derived ortho-phosphate, determination of DRP mass alone will not adequately quantify internal load in many circumstances. Furthermore, because analytical determination of DRP is problematic in anoxic waters, I suggest quantifying internal load as mass of TP (molybdenum blue analysis after complete digestion; e.g., CitationMurphy and Riley 1962) for routine determination. This method analyzes all P compounds in a water sample as TP, is easy to carry out and relatively free of interference, if contamination is avoided.
Inexact definitions
Internal load estimates are sometimes ambiguous because studies relate internal load to various time periods, including year, growing season, summer, month, anoxic period or stratified period. Some studies contemplate the question about the original source of internal load, which in eutrophic lakes most often is external. Considering the long-term, such load is not actually internal (CitationHupfer and Lewandowski 2008; Michael Coveney, St Johns River Water Management District, Department of Water Resources, Palatka, FL, U.S.A., 2006, pers. comm.). Other studies relate internal load to varying areas, such as total lake surface and anoxic or hypolimnetic area. For example, the Lake Mendota research group refers only to the portion of the sediment-released P that is entrained into the epilimnion throughout the growing season and has an immediate effect on phytoplankton (e.g., CitationLathrop et al. 1999).
However, calculations that do not consider gross release from the sediments during the whole year under investigation, underestimate internal load and its contribution to lake P concentration; the origin (internal vs. past external) is irrelevant for quantification unless the load originates from external sources of the current study year. In some highly eutrophic systems a large portion of the present external load possibly settles and becomes recycled within the same year, resembling sediment-released P (CitationCoveney et al. 2005; CitationHupfer and Lewandowski 2005). In such cases the differentiation of internal from external sources is ambiguous, and the direct impact of external on the apparent internal load is obvious. If significant, the portion of the present internal load that stems from such a current external source is best distinguished from a former external source (e.g., 30% of the sediment released TP is recycled from this year's external load, the source of the present internal load stems mostly from the years before sewage diversion or it was created during 10 years of aquaculture).
In general, the expression of gross internal load as P mass released per lake surface area and year (Lint, mg/m2/yr) is comparable and analogous to the definition of external P load (Lext, mg/m2/yr). In comparison, and to prevent ambiguity, an anoxic P release rate (RR, mg/m2/d) can be defined as a rate of P release from actively releasing sediments, with units of P mass released per anoxic sediment surface area and day.
Confusing net with gross estimates of internal load
The terms gross and net with respect to internal load refer to whether the proportion that settles annually to the lake bottom has been considered in the calculations. For example, external loading is usually expressed as gross because it is calculated from individual external P sources such as precipitation, runoff and point sources before it enters the lake; it has not been subjected to changes within the lake such as biological and physical uptake and sedimentation. In contrast, internal loading originates from the lake bottom in any given year, and its measurement may include settling events of the summer season that have taken place before its final quantification.
Considering theses processes, CitationNürnberg and LaZerte (2001) and Nürnberg et al. (in prep.) proposed three principally different procedures to determine internal load depending on available information:
| 1. | A partially net estimate from in situ summer increases | ||||
| 2. | Net (and gross) estimates from complete P budgets (mass balance approach) | ||||
| 3. | A gross estimate from P release (anoxic areal P release rate, RR in mg/m2/d) and measured anoxia (as the anoxic factor, AFmeas, in days per summer) in stratified lakes and predicted anoxia (AFpred) in polymictic lakes (CitationNürnberg 2005; AF × RR approach) | ||||
While Methods 1 and 2 are independent of theoretical considerations with respect to the origin of internal load, Method 3 assumes that the release depends on and is enhanced when sediment surfaces become anoxic. This is expected during microbial mediated anoxia for the classic release from ferrous oxy-hydroxides as well as from polyphosphates of organic compounds and does not require anoxia in the overlaying water (CitationHoldren and Armstrong 1980). Approaches, underlying assumptions, models and formulas in these methods are sometimes different for stratified and shallow lakes or water bodies with unusual features such as hard water, bottom water outlet, and for fast flushed reservoirs.
Method 1: In situ P increases (Lint_1)
When using summer in situ TP increase to estimate internal load according to equation Equation1, some sedimentation has already happened before quantification, yielding a partially net estimate.
In strongly stratified lakes, Lint_1 is the difference between the end-of-summer hypolimnetic or whole water column TP mass (P_t2 × V_t2) and the respective TP mass at the beginning of the anoxic period (P_t1 × V_t1), divided by lake area. Any winter internal load under ice could be determined in a similar fashion.
In polymictic and weakly stratified lakes with early thermocline erosion, or where anoxia approaches the thermocline, the maximum summer whole water column mass of the anoxic period may be more appropriate to use as final mass (P_t2 × V_t2), rather than a mass from the end of summer, to account for entrainment of sediment released P into the surface water and subsequent loss though the outflow. Such calculations are presented for polymictic Lake Mitchell, where the inflows were low or not existent in the summer (; ).
Table 3 Internal load estimates (Lint_1, equation Equation1) for 11 summers (summer, su = May to “End-Date”) from in situ TP concentration changes in polymictic reservoir Lake Mitchell, SD.
When external water and TP inputs and exports are variable and high during the period of computation, the unsettled portion of external load should be included:
In a related approach (CitationMoore and Christensen 2009), partial P budgets throughout the growing season were calculated for each sampling event, and those indicating an internal source were summed. In this way, several internal loading occasions were documented and quantified and provided some evidence of the extent of internal loading. Such an estimate is also partially net because it does not take any previous settling into account.
As in any lake characteristic calculated from seasonally and spatially variable data, the involved monitoring frequency, quality and quantity all contribute to the accuracy of the in situ internal load assessment. This contributes to the commonly high variability observed in Lint_1.
Method 2: Annual TP mass balance (Lint_2)
A net estimate of internal load can be obtained from an annual TP mass balance in stratified and polymictic lakes (CitationNürnberg 1998, CitationNürnberg and LaZerte 2001). Because it is based on an annual mass balance, Lint_2 includes summer and winter loads. This process is data intensive because it requires annual input and export data for both TP and water. When winter data are not available, Lint_2 can be approximately estimated from summer mass balances alone (CitationWelch and Jacoby 2001).
The main complication is that upward (internal load) and downward (settling) fluxes are combined in a lake budget. This means that the proportion of P that is retained and can be determined from mass balances as measured retention (Rmeas, equation Equation3) yields net estimates of retention:
Accordingly, only if sedimentation is known can internal load be determined from mass balance studies because sedimentation occurs in all but fast flushing reservoirs and is often quite high. For example, in approximately 100 of 305 lakes at least 60% of the incoming P mass was retained (CitationBrett and Benjamin 2008). Even when P export exceeds input by a certain amount on an annual basis (in lakes with high internal loading after sewage diversion; CitationSas 1989) it would be wrong to suppose that only this marginal amount is due to sediment release. Such an assumption would underestimate internal load because a certain fraction (the exact amount can be modeled by Rsed depending on hydrology and morphometry) of both external and internal load has settled over the year.
Average P settling or sedimentation rates are extremely hard to determine in lakes and therefore are almost always modeled instead. Consequently, Method 2 largely depends on a retention model that determines the part that settles (Rsed) so that downward fluxes can be separated from upward fluxes of TP. Such models are described in detail in Problematic retention.
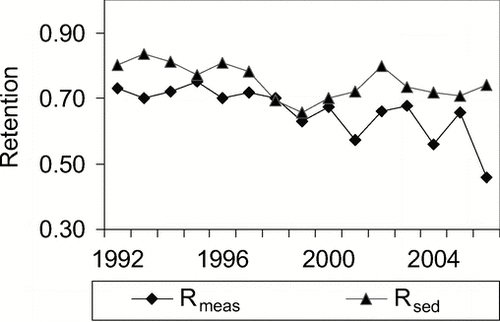
Because it only represents the (downward) sedimentation, predicted retention is larger than measured retention in lakes with internal loads (), approximately by the amount of P released from the sediments, so that the difference of the retentions quantifies the net upward flux out of the sediment or net Lint_2 (CitationNürnberg 1984b):
Figure 5 Comparison of measured net P retention (Rmeas from mass balance) and predicted gross retention (Rsed) for Cherry Creek Reservoir, Denver, CO (). Differences are due to internal load.
The conversion of net to gross estimates is possible when the proportion of internal load that has settled within the year is known. The annual retained (settled) proportion of internal load is not dissimilar to that of external load according to CitationNürnberg (1998), so that gross estimates of Lint_2 can be computed:
Note that values of net and gross loads are extremely different (), and mixing net, partially net and gross loads severely compromises the quantification of internal load.
Table 4 Comparison of long-term median net and gross internal load estimates (equation Equation5) according to Method 2 in several water bodies. Years indicates the sample size as the number of years with mass balance estimates.
Method 3: Product of active (anoxic) sediment area and P release rate (L int_3)
A more direct method of determining gross internal load is based on its components, the P releasing area and rate. Anoxic release rates (RR) have been shown to yield gross internal load when multiplied by an expression for the releasing sediment area in stratified (CitationNürnberg 1987) and polymictic lakes (CitationNürnberg 2005). In this method, gross internal load is determined separately for summer and winter and then summed for an annual estimate:
In stratified lakes, summer internal load is determined as the product of release rate and anoxic factor according to (CitationNürnberg 1987):
In polymictic lakes summer internal load (equation Equation8) is determined as the product of release rate and the representation of the actively releasing area modeled as anoxic factor, AFpred (equation Equation9, Nürnberg 1996):
A base (or average) areal release rate for stratified or polymictic lakes can be determined in the lab, or predicted from sediment TP concentration, reductant-soluble P fractions, or lake trophic state (CitationNürnberg 1988) to yield a long-term average internal load.
P release rates and releasing area are influenced by temperature (CitationJensen and Andersen 1992), and variability in internal load due to varying temperature between years is obvious in polymictic lakes (Nürnberg, unpubl. data). To include this variability, both AFpred and RR can be adjusted with respect to the summer mixed layer water temperature to follow the Q10 rule of Van Holst:
Winter internal load is typically small because the low temperature decreases oxygen consumption and P release. If necessary, it can be determined in a manner similar to summer internal load from the product of the winter anoxic factor and a winter release rate (equation Equation11), although the RR and AF models do not apply to winter temperatures. The AFwinter can be computed from oxygen profiles under ice, and winter release rates (RRwinter) can be either measured in the lab at in situ temperature or predicted from the summer release rate average according to the Q10 rule.
Different from the other two methods, submodels are used that were originally developed for P release from anoxic sediments. Therefore, if other internal sources exist, for example those of senescing macrophytes (CitationGraneli and Solander 1988), littoral P release (CitationCyr et al. 2009), benthivorous fish (CitationSereda et al. 2008), and groundwater, these sources may have to be quantified independently. However, they are usually small and negligible compared to the sediment derived portion (CitationSereda et al. 2008).
Comparison
Quite often only one of the three methods presented here can be applied, and sometimes only for an overall long-term average because detailed data are seldom available. In such cases the large uncertainty of any internal load estimate has to be considered. In addition, long-term annual estimates produced by the three methods can be expected to differ from each other because of the various uncertainties discussed previously. However, if the overall long-term averages are similar, while considering differences in net versus gross estimates, it lends credibility to the estimates derived from the different methods.
In particular, Lint_1 may be less exact in polymictic lakes and can always be expected to be lower than the other two methods because it is partially net. The method of choice may be Lint_2 in cases where in- and outflows have been monitored intensely, as in important reservoirs. Lint_3 does not capture much annual variability when its components are largely predicted from average lake TP concentrations, and the other methods are more useful if estimates of long-term trends and annual variability are required.
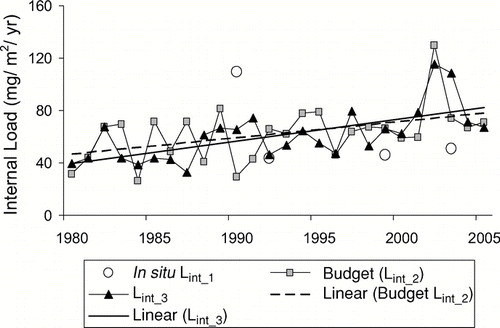
All three methods were used in the internal load assessment of polymictic Cherry Creek Reservoir near Denver, Colorado, where hard water and a bottom outlet influenced predictions of sedimentation (, , and discussed in Modeling lake TP concentration) and of polymictic Lake Pyhäjärvi, where the TP cycle was complicated by biomanipulation efforts ( and Nürnberg et al. in prep.). While differences between estimates are due to errors in the input data as well as model structure, differences with respect to Lint_1 may be due to gross versus net estimates to a varying degree. It is therefore important to determine whether a load estimate represents a gross, net, or partially net estimate; only then can it be appropriately used for TP predictions.
Figure 6 Comparison of different internal load estimates for polymictic Cherry Creek Reservoir, Denver, CO. Lint_3 estimates are based on a constant RR and moderately variable active areas.
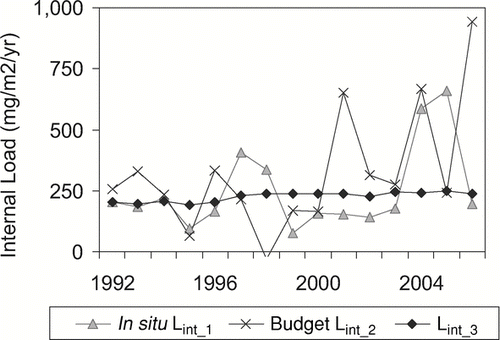
Figure 7 Comparison of different internal load estimates in partially polymictic Lake Pyhäjärvi, Finland. Lint_3 estimates include annual changes in summer temperature. Note that there was a variable biomanipulation effort between years with fish catch exporting as much TP as the outflow. An increasing trend with time is detectable for both estimates (R2 = 0.21 for Lint_2; R2 = 0.43 for Lint_3).
Modeling lake TP concentration in lakes with internal load
The main reason for quantifying internal load is to determine its effect on the trophic state and water quality of a lake. In such an attempt, internal load is best compared to external load and explicitly included in a model that predicts lake TP concentration.
Empirical P mass balance modeling approaches are the most often used and are readily available. In a generic mass balance (equation Equation12, areal external P load divided by areal waterload of the outflow, corrected for retention, Rmeas as in equation Equation3; CitationDillon and Rigler 1974), annual average outflow TP concentrations, which are deemed to be similar to annual surface concentrations, are computed from the measured inputs and outputs:
However, models of this structure do not always predict TP concentrations in lakes with internal load accurately because of un-met assumptions and the need of modeling retention.
Problems using outflow
Being based on P export, the mass balance is solved for annual average outflow P concentration, which is assumed to be similar to annual average lake P concentration; however, lake P concentrations during the growing season are the most important and determine the trophic state. In a lake with seasonal P inputs like internal load, summer P concentrations would be underestimated because they are usually higher than annual average (surface) outflow concentration, especially in polymictic lakes. Also, the mass balance does not apply to lakes and reservoirs with a mid-depth or bottom outlet. In these cases P concentration of the surface mixed layer would be overestimated because much of the P released from sediments at greater depths is lost through the outflow.
Problematic retention
Even if the assumptions of the mass balance in equation (Equation12) are fulfilled, this method is rarely applicable because detailed P budgets including retention are not often available. Consequently, retention models (R-models) have been developed, and their performance has been discussed since their explicit initial definition (CitationDillon and Rigler 1974) because of their uncertainty in lake and reservoir P modeling (CitationSøndergaard et al. 2001, CitationHejzlar et al. 2006, CitationBrett and Benjamin 2008).
Retention measured from mass balances as Rmeas (equation Equation3) is a net estimate and includes upward and downward fluxes. Accordingly, R-models developed on mass balances from these lakes also represent net estimates. When models were developed with data that included lakes with any internal loading, the upward flux partially canceled out the downward flux so that flux issues were confused, and the separation between sedimentation and internal load was prevented. Regrettably, many R-models include internal load to an unspecific degree, depending on the original lakes that were used in model development.
As a result, TP concentration tends to be underestimated if such an R-model is applied to a lake with internal load; however, TP is overestimated if the model is applied to a lake without internal load. These R-models have been widely used for a number of years (see review by CitationBrett and Benjamin 2008), despite the known problem (CitationNürnberg 1984b). To avoid the most severe values of internal load, sometimes water bodies with negative R (more export than input) were excluded when developing a model (CitationHejzlar et al. 2006) or only oligotrophic and mesotrophic lakes were used (CitationLarsen and Mercier 1976). But lakes with less apparent internal loading were still included.
Low predictability of previous R-models has spawned a number of attempts to improve them by expanding the dataset, often combining the lakes of former studies on R-models (CitationBrett and Benjamin 2008), by using different and more elaborate statistical approaches (CitationPrairie 1989), by investigating the importance of specific variables in a hierarchical analysis (CitationCheng et al. 2009), by distinguishing between reservoirs and lakes (CitationStraskraba 1998, CitationHejzlar et al. 2006) and by including additional variables that address the outflow problem (e.g., ratio of outflow to annual average lake P concentration; CitationSchauser and Chorus 2007).
One general approach with some mechanistic appeal is based on the annual average settling velocity, v (m/yr), that can be constant or variable, and annual water load, qs (equation Equation13). I describe it here in more detail because it makes theoretical sense to use an expression of v for the prediction of sedimentation (without upward fluxes) in an R-model:
Constant values between 10 and 30 m/yr for v were found empirically to fit annual mass balances in lakes and reservoirs that included some lakes with internal loads and hypolimnetic anoxia (reviewed in CitationNürnberg 1984b). In a study where more than half the lakes were eutrophic (and likely releasing P from their sediments), optimization of v yielded a small value of 5.1 m/yr, as can be expected because downward fluxes are partially canceled by upward fluxes in such lakes (CitationBrett and Benjamin 2008).
A specifically designed R-model that predicts only sedimentation (and not release) was developed from a more general form of equation Equation13, using two different constants. The constants were calibrated with a dataset of lakes with no or low potential of sediment P release because there was no evidence of hypolimnetic anoxia (CitationNürnberg 1984b). In such lakes, Rmeas (equation Equation3) essentially describes the proportion that settles because it can be assumed that internal load does not occur or is only marginal. The specific R-model (Rpred, equation Equation14) developed for these lakes therefore represents the downward flux of TP due to settling and sedimentation:
While the parameters in Rpred (equation Equation14) seem to predict sedimentation well in many lakes (CitationNürnberg 1984b), it underestimates P sedimentation in certain lake types. For example, settling is enhanced in hardwater lakes experiencing P-calcite co-precipitation. Retention averaged 39% higher than predicted by Rpred in stratified, calcium-rich alpine lakes with 11–50 mg/L Ca (CitationNürnberg 1998). In some of these cases, calibration to find a more appropriate value of v can produce an adequate R-model. For example, average annual settling velocity was 25.5 m/yr in a study on 28 stormwater detention ponds and urban lakes designed to settle solids; it was also higher in run-of-the-river reservoirs (Nürnberg, unpublished studies).
In some cases where R-models based on a constant v have not been successful, those based on a variable v were more useful. For example, models especially designed for shallow lakes and reservoirs relate v to annual water detention time, τ (“tau,” also called water residence time, with units in year, measured as lake volume divided by annual outflow volume) and qs, along with a constant k:
Inserting this v relationship (equation Equation15) into equation Equation14 with simplification (equation Equation16) leads to the following retention model, Rτ (equation Equation17):
The original relationship was developed for natural lakes with k = 1 (CitationLarsen and Mercier 1976). An Organisation for Economic Co-operation and Development (OECD) study of 43 European, Australian and Japanese man-made reservoirs and some shallow lakes determined k = 2 (CitationClasen 1980). A study with 119 records of European and North American reservoirs that ranged from deep to shallow and oligotrophic to eutrophic determined k = 1.84 (CitationHejzlar et al. 2006). This study also found, like previous studies (CitationStraskraba 1998, Nürnberg, unpubl. data), that retention of reservoirs is far higher than retention in natural lakes.
As in most other discussed models, there was no provision in these models to accommodate internal load separately from sedimentation, although, for example, more than 70% of the lakes and reservoirs in the OECD dataset were eutrophic with lake TP concentrations between 30 and 100 μ g/L. Furthermore, sediment P release was deemed to occur in the more eutrophic OECD systems (CitationClasen 1980) as well as in the CitationHejzlar et al. (2006) study, although they excluded reservoirs with obvious and large amounts of internal load (determined from negative net retention in the mass balance). Consequently, it can again be argued that retention, and in particular k, is underestimated in most of these models because of the omission of sediment-released P, and that their computed retention is actually a net estimate that includes upward fluxes.
The R-models discussed here were developed on long-term averages for several years so that they do not necessarily apply to individual lakes and even less to specific years. By using models that include a term that relies on lake-specific hydrology, like qs (equation Equation14) or τ (equation Equation17), annual variances in hydrology and climate can be captured and modeled even for a given year of a specific lake.
Therefore, a lake-specific calibration of Rτ (equation Equation17), for example, can create a valid R-model as long as internal load is considered. Eutrophic, hardwater Cherry Creek Reservoir near Denver, Colorado (), has a bottom outlet, and therefore average summer TP (Jul–Sep) could not be predicted from available R-models because the simple Rpred model (equation Equation14) did not apply. Using 15 years of TP budgets, the most appropriate R-model was determined to be of the Rτ form (equation Equation17) with a k value of 2.7. Calibration was done simultaneously with independent estimates of internal load (Lint_1; ) and seasonal TP values (see next section).
For the reasons discussed here, any R-model must be tested and, if necessary, calibrated to accurately predict TP concentrations in a specific lake or reservoir. I recommend testing the simple Rpred model first and if results are considered inaccurate, to calibrate v in the model of the type Rv (equation Equation13) or Rτ (equation Equation17). In the end, and despite theoretical considerations, the model that fits the data is the best or most appropriate model in any particular case (CitationChapra 1997) and may have to be determined by trial and error (testing). Consequently, model testing (and hopefully verification) with several seasons of lake TP concentration data is essential.
Application of R sed in P model
The prediction of lake TP concentration, the ultimate goal for the development of R-models, can be accomplished by adding an internal loading term and replacing Rmeas with Rsed in equation Equation12, where Rsed is any R-model specific to downward fluxes, such as Rpred (equation Equation14). In this way, both upward and downward fluxes are considered, and specific seasonal or annual TP concentrations can be predicted in stratified and polymictic lakes according to Nürnberg (Citation1998, Citation2005).
The model for average TP concentration is based on the same term of retention for both external and internal loads and TP is predicted using:
This model predicts annual average TP in stratified lakes with and without internal loading () and growing season concentrations of polymictic lakes (CitationNürnberg 1998). This formula also seems to be the most useful for the calibration of a lake specific R-model as described in the previous section ().
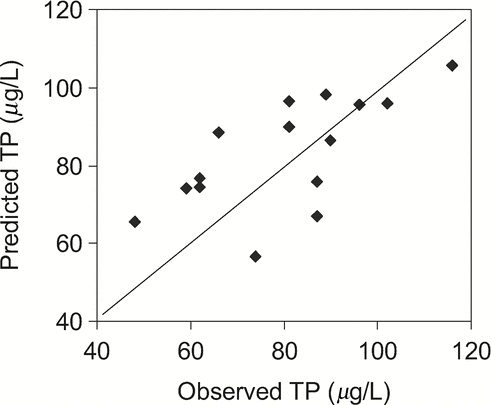
Figure 8 Comparison of observed summer TP averages with those predicted from the TP model (equation Equation18 with Rsed = Rpred of equation Equation14 and Lint) for 33 non-alpine lakes with data from Nürnberg (Citation1998; R2 = 0.84, p < 0.0001, slope is not significantly different from one and t-test on differences is not significant, p = 0.383). The line of perfect prediction (solid) and the 95% confidence band around the regression line (broken) are shown.
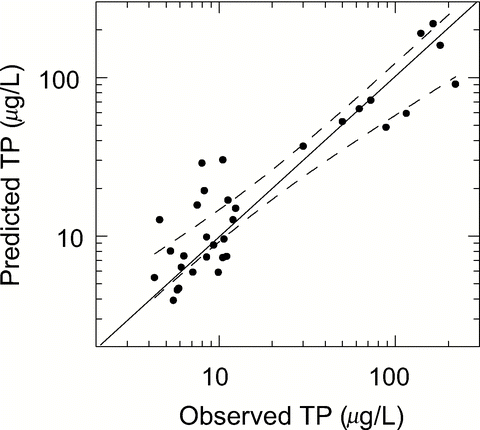
Figure 9 Comparison of observed Jul-Sep TP averages with those predicted from the TP model (equation Equation18 with Rsed = Rτ of equation Equation17 and k = 2.7 and Lint_1) for 15 years of polymictic Cherry Creek Reservoir, Denver, CO (p < 0.01, R2 = 0.38, n = 15). The line of perfect prediction is shown.
Because of the seasonality of sediment release, early summer TP minimum can be predicted without considering internal load (equation Equation19) and a late-fall maximum that does consider the entire gross internal load (equation Equation20). However, these two models usually deliver extreme values (too small or too large) and are mainly used to indicate ranges (CitationNürnberg 1998):
As with the R models, TP models that do not consider internal load are not applicable to eutrophic lakes (with internal load). For example, when the original and most influential TP model is used (that does not consider internal load specifically; CitationVollenweider 1976), TP concentrations are underestimated in lakes where internal load is large (; CitationWelch and Jacoby 2001). This is a well-known and acknowledged problem (Vollenweider, Canadian Centre of Inland Waters, pers. comm., 1982; CitationLarsen and Mercier 1976, CitationHejzlar et al. 2006, CitationBrett and Benjamin 2008), discrediting all P mass balance modeling in lakes and reservoirs. Of course the main reason for using models that include downward and upward fluxes in one expression is the difficulty of quantifying internal load. If the different methods (Lint_1, Lint_2, Lint_3) provided here are applied, only the downward flux has to be predicted by R-models.
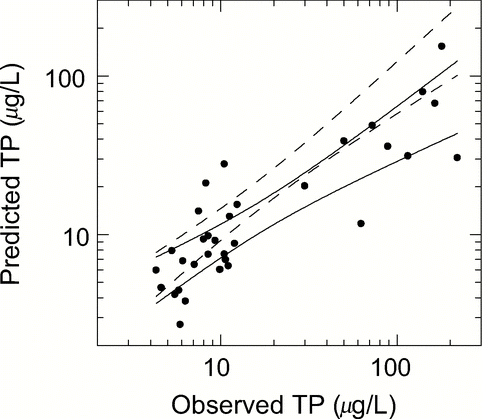
Figure 10 Comparison of observed summer TP averages with those predicted from the Vollenweider model (with Rv of equation Equation13 for v = 10, and no provision for internal load) for 44 stratified, non-alpine lakes (including 33 of , computed with data from CitationNürnberg, 1998. R2 = 0.72, p = 0.0001, n = 44, slope = 0.697, SE = 0.068 significantly different from one, and t-test on differences significant, p < 0.05). The 95% confidence bands are shown (solid). Bands of the TP model of , are shown for comparison (broken).
Recommendations (summary and conclusion)
Remediation and restoration of lakes with unsatisfactory water quality is challenging. The first and only pretreatment investigation in eutrophic lakes usually includes the determination of the different P sources. While external sources are generally more obvious, internal sources are harder to quantify and are often ignored. In particular, internal P loading as sediment-released P can be large but undetected. Internal load seems to be more effective and harmful in shallow polymictic lakes than in deep stratified lakes, even if more difficult to identify. Destratification and aeration treatments that effectively turn stratified lakes into polymictic ones and mix sediment-released P into the trophic zone often increase TP concentration and algae biomass, even favoring cyanobacteria (CitationNürnberg et al. 2003; reviewed in CitationCooke et al. 2005). But internal load also negatively affects the trophic state in stratified lakes. When internal load was moderated with the restoration treatment of hypolimnetic withdrawal, water quality improved as long as stratification was maintained. As expected, however, this treatment was not beneficial in polymictic lakes where sediment-released P was mixed throughout the water column (CitationNürnberg 2007).
The following suggestions may simplify studies on lakes with internal loading:
| • | Distinguish between stratified (i.e., mono- and dimictic lakes) and shallow polymictic lakes. | ||||
| • | Use TP for the quantification of internal load to avoid analytical problems inherent to the analysis of DRP and other P fractions. | ||||
| • | Reserve the term “internal load” for the gross P mass released from sediment surfaces and express it per lake surface area and per year (Lint, mg/m2/yr) to render it comparable and analogous to the definition of external P load (Lext, mg/m2/yr). It can be quantified from mass balance after correcting for its sedimentation (conversion of net into gross) or from the active area and release rate (AF × RR). Determination from in situ summer increases delivers a partially net estimate. Internal load should correspond to the specific study year or average of a period of years; this definition does not consider the previous contribution of external load to the current internal load. | ||||
| • | Use internal load estimates in TP mass balance models, so that downward fluxes and separated from upward fluxes and only the downward flux has to be predicted. | ||||
| • | Predict downward fluxes with a retention model (Rsed), test and calibrate Rsed, if necessary, with independent internal load estimates and seasonal lake TP concentration. | ||||
Glossary – definition of terms (note that all terms refer to lakes and reservoirs)
V (106 m3) : lake volume | |||||
Ao (106 m2) : lake surface area | |||||
qs (m/yr) : annual areal water load, annual outflow volume (Q, m3) per surface area (Ao, m2), where qs = Q/Ao. | |||||
τ (yr) : annual water detention time or annual water residence time, lake volume (V) divided by annual outflow volume (Q), where τ = V/Q. | |||||
v (m/yr) : settling velocity, average distance that TP settles downward within one year. | |||||
Lext (mg/m2/yr) : external (gross) load. | |||||
Lint (mg/m2/yr) : internal (gross) load. | |||||
Rmeas : measured or observed retention, proportion of external load (measured as in–out/in, equation Equation3) | |||||
Rsed : predicted retention, proportion of settling loads, generic expression that is predicted by models for specific lakes (e.g., equation Equation14: Rpred = 15/(18 +qs). | |||||
Acknowledgment
This manuscript is based on workshops and various presentations (SIL and NALMS 2007, 2009) and the discussion with participants and audience is greatly appreciated. The critical review and helpful comments from my colleagues, Chris Holdren, Bruce LaZerte, Barry Moore, Eugene Welch, and one anonymous reviewer greatly improved the manuscript. Data provision from various contract work is acknowledged for the case studies listed in .
Notes
*s = stratified; m = mixed; s/m = depending on location or year: s or m.
**Based on classification of CitationNürnberg (1996). o = oligotrophic; m = mesotrophic; e = eutrophic; h = hyper-eutrophic.
*Based on modeled external load.
References
- Auer , M. T. , Doeer , S. M. , Effler , S. W. and Owens , E. M. 1997 . A zero degree of freedom total phosphorus model; 1. Development for Onondaga Lake, New York . Lake Reserv. Manage. , 13 : 118 – 130 .
- Brett , M. T. and Benjamin , M. M. 2008 . A review and reassessment of lake phosphorus retention and the nutrient loading concept . Freshwater Biol. , 53 : 194 – 211 .
- Chapra , S. C. 1997 . Surface water-quality modeling , 844 Boston : McGraw-Hill Companies, Inc. .
- Cheng , V. , Arhonditsis , G. and Brett , M. 2009 . A revaluation of lake-phosphorus loading models using a Bayesian hierarchical framework . Ecol Res. , 0912-3814 (Print) 1440-1703 (Online)
- Clasen , J. 1980 . Shallow lakes and reservoirs , 246 Oganisation for Economic Co-operation and Development. OECD .
- Cooke , G. D. , Welch , E. B. , Peterson , S. A. and Nichols , S. A. 2005 . Restoration and management of lakes and reservoirs , 576 Boca Raton, FL , , USA : CRC .
- Coveney , M. F. , Lowe , E. F. , Battoe , L. E. , Marzolf , E. R. and Conrow , R. 2005 . Response of a eutrophic, shallow subtropical lake to reduced nutrient loading . Freshwater Biol. , 50 : 1718 – 1730 .
- Cyr , H. , McCabe , S. K. and Nürnberg , G. K. 2009 . Phosphorus sorption experiments and the potential for internal phosphorus loading in littoral areas of a stratified lake . Water Res. , 43 : 1654 – 1666 .
- Dillon , P. J. and Rigler , F. H. 1974 . A test of a simple nutrient budget model predicting the phosphorus concentration in lake water . J. Fish. Res. Board Can. , 31 : 1771 – 1778 .
- Einsele , W. 1936 . Über die Beziehungen des Eisenkreislaufs zum Phosphatkreislauf im eutrophen See . Arch. Hydrobiol. , 29 : 664 – 686 .
- Gächter , R. and Meyer , J. S. 1993 . The role of microorganisms in mobilization and fixation of phosphorus in sediments . Hydrobiologia , 253 : 103 – 121 .
- Gächter , R. and Müller , B. 2003 . Why the phosphorus retention of lakes does not necessarily depend on the oxygen supply to their sediment surface . Limnol. Oceanogr. , 48 : 929 – 933 .
- Graneli , W. and Solander , D. 1988 . Influence of aquatic macrophytes on phosphorus cycling in lakes . Hydrobiologia , 170 : 245 – 266 .
- Håkanson , L. 2004 . Break-through in predictive modelling opens new possibilities for aquatic ecology and management – a review . Hydrobiologia , 518 : 135 – 157 .
- Hejzlar , J. , Šamalova , K. , Boers , P. and Kronvang , B. 2006 . Modelling phosphorus retention in lakes and reservoirs . Water Air Soil Pollut , 6 : 487 – 494 .
- Holdren , G. C. and Armstrong , D. E. 1980 . Factors affecting phosphorus release from intact lake sediment cores . Environ. Sci. Technol. , 14 : 79 – 87 .
- Hupfer , M. , Gloess , S. and Grossart , H.-P. 2007 . Polyphosphate-accumulating microorganisms in aquatic sediments . Aquat. Microb. Ecol. , 47 : 299 – 311 .
- Hupfer , M. and Lewandowski , J. 2005 . Retention and early diagenetic transformation of phosphorus in Lake Arendsee (Germany) – consequences for management strategies . Arch. Hydrobiol. , 164 : 143 – 167 .
- Hupfer , M. and Lewandowski , J. 2008 . Oxygen controls the phosphorus release from lake sediments – a long-lasting paradigm in limnology . Int. Rev. Hydrobiol. , 93 : 415 – 432 .
- Jensen , H. S. and Andersen , F. O. 1992 . Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of four shallow, eutrophic lakes . Limnol. Oceanogr. , 37 : 577 – 589 .
- Larsen , D. P. and Mercier , H. T. 1976 . Phosphorus retention capacity of lakes . J. Fish. Res. Board Can. , 33 : 1742 – 1750 .
- Lathrop , R. C. , Carpenter , S. R. and Robertson , D. M. 1999 . Summer water clarity responses to phosphorus, Daphnia grazing, and internal mixing in Lake Mendota . Limnol. Oceanogr. , 44 : 137 – 146 .
- Mataraza , L. K. and Cooke , G. D. 1997 . A test of a morphometric index to predict vertical phosphorus transport in lakes . Lake Reserv. Manage. , 13 : 328 – 337 .
- Moore , B. C. and Christensen , D. 2009 . Newman Lake restoration: A case study. Part I. Chemical and biological responses to phosphorus control . Lake Reserv. Manage. , 25 : 337 – 350 .
- Mortimer , C. H. 1941 . The exchange of dissolved substances between mud and water in lakes . J. Ecol. , 29 : 280 – 329 .
- Murphy , J. and Riley , J. P. 1962 . A single-solution method for the determination of phosphate in natural waters . Anal. Chim. Acta , 27 : 31 – 36 .
- Nürnberg , G. K. 1984a . Iron and hydrogen sulfide interference in the analysis of soluble reactive phosphorus in anoxic waters . Water Res. , 18 : 369 – 377 .
- Nürnberg , G. K. 1984b . The prediction of internal phosphorus load in lakes with anoxic hypolimnia . Limnol. Oceanogr. , 29 : 111 – 124 .
- Nürnberg , G. K. 1985 . Availability of phosphorus upwelling from iron-rich anoxic hypolimnia . Arch. Hydrobiol. , 104 : 459 – 476 .
- Nürnberg , G. K. 1987 . A comparison of internal phosphorus loads in lakes with anoxic hypolimnia: laboratory incubations versus hypolimnetic phosphorus accumulation . Limnol. Oceanogr. , 32 : 1160 – 1164 .
- Nürnberg , G. K. 1988 . Prediction of phosphorus release rates from total and reductant-soluble phosphorus in anoxic lake sediments . Can. J. Fish. Aquat. Sci. , 45 : 453 – 462 .
- Nürnberg , G. K. 1996 . Trophic state of clear and colored, soft- and hardwater lakes with special consideration of nutrients, anoxia, phytoplankton and fish . Lake Reserv. Manage. , 12 : 432 – 447 .
- Nürnberg , G. K. 1998 . Prediction of annual and seasonal phosphorus concentrations in stratified and polymictic lakes . Limnol. Oceanogr. , 43 : 1544 – 1552 .
- Nürnberg , G. K. 2004 . Quantified hypoxia and anoxia in lakes and reservoirs . TheScientificWorldJo , 4 : 42 – 54 .
- Nürnberg , G. K. 2005 . Quantification of internal phosphorus loading in polymictic lakes . Verh. Internat. Verein. Limnol. , 29 : 623 – 626 .
- Nürnberg , G. K. 2007 . Lake responses to long-term hypolimnetic withdrawal treatments . Lake Reserv. Manage. , 23 : 388 – 409 .
- Nürnberg , G. K. and LaZerte , B. D. 2001 . “ Predicting lake water quality ” . In Managing lakes and reservoirs , Edited by: Holdren , C. , Jones , W. and Taggart , J. 139 – 163 . Madison , WI : North American Lake Management Society, Terrene Institute in cooperation with Office Water Assessment Watershed Protection Division U. S. Environ. Prot. Agency .
- Nürnberg , G. K. , LaZerte , B. D. and Olding , D. D. 2003 . An artificially induced Planktothrix rubescens surface bloom in a small kettle lake in southern Ontario compared to blooms world-wide . Lake Reserv. Manage. , 19 : 307 – 322 .
- Nürnberg , G. K. and Peters , R. H. 1984a . Biological availability of soluble reactive phosphorus in anoxic and oxic freshwater . Can. J. Fish. Aquat. Sci. , 41 : 757 – 765 .
- Nürnberg , G. K. and Peters , R. H. 1984b . The importance of internal phosphorus load to the eutrophication of lakes with anoxic hypolimnia . Verh. Internat. Verein. Limnol. , 22 : 190 – 194 .
- Prairie , Y. T. 1989 . Statistical models for the estimation of net phosphorus sedimentation in lakes . Aquat. Sci. , 51 : 192 – 210 .
- Prairie , Y. , de Montigny , C. and del Giorgio , P. 2001 . Anaerobic phosphorus release from sediments: a paradigm revisited . Verh. Internat. Verein. Limnol. , 27 : 4013 – 4020 .
- Sas , H. 1989 . Lake restoration by reduction of nutrient loading: expectations, experiences, extrapolations , 497 Sankt Augustin , , Germany : Hans Richarz Publikations-Service .
- Schauser , I. and Chorus , I. 2007 . Assessment of internal and external lake restoration measures for two Berlin lakes . Lake Reserv. Manage. , 23 : 366 – 376 .
- Sereda , J. M. , Hudson , J. J. and McLoughlin , P. D. 2008 . General empirical models for predicting the release of nutrients by fish, with a comparison between detritivores and non-detritivores . Freshwater Biol. , 53 : 2133 – 2144 .
- Søndergaard , M. , Jensen , J. P. and Jeppesen , E. 2001 . Retention and internal loading of phosphorus in shallow, eutrophic lakes . TheScientificWorldJo , 1 : 427 – 442 .
- Søndergaard , M. , Jensen , J. P. and Jeppesen , E. 2005 . Seasonal response of nutrients to reduced phosphorus loading in 12 Danish lakes . Freshwater Biol. , 50 : 1605 – 1615 .
- Steinman , A. D. and Ogdahl , M. 2008 . Ecological effects after an alum treatment in Spring Lake, Michigan . J. Environ. Qual. , 37 : 22 – 29 .
- Straskraba , M. 1998 . Limnological differences between deep valley reservoirs and deep lakes . Int. Rev. Gesamt. Hydrobiol. , 83 : 1 – 12 .
- Vollenweider , R. A. 1976 . Advances in defining critical loading levels for phosphorus in lake eutrophication . Mem. Ist. Ital. Idrobiol. , 33 : 53 – 83 .
- Welch , E. B. and Jacoby , J. M. 2001 . On determining the principal source of phosphorus causing summer algal blooms in western Washington Lakes . Lake Reserv. Manage. , 17 : 55 – 65 .