
Abstract
With new direct dates from rock paintings comes an unprecedented opportunity to relate excavated archaeological data to the parietal record in southern Africa’s Maloti-Drakensberg Mountains. Anchoring dated art to recovered palaeoenvironmental, faunal and technological data enables the incorporation into socioecological models of ideational inferences, affording insights into how hunter-gatherers perceived their mountain habitats. Of particular interest is the late Holocene Neoglacial (∼3.5–2 kcal BP), during which skilled paintings were being made just as the region experienced dynamic changes owing in part to climate change. Responses of local foragers are evident across a range of cultural spheres, including dramatic subsistence transformations. With the Maloti-Drakensberg’s well-known “traditional corpus” of fine-line art now known to extend back to at least 3 kcal BP, here we explore how such changes may have precipitated – and in turn been influenced by – ontological shifts in relation to the food quest. As desirable game declined and hunting windows narrowed, we suggest that Neoglacial foragers sought to manage scheduling and social conflicts through enhanced spiritual negotiation with non-human entities in the landscape. Facilitated by the supernaturally charged nature of their elevated cosmos, this intensified spiritual labour may have found material expression in an elaborate new style of painting.
1. INTRODUCTION
Scholars of past hunter-gatherers have long sought to make the excavated and rock art records mutually intelligible. Difficulties dating rock art, however, together with incongruencies in the timescales to which each record speaks, bedevil most attempts to do so. In southern Africa’s Maloti-Drakensberg Mountains (), the potential power of articulating these records was understood from the outset of systematic research there. The pioneering rock art researcher, Patricia Vinnicombe (Citation1972:195), argued compellingly that the discordance between the frequency of animal taxa painted versus those recovered in excavation shows Maloti-Drakensberg societies did “not necessarily relate life to the rumblings of their stomach”. Her then husband, the “dirt” archaeologist Patrick Carter (Citation1978), listed the frequency of painted sites as among the main reasons for choosing southeastern Lesotho for his seminal doctoral study. Nonetheless, Vinnicombe and Carter’s efforts were hamstrung by an overall dearth of available data, competing interests, and, most intractably, the inability to chronologically anchor the rock art. There was a will, in other words, but not a way.
Figure 1. Map of southern Africa and Maloti-Drakensberg region with key sites/locales discussed in text or with Neoglacial-aged deposits. AT: Athol; BV: Belleview; BW: Bonawe; CW: Colwinton; D1: Diamond 1; EVL: eMvuleni; GD: Grootdrink; GH: Good Hope; GT: Groot Toren; JB: Jagersberg; KC: Kilchurn; KP: Kenegha Poort; KS: Kopshoring; LIK: Likoaeng; MF: MG: Mpongweni; Mafadi Summit; MH: Mahwaqa; PP: Popple Peak; NS: Njesuthi Summit; RSS: Rain Snake Shelter; SB: Strandberg; ST: Sani Top; SEH: Sehonghong; SK: Sekhokong; STA: Strathalan A; STB: Strathalan B; TK: Tierkloof; TN: Thabana Ntlenyana; TD: Tiffindell; TP: Tlaeeng Pass; WB: Wittberg; WV: Waterval.
Though much data has been generated in the intervening half century, the situation remains largely unchanged (Mitchell, Citation2009:99). Yet, success in this old endeavour is taking on a new urgency. Novel theoretical insights, rendered under the umbrella of the “New Animisms”, are making it increasingly clear that members of small-scale societies, and hunter-gatherers in particular, lived lives steeped in belief and religion. No aspect of their cultural system – not subsistence, mobility, technology, exchange, social relations, nor anything else – can be divorced from the ideational (Hallowell, Citation1960; Tanner, Citation1979; Bird-David, Citation1999; Viveiros de Castro, Citation2004; Willerslev, Citation2004; Hornborg, Citation2006; Wallis, Citation2009; Harvey, Citation2010; Descola, Citation2012; Halbmayer, Citation2012). These insights are born of both re-readings of traditional ethnographies and newly undertaken ethnographic studies across the globe. Southern Africa itself is one such region of active contribution, in which it can be shown that forager “spirituality” and “everyday life” are indivisible, especially since animals, spirits, people, ancestors and elements all have agency and personhood (e.g. McGranaghan and Challis, Citation2016; Guenther, Citation2020a,Citation2020b; Challis and Skinner, Citation2021; Skinner and Challis, Citation2022; Stewart, CitationIn press). It follows that archives of hunter-gatherer pasts in southern Africa are as much a product of spiritual life as they are of “making a living”.
For the San and their ancestors, these archives are threefold: (1) ethnography, (2) rock art, and (3) excavated archaeology. In a seminal study, Ouzman and Wadley (Citation1997) advocated for the integration, where possible, of these archives if we are to gain a fulsome picture of Holocene foragers in southern Africa. Taken together, their cumulative strength forms a cable of inference (cf. Wylie, Citation1989) that is greater, they argued, than the sum of its parts. Importantly, the second archive – rock art – can be deciphered with a relatively high degree of resolution when analysed in conjunction with the first – ethnography – and especially that pertaining to San beliefs. Most famously, David Lewis-Williams triangulated (e.g. Citation1998) between the ethnographic record of the twentieth century Northern (Kalahari) San, the historical nineteenth century record of the Southern (Karoo and Mountain) San, and the Maloti-Drakensberg rock art record in order to interpret the latter. The cohesiveness of the visual metaphors exposed and developed since has firmly established rock art as an independent pillar of ontological inference, especially when applied to the hyper-local. What is still lacking, with some notable exceptions (e.g. Ouzman and Wadley, Citation1997; Challis et al., Citation2008; Mazel, Citation2009; Kinahan, Citation2017), is a correspondence between archives 2 (rock art) and 3 (excavated evidence) (Mitchell, Citation2002; cf. Witelson, Citation2022). Both are exceptionally well preserved in the Maloti-Drakensberg, owing to the mountains’ inaccessibility and distance from the built environment.
Moving forward, and drawing inspiration from the New Animisms, we advocate in this paper for a new approach to integrating these three dimensions of San archive: using the lens of what we term environmental ontology to generate hypotheses of holistic cultural transformations in the past. Environmental ontology refers to the way in which the landscape would have been seen by indigenous people – the San, their ancestors and descendants – an ethnographically-informed indigenous perspective on the environment and its resources, especially relating to peoples’ relationships with the entities that embody and influence those resources (e.g. Challis, Citation2019; Guenther, Citation2020b; Challis and Skinner, Citation2021; Skinner and Challis, Citation2022; Stewart, CitationIn press). Because environments and their resource availabilities change, so foragers must continually broker relations with the non-human beings in their dynamic world. Working from the known to the unknown, we utilise ethnographic understandings allied with the hyper-local parietal (painted) archaeological record as an interpretive mesh to “sieve” spiritual insights from its sedimentary counterpart.
Specifically, we explore the spiritual ramifications of the Neoglacial (∼3.5–2 kcal BP), a late Holocene cold phase that corresponds with evidence for environmental and cultural dynamism in the Maloti-Drakensberg (Stewart and Mitchell, Citation2018a, Citation2018b). We show how fairly dramatic subsistence transformations, rooted in ecological change, may have precipitated – and in turn been facilitated by – shifts in San-specific environmental ontology. Mountains played a central role in San belief systems (Orpen, Citation1874; D. Bleek, Citation1933:310). Local and regional historical testimonies bring the mountains to life by suggesting that broken topography animated the cosmology of the San idiom. Such convictions are likely to have considerable time depth, as indicated by local rock art imagery (Challis, Citation2019). Yet belief systems, while perhaps not changing radically year to year, would likely have accommodated shifting socioecological conditions (McGranaghan, Citation2012, Citation2014a,Citation2014b; Guenther, Citation2015). Harnessing contextual data afforded by the excavated archaeological record, here we explore possible ontological impacts of environmentally-driven shifts in the abundance and structure of key foraging resources that lie at the core of such beliefs.
Central to our argument are polychrome (many-coloured) paintings of common eland (Taurotragus oryx), especially those rendered with the exquisite shading technique (), termed “shaded polychrome”. These are images that have brought world-renown to southern African rock art (e.g. Vinnicombe Citation1976). This technique seems to have begun contemporaneously across the Maloti-Drakensberg, appearing at the same point in the relative sequences (Mazel, Citation2009:92–3) of three research areas (Pager, Citation1971; Vinnicombe Citation1976; Russell, Citation2000). New direct chronometric dates for shaded polychrome eland place their origins firmly within the Neoglacial (Bonneau et al., Citation2017a), allowing us to articulate them with dated archaeological and paleoenvironmental records. We find that eland paintings become richly embellished just as this species goes from being the sole large prey to effectively disappearing from the highland archaeological record. Approaching these results with emic insights derived from San ontology and the knowledge that rock paintings are a negotiation with supernatural entities in the landscape (animals, the rain, the spirits), we contend that shaded polychromes’ inception during the Neoglacial is an outcome of intensified brokering of spiritual relations. More care and attention came to be afforded to the relationship with these entities, especially eland (Challis and Skinner, Citation2021), including its painted form. Indeed, we take our title, following Vinnicombe, from the Phuthi man, Mapote, who said he would paint an eland because the Mountain San were people “of the eland” (How, Citation1962:38).
Figure 2. Examples of shaded polychrome eland paintings from various Maloti-Drakensberg locales, including: (a) Sehlabathebe, highland Lesotho; (b) Mount Fletcher, Eastern Cape, where a shaded polychrome eland is clearly superimposed on non-shaded bichromes; and (c & d) Game Pass Shelter, KwaZulu-Natal, so-named on account of its position overlooking a historic game trail into and out of the highlands. Images by the authors.

Before detailing these shifts, we first outline today’s mountain environment and vegetation belts, as well as its historically known fauna. Next, we set the indigenous idiom of San belief in these mountains – the emic, or insider’s, view of the topography – before moving on to the Neoglacial environmental changes that would have prompted ideational responses.
2. ECOLOGICAL STRUCTURE: CLIMATE, VEGETATION AND FAUNA
The Maloti-Drakensberg Mountains cover nearly 40 000 km2 and constitute southern Africa’s most extensive highland zone (; Carbutt, Citation2019). They centre on the deeply dissected plateau of Lesotho and encompass adjacent parts of South Africa’s KwaZulu-Natal, Eastern Cape and Free State Provinces. The tallest peaks rise above 3000 m, with Thabana Ntlenyana (3482 m asl) in eastern Lesotho Africa’s highest south of Kilimanjaro. The mountains’ intricate network of drainage feeds southernmost Africa's largest river, the Orange, known in Lesotho as the Senqu. Rising in that nation’s high northeast, the Orange-Senqu first flows south, partitioning the Drakensberg range and Central range of the Maloti (), before heading west to cross over 2000 km of increasingly arid interior plateau to the Atlantic Ocean. East-flowing drainages along the uKhahlamba-Drakensberg Escarpment, which constitutes Lesotho’s eastern border with South Africa, form some of the largest rivers on the Indian Ocean coastline: the Thukela, Mkhomazi, Mzimkhulu, Mzimvubu and Kei.
2.1. Present-day climate
The current climate of the Maloti-Drakensberg region is continental. Summers are warm and humid, and winters dry and cool to cold. Situated in southern Africa’s summer rainfall zone, the region obtains roughly 80% of its annual rainfall between October and March in the form of high-intensity thunderstorms. Rainfall is highest along the uKhahlamba-Drakensberg Escarpment, where mean annual precipitation (MAP) can exceed 1500 mm and is augmented by orographic mists (Killick, Citation1963; Mucina and Rutherford, Citation2006; Schulze, Citation1979; Sene et al., Citation1998). The highland valleys in the rain shadow immediately west of the Escarpment see a sharp drop in precipitation (MAP: 500–600 mm), which increases again as one moves still further west towards lowland Lesotho (MAP: 700–800 mm) (Sene et al., Citation1998; Hydén, Citation2002).
Temperatures vary strongly by altitude, season and time of day. Mean annual temperatures (MAT) range from ∼15°C in lowland Lesotho to ∼6°C in the high Maloti-Drakensberg (Grab, Citation1994, Citation1997). Frost is widespread and ranges from ∼31 days per year in the lowlands to ∼150 days per year in the highlands (Schwabe, Citation1995). Frosts in the eastern highlands often exceed 200 days per year (Killick, Citation1978; Grab, Citation1997). While westerly cold fronts can bring snow to the highlands year-round, snowfalls usually occur from April to October with the heaviest incidences from June to August (Mulder and Grab, Citation2009). Due to their latitudinal position, the southern Maloti-Drakensberg are colder than the north and springtime (September) snowfalls there are both more frequent and can occur at lower altitudes (>2800 m asl) (Mulder and Grab, Citation2009:231). Dense fog can occur year-round and high mountains regularly experience thick orographic mists (Killick, Citation1963:196). Lightning activity in summer is exceptional even by global standards, with strikes on basalt summits violent enough to shatter bedrock and so contribute to landscape formation (Knight and Grab, Citation2014; Mayet et al., Citation2016).
2.2. Vegetation belts
The Maloti-Drakensberg region lies within southern Africa’s Grassland Biome, although its topography sets it apart from other grassland habitats. Grass communities in southern Africa are often differentiated according to palatability, or degree of nutritive benefit, to grazing animals. “Sweetveld” is dominated by grasses that remain palatable and sustain animal body condition through the year. They grow in relatively warm and dry environments on soils derived from acidic (e.g. sandstones, quartzites) to intermediate (e.g. granites, shales) parent material, but in mesic conditions when soils are more basic (e.g. volcanics) (Ellery et al., Citation1995; O’Connor and Bredenkamp, Citation1997). In contrast, “sourveld” occurs in areas where available moisture is high and soils are intermediate to basic, or in warmer and drier areas where soils are acidic. Sour grasses lose nutritive value after the growing season (Rowe-Rowe and Scotcher, Citation1986:14), leading to health declines should animals continue to forage exclusively on them in winter (Ellery et al., Citation1995; O’Connor and Bredenkamp, Citation1997). Complicating the situation is that while some grass species are either sour or sweet, many can be either depending on local conditions (Mentis and Huntley, Citation1982). In the mountainous Maloti-Drakensberg, cool temperatures and high rainfall on mostly basaltic substrates generally favour sour grasses. Nevertheless, there are strong gradients of palatability conditioned by altitude and aspect, as well as localised substrate-related anomalies.
Vegetation communities in the mountains occur as a series of altitudinally-stacked belts. On the western (Lesotho) side of the uKhahlamba-Drakensberg Escarpment, which constitutes the Maloti mountains and the vast bulk of the region (see ), the lowest altitude (∼1600–1900 m asl) areas occur along the terraces and valley slopes of the Orange-Senqu River and its tributaries. Today this fluvial network is covered with Senqu Montane Shrubland, characterised by diverse and abundant evergreen trees and shubs, and thicket in sheltered valleys (Mucina and Rutherford, Citation2006). Interspersed with the woody taxa are tall, heliophyte, perennial grasses; all are C4-photosynthesizing, and numbers are mildly to highly palatable, including species of Eragrostis, Hyparrhenia and Themeda. However, much of the vegetation comprising this unit stems from recent eastern intrusions of more xerophytic, lowland-affiliated taxa. Prior to the onset of highland settlement by isiNtu-speaking groups in the nineteenth century, Themeda triandra – a C4 species renowned for its superb pasturage (Tainton, Citation1981; Snyman et al., Citation2013) – was almost certainly dominant here (Staples and Hudson, Citation1938; Bawden and Carroll, Citation1968; Swallow et al., Citation1987).
On the overlying plateaus and high mountain ridges, at altitudes of ∼1900–2900 m, is Lesotho Highland Basalt Grassland (Mucina and Rutherford, Citation2006). This is a denser and shorter C4 grassland with Themeda triandra prevalent up to ∼2100 m on south-facing (cooler) slopes and ∼2700 m on north-facing (warmer) slopes. Sparse shrubs dominated by Passerina montana and Chrysocoma ciliata also occur, especially in disturbed areas. As with Senqu Montane Shrubland, however, these and numerous other pioneer taxa are considered to be recent invaders resulting from more than a century of overgrazing (Swallow et al., Citation1987:20; Mucina and Rutherford, Citation2006).
Above these altitudes, but still within the Lesotho Highland Basalt Grassland belt, a major ecological transition occurs from a predominance of C4 to one of C3 grasses. These are cold-adapted (Pooid) afroalpine grass types that dominate the highland landscape up to ∼2900 m asl, with species of Festuca and Merxmuellera the most prevalent. Shorter, harder and less sustaining to grazers than Themeda, they are palatable when young but quickly become unappealing and indigestible (letsiri or “sour”) after the relatively short growing season at these altitudes (Tainton, Citation1981). The ecological contrast between the two grasses is such that some advocate splitting this vegetation unit in two, with a montane belt (∼1900–2300 m; C4) below an upper subalpine belt (2300–2900 m; C3) (Morris et al., Citation1993). However, the large vertical overlap between Themeda and Festuca speaks to this unit’s cohesiveness as a montane belt, albeit one with complex, aspect- and altitude-driven vegetation gradients. Nevertheless, the changing vertical position of vegetation belts dominated by C3 versus C4 grasses has long been recognised as an important indicator of regional climate change (Vogel, Citation1983).
The Maloti-Drakensberg’s highest reaches (2900–3400 m asl) constitute southern Africa’s only alpine environment, and a well-known hotspot of African plant biodiversity and species endemism (van Wyk and Smith, Citation2001; Carbutt and Edwards, Citation2004, Citation2006, Citation2015; Carbutt, Citation2019). Here we find Drakensberg Afroalpine Heathland (Mucina and Rutherford, Citation2006), a fynbos-affiliated Erica-Helichrysum heath with hard-leaved, dwarf alpine bushes interspersed with short, tussocky Festuca-Merxmuellera grasses (all C3Pooids). The basalt-derived soils on which these temperate grasses mostly grow are heavily leached by high, regular rainfall at these altitudes, impoverishing their nutrient base and limiting grazing potential (Rowe-Rowe and Scotcher, Citation1986; Carbutt and Edwards, Citation2001, Citation2015; Grab and Nüsser, Citation2001; Mucina and Rutherford, Citation2006). As a consequence, the alpine zone today is used almost exclusively for small stock and only in summer when foliage is young. But even then, and in contrast to the rest of Lesotho, grazing pressure here is low (Quinlan and Morris, Citation1994). Summer is also when alpine forbs are available, whereas “in spring and late autumn there is very little in flower and in winter practically nothing” (Killick, Citation1963:91). Within the alpine zone are patches of streambank communities and peat-forming mires, which regulate headwater flow into the Orange-Senqu fluvial system (Jacot Guillarmod, Citation1971; van Zinderen Bakker and Werger, Citation1974; Killick, Citation1978, Citation1990; Grab, Citation1997). Grass and sedge communities on the mires offer this zone’s most palatable vegetation, which itself is subject to intense summertime overgrazing (Schwabe, Citation1995; Grab and Deschamps, Citation2004).
2.3. Mountain fauna
Until overhunting, farm fencing and habitat degradation in the nineteenth and twentieth centuries led to precipitous declines, the Maloti-Drakensberg teemed with animal life (Du Plessis, Citation1969; Ambrose, Citation2006; Grab and Nash, Citation2022). Many taxa were central to hunter-gatherer lifeways in the region, as evidenced from its Late Quaternary excavated and rock art records. The largest were dominated by a rich variety of grazing ungulates, the most common and persistent of which were black wildebeest (Connochaetes gnou), red hartebeest (Alcelaphus buselaphus), blesbok (Damaliscus pygargus phillipsi), plains zebra (Equus quagga) and quagga (Equus quagga quagga). Also noted by numerous historical observers (e.g. Clarke, Citation1888; Germond, Citation1967; Arbousset, Citation1991) were common eland, which came to hold acute significance both dietarily and in the spiritual life of the Mountain San (How, Citation1962; Vinnicombe Citation1976; Lewis-Williams and Dowson, Citation1989). Many medium- to small-sized antelope also maintained a long-term presence in the mountains, including grey rhebok (Pelea capreolus), oribi (Ourebia ourebi) and klipspringer (Oreotragus oreotragus), all three being resident year-round in the alpine belt, as well as springbok (Antidorcas marsupialis), mountain reedbuck (Redunca fulvorufula), steenbok (Raphicerus campestris), common duiker (Sylvicapra grimmia) (Plug, Citation1997; Plug and Mitchell, Citation2008).
Numerous other ungulates only appeared sporadically at various times as Late Quaternary climatic and associated habitat changes opened windows of opportunity, among them blue antelope (Hippotragus leucophaeus), roan antelope (Hippotragus equinus), blue wildebeest (Connochaetes gnou), bushbuck (Tragelaphus scriptus), impala (Aepyceros melampus), and blue and red duiker (Cephalophus monticola and natalensis) (Ambrose, Citation2006; Germond, Citation1967; Grab and Nash, Citation2022). A particularly marked change in species composition occurred in the highlands after the mid-Holocene, when a series of large grazers that had been regular components of archaeofaunal assemblages there disappeared, either largely or entirely. Likely dissuaded from penetrating the mountains by late Holocene temperature downturns, these taxa include roan antelope, quagga, plains zebra, springbok, blesbok, blue antelope and black wildebeest (Plug and Engela, Citation1992; Plug, Citation2003; Plug et al., Citation2003; Plug and Mitchell, Citation2008). In contrast, late Holocene faunal assemblages across the Maloti-Drakensberg region register an uptick in smaller browsing antelopes, signalling enhanced woody vegetation cover at this time (Opperman, Citation1987; Plug and Mitchell, Citation2008). In the highlands themselves, the only large ungulate to endure into the late Holocene in any abundance was eland, a mixed feeder (Plug et al., Citation2003:147).
Freshwater fish were another important dietary resource for past peoples in the Maloti-Drakensberg (Hobart, Citation2003; Plug and Mitchell, Citation2008; Plug et al., Citation2010; Stewart and Mitchell, Citation2018a). Fourteen species of indigenous fish currently exist in the upper Orange-Senqu system (Schrijvershof, Citation2015; Skelton, Citation2001), of which four were of particular interest to prehistoric people: three cyprinids – the largemouth yellowfish (Labeobarbus kimberleyensis), the smallmouth yellowfish (Labeobarbus aeneus) and the Orange River mudfish (Labeo capensis) – and the rock catfish (Austroglanis sclateri). Exploited intermittently from at least the late Middle Stone Age, the cyprinids in particular were heavily targeted by afromontane foragers due no doubt in part to their size. All are very large-bodied, with the largemouth yellowfish southern Africa’s largest scale-bearing indigenous fish, reaching fork lengths of 82.5 cm and weights of 22.2 kg (Arthington et al., Citation1999; Skelton, Citation2001). The smallmouth yellowfish and Orange River mudfish are smaller, but both can still reach fork lengths of 50 cm and weights of 9 and 3.8 kg, respectively (Skelton, Citation2001). Size estimates from archaeological specimens of all three cyprinid taxa in highland Lesotho have, moreover, exceeded these modern maxima (Plug, Citation2008). The smallmouth yellowfish is known for its hardiness and adaptability, thriving in colder waters and at higher altitudes (≥2200 m asl) than other taxa indigenous to the Orange-Vaal River system (Skelton, Citation2001; Gerber et al., Citation2012). Fishing scenes in Maloti-Drakensberg rock art provide some indication of the capture techniques, which included spearing and leistering from small rafts, as well as diverse forms of mass capture involving nets, baskets and/or drag screens (Stewart and Mitchell, Citation2018a).
2.4. Game and fish migrations
A number of the region’s grazers – black and blue wildebeest, springbok, blesbok and the two equids (quagga and plains zebra) – appear to have been collective migrants into the Maloti-Drakensberg region. Documentary and oral historical data indicate that aggregated herds of these ungulates once migrated across the southern Highveld, in numbers potentially rivalling East Africa’s famous Serengeti-Mara migrations (Harris et al., Citation2009; Boshoff and Kerley, Citation2013, Citation2015; Boshoff et al., Citation2016; Grab and Nash, Citation2022). While the collapse of these systems in the nineteenth century makes specific routes difficult to reconstruct, a broad migratory pattern has emerged. Specifically, herds of large ungulates appear to have traversed rainfall gradients on a seasonal basis. In the autumn and winter months they were observed grazing sweetveld at the relatively low-lying margins of the Highveld and beyond, including the eastern and northern Free State, the southeastern North West Province, and the KwaZulu-Natal midlands. The spring and summer months saw them move to the Highveld’s higher, cooler, better-watered plateau regions, where sourveld grassland offers palatable new forage (Boshoff and Kerley, Citation2013, Citation2015).
Bounding the Highveld’s southeastern margin, the Front Range of the Maloti has been argued to have presented these dry plains-adapted migrants with something of a habitat edge, with the uKhahlamba-Drakensberg Escarpment beyond forming a biogeographic barrier (; Grab and Nash, Citation2022). However, zooarchaeological remains from the Lesotho Highlands show that these plains species, throughout the Late Quaternary, did filter into the highlands (Carter, Citation1978; Plug and Badenhorst, Citation2001; Plug et al., Citation2003; Plug and Mitchell, Citation2008; Boshoff and Kerley, Citation2013; Boshoff et al., Citation2016), at times reaching high altitudes >2200 m asl (Badenhorst et al., Citation2019). Drawing them in was likely the burst of nutritious sourveld grazing that becomes available in the growing season. As spring/summer warmth progressively greened the grasses in an upslope direction – primarily the low- and mid-altitude C4 Themeda belts and to a lesser extent the high-altitude C3 Festuca-Merxmuellera zones – animals would have increased their elevation. The subsequent downwards “souring” of the mountain grasses with the coming of autumn pushed these grazers back to lower elevations surrounding the Maloti-Drakensberg and in places off the Highveld altogether.
Eland and red hartebeest were year-round residents of the region rather than seasonal visitors. These species have wider ecological tolerances than the plains taxa just discussed, including cool, topographically broken uplands (Skinner and Chimimba, Citation2005). Vibrant populations were encountered as early nineteenth century Europeans settled the mountain peripheries (Arbousset, Citation1991) before the former retreated into the mountain core “for which [they have a] decided preference” (Arbousset and Daumas, Citation1846:4). Nevertheless, both historical and more contemporary data suggest that these taxa, too, took advantage of seasonal windows of high-quality pasturage at altitude. Boshoff and Kerley (Citation2013:295), for example, found historical instances of “local, altitudinal, migrations of [eland and red hartebeest] from high and exposed plateaux to deep, sheltered, neighbouring valleys … in central and eastern Lesotho” on a seasonal basis. Similar movements still take place on the South African side of (and over) the Escarpment, where once-threatened eland populations now thrive thanks to the early establishment of protected areas.
Large terrestrial game animals were not the only faunal resources to make use of the highlands on a seasonal basis; the large cyprinids – the largemouth and smallmouth yellowfishes – were also seasonal visitors. Before hydroelectric damming transformed the flow and temperature dynamics of the Orange-Vaal River system, adults of these two Leobarbus species would migrate far upstream from the middle Orange in the Karoo to the river’s upper reaches in highland Lesotho in order to breed (Shortt-Smith, Citation1963; Jubb, Citation1966). Here, they would spawn on gravel or sand beds in swiftly moving, well-oxygenated waters (riffles and rapids) of the main river channel and larger tributaries (Mulder, Citation1973; Cambray, Citation1985; Skelton, Citation2001). For the hardier smallmouth yellowfish, these coordinated mass annual spawning runs are triggered by the downstream arrival of floodwaters with the first spring rains (Tómasson et al., Citation1984). Their largemouth counterparts followed some four to six weeks later, as water temperatures warmed. For the early-migrating smallmouth yellowfish that season can last from October to January, whereas for their latecomer largemouth counterparts spawning typically takes place in January and February (Mulder, Citation1973; Tómasson et al., Citation1984). In contrast, the Orange River mudfish and the rock catfish – the other two fish species exploited regularly by afromontane foragers – do not appear to undertake long-distance longitudinal migrations, although the former does expand laterally with early spring to summer rains to lay eggs on newly inundated highland floodplains (Mulder, Citation1973; Gaigher et al., Citation1980; Tómasson et al., Citation1984).
3. COSMOLOGICAL STRUCTURE: POTENCY NEGOTIATION AND HIGH-ALTITUDE HUNTING
How can we know how hunter-gatherers might have perceived the Maloti-Drakensberg? Owing to the work of nineteenth-century linguists Lucy Lloyd and Wilhelm Bleek, we have around 13 000 pages of verbatim San testimony from the |Xam San of the arid interior – some of which pertain to how mountains, such as the “Wittberg” in the southwestern Cape, were conceptualised (LL.II.-25.2239; Bleek and Lloyd, Citation1911; Hollmann, Citation2022:268). Notably, we have one hyper-local attestation from the Maloti by the Mountain San man, Qing, recorded in 1873 by Joseph Orpen (Orpen, Citation1874). The only testimony of its kind, it details the location of deities at altitude, their proclivities, and the legends in which they affect the lives of people and animals.
Beliefs of the |Xam San accord, in multiple instances of “fit”, with the words of Qing in the Maloti (e.g. McGranaghan et al., Citation2013). Much of the concordance between ethnographic texts has to do with the supernatural control of the weather – in particular “rain-making” or rain control, by way of negotiating with, or influencing, the water deity, !Khwa, and the creatures in which it was manifest or with which it was associated. These !Khwa-ka-xoro or “rain’s animals” include “Water Bulls” or “Water Cows” (he-rain or she-rain) that can take the form of bovids and hippopotami with manes and tusks (Lewis-Williams and Pearce, Citation2004); snakes such as cobras (Naja sp. or H. haemachatus) and puff adders (Bitis arietans) that come out in the rainy season, or Water/Rain Snakes of enormous stature and power (Orpen, Citation1874; Schmidt, Citation1979; Hoff, Citation1997, Citation1998; Mallen, Citation2005; Challis et al., Citation2013; McGranaghan and Challis, Citation2016; Skinner, Citation2021; Skinner and Challis, Citation2022).
Allied to this, and other startlingly correlative instances of fit, is the San conception of the world and how it is mapped out in the mind, not only among the southern San (the aforementioned |Xam and Qing) but also the northern San of the Kalahari. Famously fluid and changeable (e.g. Guenther, Citation1999), San cosmology exhibits plenty of features common to all groups (e.g. Barnard, Citation1988). At the regularly-held healing dances, San doctors push themselves into ecstatic states whereby they fall unconscious and their spirits travel downwards into waterholes or rivers, where they might deal with snakes and rain’s animals, and upwards along threads-of-light into the sky, so that they may plead with their god for the souls of the sick, to influence the rain, or to influence the movements of the game animals (Lewis-Williams et al., Citation2000). Importantly, contact with the spirits can only be achieved with the power of certain potent animals – the eland being chief among these – whose essence the women summon, in song and rhythm, at the dances for the men to harness (Orpen, Citation1874; Marshall, Citation1969; Lewis-Williams and Biesele, Citation1978; Katz, Citation1982; Biesele, Citation1993). As well as symbolising game, therefore, eland equate to healing and rain-making. Importantly, the eland is one of the animals whose form the rain takes (Lewis-Williams, Citation1981:106; Citation2000:222–3).
3.1. Rain-making in an elevated cosmos
Influence over rain, and weather more generally, was often performed at elevation: the |Xam informant, healer and rainmaker, ||Kabbo, spoke of a grandfather talking to his grandson (LL.II.-24.2213-2226; D. Bleek, Citation1933:309–310) thus: “I will milk a she-rain, I will cut her, by cutting her I will let the rain’s blood flow out so that it runs along the ground … to make all places wet”. The grandson replied: “I understand, for the she-rain is drawing her breath which resembles mist; you must please go and cut the rain at the great waterpits which are on the mountain”. To which the old man said:
I will really ride the rain up the mountain on top of which I always cut the rain. It is high, so the rain’s blood falls down.
These associations are not limited to the |Xam. In Botswana, a Ju|'hoãn informant told Megan Biesele (Citation1974:3) that the weather originates from the Tsodilo Hills, inside which the rain clouds dwell and from which they emanate. Sigrid Schmidt (Citation1979:207) was told by Nama and Damara informants that Brukkaros Mountain in Namibia should avoid being climbed by women on account of a huge white snake that lived there. More recently, Andrew Skinner (Citation2021, Citation2022a) has found that today’s inhabitants of the Maloti-Drakensberg, the Sesotho-, Sephuthi- and isiXhosa-speakers who inherited the mountains from the San, have inherited the local idiom of water and rain beliefs, and mountain river pools are no exception. Like the San, they describe the “owner of the pool”, (Sesotho: Khanyapa-fito), as variously an enormous “river snake” (cf. noha ea metsi, Rakotsoane, Citation1996, Citation2008) giving names for numerous adders, cobras and pythons, as well as water monitors and, in one instance, hippopotamus. Importantly, for the San and their descendents these entities are “persons” – non-human persons – with whom specialists establish and broker relations (Challis and Skinner, Citation2021). Such entities are known, not by what they are in western taxonomic terms, but in terms of where they are and by their actions; equally, the specialists who deal with them are known by their expertise in behaving properly towards them (Skinner, Citation2021; Challis and Skinner, Citation2021; Skinner, Citation2022b; Skinner and Challis, Citation2022). Among the |Xam, those able to influence the rain were known as !Khwa-ka-!gi:tən (D. Bleek, Citation1933:377; cf. Hollmann, Citation2022).
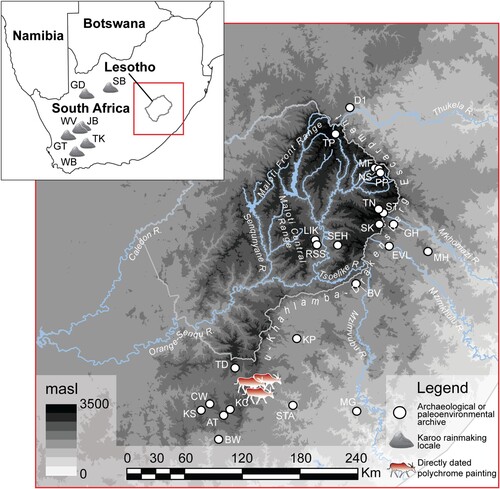
The same goes for specialist hunters, known by the |Xam as ʘpwai:tən-ta-!gi:tən, who influence the movements of the game animals. Only by acting “nicely” and with “understanding”, observing all ritual protocols and using respect words on the hunting ground, can a hunter induce calm or “tame” behaviour so that animals might be caught (McGranaghan and Challis, Citation2016). Negotiating one’s place among entities in the landscape is key to navigating it; the physical topography is a social topography and so, in the rock art of previous millennia, we see a forager idiom of negotiation-as-navigation (Challis and Skinner, Citation2021:20). From Qing’s testimony (Orpen, Citation1874:10), we learn that those who were able to “tame” the game animals and control their potency were also those who were called upon to “tame” snakes and other creatures of the rain () – in other words, to influence the weather. !Khwa, in animal form, whether bovid or serpentine, had to be stalked, placated and captured, much as if it were a game animal. So-called “rain's animals" (or rain-animals) are a common feature of the religious San rock art of the Maloti-Drakensberg, and are typically depicted being approached, placated and captured – drawn out through cracks and fissures (from which water runs down the rockface following heavy rains) (). In the highlands, moreover, they tend to occur in shelters in the valley bottoms overlooking river pools (Challis et al., Citation2013; Skinner and Challis, Citation2022).
Figure 3. In (a) rhebok-headed tamers-of-the-game (left) approach a dangerous tusked rain serpent (top) painted to suggest that it emerges from behind the rock face, while other transforming rhebok-headed dancers interact with a smaller “tamed” snake – knowable by its rhebok head. In (b) a tamer-of-the-game with human body and rhebok head, surrounded by eland, bleeds from the nose and carries a hunter’s bow, while a rhebok emerges from a shadowy step in the rock face. Note the white spoor/hoof prints. Images by Challis.
3.2. Taming the game in God's house: rock art and landscape
On the South African side of the Escarpment rock art shelters are commonly located in the sandstone cliffs overlooking the passes (). Eland move through these passes on their annual summer migration into the high Maloti of what is today Lesotho (other eland once moved up from Lesotho’s lowlands). The San, following the eland up through the passes and through the otherwise impenetrable mist, desired eland for their peerless fat content – the supernatural potency or essence harnessed at the dance, sometimes immediately following a fresh kill. Following the eland though the mist and up into the altitudinous hinterland was a pilgrimage both physical and spiritual (Lewis-Williams and Challis, Citation2011; ).
Figure 4. Eland antelope move through a southern Drakensberg valley towards the high passes into the spiritual hinterland of the high Maloti, where the San deity, |Kaggen, lives. They concentrated in large numbers, drawing hunter-gatherers to these altitudinous locales in a spring-summer aggregation phase that required heightened social and spiritual negotiation. Image by Challis.

We can safely say that seasonal migrations of eland would have aggregated in highland Lesotho’s pastures in their hundreds, if not thousands (cf. Carter, Citation1970, Citation1978:229; Vinnicombe, Citation1976; Rowe-Rowe, Citation1983:5; Plug, Citation2017:390). Indeed, today, reintroductions in the southern Drakensberg conservation area have led to sightings of herds up to 120–160 strong, feeding up high when there are good rains and returning to the valleys in autumn (D. Guy pers. comm.). To interpret such a concentration of eland from the perspective of the San, it would have represented a superabundance of fat and meat, and therefore of supernatural potency. Like electricity, it was desirable if controlled correctly but dangerous if mishandled (Marshall, Citation1969:351). Large concentrations of eland at altitude betokened the presence of the San god, |Kaggen (which Joseph Orpen recorded as Cagn), as Qing said: “have you not hunted and heard his cry, when the elands run to his call? Where Cagn is, eland are in droves like cattle” (Orpen, Citation1874:3). Such locations, having potency, would therefore have been both powerful and socially dangerous.
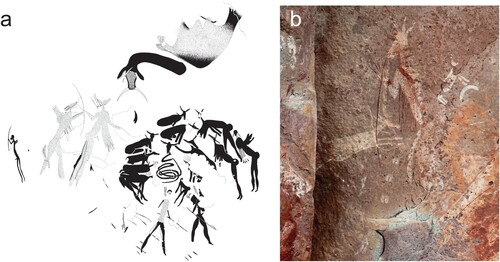
One example of excess potency comes from a particularly altitudinous painting in the Sehlabathebe National Park. Occuring at 2387m, it overlooks one of the most renowned grazing valleys in the region (R. Mokhachane pers. comm.) to which eland still venture up from below the Escarpment (Challis, Citation2019:177). The painting depicts perhaps the largest anthropomorph in the subcontinent and one which arguably cautions against the “monstrous” repercussions of excess consumption and the mishandling of potency (; Challis, Citation2019; cf. Mullen, Citation2018). A summer shelter (winter temperatures here dropping to below −20°C), different family bands would likely have congregated here seasonally when the eland arrived (Carter, Citation1970). The superabundance of fat and potent eland would bring people together, but the aggregation of extended family and friends would exacerbate tensions. Resource distribution, of meat in particular, gives rise to accusations of gluttony, causing jealousy, antagonism and fighting (Marshall, Citation1976, 287–312; Citation1999, 65, 94), which is a serious threat to survival in an otherwise un-policed society. Showing respectful understanding, especially amongst extended family – known as !k”werri:tǝnn the |Xam San (D. Bleek, Citation1956:510; McGranaghan, Citation2012, 193–194) would have been paramount. People whose specialty it was to mediate with such forces for the good of the community would therefore have been called upon, and healing dances would be held (e.g. Katz, Citation1982:37–38).
Figure 5. In Lesotho’s Sehlabathebe National Park is a 1m long painting of a ritual specialist heralding the potential dangers of a superabundance of resources that concentrate seasonally at altitude (Challis Citation2019). With bulging stomach (evoking associations of gluttony and poor resource distribution), tusks, and three legs with clawed toes (evoking leonine proportions of both power and gluttony), the figure in question may represent just such an instance of the strong ritual specialist struggling to control excess fat and potency while dealing with the entities above and below the water. Image courtesy of James Pugin.

Those who tamed eland (the game) and snakes (the rain) were able to harness antelope potency in the dances, enabling them to influence (and sometimes capture) these entities. If the potency is too great, however, healers can lose control of themselves and, with an undesirable surfeit of power, become lions: growing lion hair, lashing out and biting their fellow dancers (D. Bleek, Citation1935; Hollmann, Citation2022, 277). Danger not only to the individual, but danger to society, is apparent in this painting. Mark McGranaghan (Citation2012, Citation2014a, Citation2014b) has highlighted the recurrence of “animal” qualities referenced in the narratives of the southern San, and lions are exemplars of behavioural impropriety – |ke||ke, “beasts of prey” – who did not act “nicely”, who swallowed down food quickly and who did not share (LL.II.20.1844, 1846, 1862; LL.II.30.2695; LL.VIII.29.8554’). Furthermore, failure to broker relations properly and respectfully with the eland and with the rain's animals would be read as “lacking understanding” and deemed “monstrous” behaviour (McGranaghan, Citation2014a; Skinner and Challis, Citation2022).
The painted anthropomorph has three legs (likely to reference the experience of polymelia in trance) each with clawed toes, and one arm with clawed fingers (). It is covered in long hair – the symbol of the beasts of prey –and has protruding teeth or tusks and lion’s whiskers (Mullen, Citation2018). It also has a bulbous stomach to signify gluttony (; McGranaghan, Citation2014a). This image, then, most probably reflects the concerns of game specialists who, having achieved the altitude that put them in the rarefied atmosphere of the gods, were struggling to maintain their humanity and not become monstrous, either socially or physically, in the presence of so much supernatural power (Challis, Citation2019). In terms of social consequences, the image betokens what the place does (its character or “who” it is) and when things happen there (Challis and Skinner, Citation2021; Skinner, Citation2022b). The timing of one’s presence in a place is key to navigation, and being at the right place at the right time is key to negotiating with entities such as eland and rain (Skinner and Challis, Citation2022).
3.3. Fishing in the rain
The Maloti-Drakensberg was arguably the San tiered cosmos manifest, giving physical form to the axis mundi by reaching up into the sky (cf. Lewis-Williams and Challis, Citation2011:172–180). Far from being a simple up/down axis, however, the spirits can be encountered anywhere unfamiliar, dangerous or liminal. Travelling far away from camp across the hunting ground, for instance, was seen as a guarantee that beasts of prey, spirits-of-the-dead and other entities will be encountered; entities with whom negotiations had not been brokered (cf. Mguni, Citation2009; McGranaghan, Citation2012, Citation2014a,Citation2014b; Skinner, Citation2017; Challis and Skinner, Citation2021; Skinner, Citation2022a,Citation2022b; Skinner and Challis, Citation2022).
Both northern and southern San perceive healers to be able to mediate with, and travel to and from, other places in the cosmos (e.g. Katz, Citation1982; Biesele, Citation1993; Marshall, Citation1999). These other places can also be reached by going downwards, underground, or into rivers and waterholes where !Khwa can be found, and mountain river pools are no exception (cf. Skinner, Citation2021, Citation2022a). San healers go into trance, travel downwards, underwater, in order to interact with entities such as the Water Snake and other rain’s animals as well as other snakes, bovids, freshwater crabs, eels, reptiles and fish (see Lewis-Williams and Pearce, Citation2004). Importantly, this is in the same sphere of ritual activity as the aforementioned “tamers” of the rain, the !Khwa-ka-!gi:tən. Those able to approach the Water Snake, !Khwa, nicely, having established proper relations with it, could influence it and its creatures – that is to say the rain and its resources.
One of the better-known painted examples of a rain's animal being tamed, and captured, is on a high and steep-sided outlier of the uKhahlamba in Underberg, “eMvuleni” or “the rain’s place”, known in English as Bamboo Mountain. In a neighbouring shelter there is a depiction of a fish trap. Indeed rain's animals and fish are often found together (see below; Challis et al., Citation2008). Across the Sani Pass from this location is perhaps the best-known example of all: the rain’s animal capture panel at Sehonghong. It was here that Qing – pointing at the bovid beast being captured – said “that animal that the men are catching is a snake (!)” (Orpen, Citation1874:10, emphasis and exclamation mark in original), thus confirming its designation as a creature of the rain. This was independently corroborated by Bleek and Lloyd’s informant Diä!kwain in the Cape (Bleek in Orpen, Citation1874:12), and later by the rediscovered image of a serpent being captured in much the same manner on the opposite bank of the Senqu River (; Challis et al., Citation2013). Qing further said,
They are all under water, and those strokes are things growing under water. They are people spoilt by the – dance,* because their noses bleed (Orpen, Citation1874:10, the asterisk is for a footnote explaining the dance of blood [the nosebleeds induced by hyperventilation]).
Figure 6. This large image – at Rain Snake Shelter on the banks of the Senqu close to Likoaeng – perhaps best displays !Khwa, the rain in the form of the giant serpent. Tamers-of-the-rain, some in dancing postures, are bending forwards and bleeding from the nose while others clap and hold out “charms” to placate the animal while it is led by a line to the nose. At its fattest point, the serpent’s body has six or seven “cut” marks, which parallel the incised cuts on the rock immediately below the painting (Challis et al., Citation2013). The cliff line in which the shelter is formed is also engraved with many cut marks, as if the place itself were the snake (Skinner and Challis, Citation2022; cf. Hollmann, Citation2007).

These mountains receive some of the highest rainfall in the subcontinent. Why, then, should there be so many rain's animals painted at altitude? The reason is arguably connected with the timing (and taming) of the rains, not simply with rain-making per se. In this area, at high altitude and in the presence of the gods, resources could be excessively plentiful – even dangerously so – but the right conditions had to obtain at exactly the right time in order for this to happen. Adjacent to the aforementioned open-air fishing site at Likoaeng in the southern Lesotho Highlands, a small overhang exhibits key imagery in this regard. Specifically, one painting exhibits strong indications, per Challis et al. (Citation2008), that it was made in order to summon the rain and so induce the running of the fish (). The image depicts a dancing figure, bending forward and clapping, with a dying shaded polychrome eland that bleeds from the nose, setting the figures unequivocally in the context of the trance dance. Eland potency thus acquired then enabled the capture of the rain’s animal, depicted moving towards the net or trap filled with fish-shaped flecks (). Because we lack an understanding of when the Likoaeng paintings were executed, their environmental and social context is ambiguous, as is whether and how they articulate with the fish-rich site immediately adjacent. However, new dates for shaded polychrome eland in the wider Maloti-Drakensberg suggest that they originated during the Neoglacial (Bonneau et al., Citation2017a,Citation2017b, Citation2022), the climatic phase to which we now turn.
Figure 7. Rock art (a & b) and excavated archaeology (c) at Likoaeng, where millions of fish remains were recovered from Neoglacial-aged deposits. In a ravine immediately around the corner from the deposits, a rock art panel shows a dying eland and dancer ∼25 cm away from a probable fishing net and rain’s animal. The paintings are interpreted as depicting the harnessing of eland potency while in trance to summon the rain and thereby induce the seasonal running of migratory fish. Images courtesy of Peter Mitchell.

4. THE NEOGLACIAL: CLIMATIC, ENVIRONMENTAL AND CULTURAL CHANGES
After a relatively warm and variably humid mid-Holocene, southern Africa after ∼4.5 kcal BP experienced a series of cool episodes. These entailed northward shifts of westerly frontal systems and strengthened atmospheric circulation leading to widespread temperature reductions and enhanced winter rainfall (Jerardino, Citation1995). The most enduring and widely registered of these was the Neoglacial, which lasted roughly one and a half millennia from ∼3.5 to 2 kcal BP. Evident in archives across the subcontinent (Talma and Vogel, Citation1992; Jerardino, Citation1995; Holmgren et al., Citation1999, Citation2003; Lee-Thorp et al., Citation2001; Scott et al., Citation2003; Nash and Meadows, Citation2012), this phase brought lower temperatures and higher (annual and/or winter) precipitation to the Maloti-Drakensberg (Fitchett et al., Citation2016a; Stewart and Mitchell, Citation2018b; Herbert and Fitchett, Citation2022). Here we first review evidence for the Neoglacial’s expression in the mountain zone, before outlining its potential impacts across diverse cultural spheres. All radiocarbon ages provided have been calibrated using the SHCal20 Southern Hemisphere Calibration Curve (Hogg et al., Citation2020). Chronological resolution is limited in the latter half of the timeframe under investigation by a notable radiocarbon plateau ∼2580–2430 radiocarbon years BP (the so-called Hallstatt Plateau).
4.1. Pollen, peat and geomorphology
Mafadi Summit (3390 m asl), on the Lesotho side of the northern Escarpment, is southern Africa’s highest wetland. Perhaps on account of its altitude, the site’s pollen and diatom records together suggest that cold and humid conditions characterised the entire period ∼4.5–2 kcal BP (Fitchett et al., Citation2017). Cold-tolerant Fragilaria and Aulacoseira diatoms spike at this time relative to the mid- or terminal Holocene, while frequencies of Poaceae (grasses) and Apiaceae (umbellifers) pollen increase at the expense of Asteraceae (forbs) and Cyperaceae (sedges). All point to the existence of a cold, shallow lake in alpine grasslands under relatively harsh conditions (Fitchett et al., Citation2017:645).
Other high-elevation wetlands inform more specifically on the ∼3.5–2 kcal BP episode itself. Sedimentary, diatom and pollen data from the Sekhokong Range on Lesotho’s southern Escarpment indicate that the period ∼3.4–1.2 kcal BP was persistently cooler than before. Wet phases are registered ∼3.26–3.19, ∼3.05 and ∼2.69–1.47 kcal BP, the last of which may have seen more regular snowfalls (Fitchett et al., Citation2016b). Likewise, increases in Poaceae, Aponogeton and sedge pollen relative to Asteraceae signal greater humidity after 3.5 kcal BP at the Mahwaqa wetland, located on an Escarpment outlier in KwaZulu-Natal's Midlands (Neumann et al., Citation2014). At the Catchment IV wetland on Cathedral Peak (South African northern Escarpment), forest pollen (mostly Podocarpus) are reasonably common until ∼3.8 kcal BP. Their subsequent decline and the onset of a heavier dominance of Poaceae pollen potentially speaks to a downslope forest retreat, a scenario bolstered by increases in cold-hardy heath and shrub taxa like Urginea and Leucosidea sericea, respectively (Lodder et al., Citation2018). A sharp spike in moisture-loving Leucosidea pollen at ∼2.8 kcal BP may signal that enhanced cold was accompanied by greater humidity. Broadly similar conclusions were reached by Herbert and Fitchett (Citation2022), who recently compiled and analysed these and other pollen datasets from across the Maloti-Drakensberg spanning the past ∼8000 years.
Geomorphological evidence, though less temporally-resolved and often discontinuous, mostly corroborates this picture of Neoglacial conditions in the mountain zone. At Sani Top near Sekhokong, for example, sedimentological analyses of cirque deposits revealed renewed peat deposition, after a previous early Holocene episode, between ∼3.3 and 2.3 kcal BP (Marker, Citation1994). These deposits are broadly contemporaneous with peats bracketed more coarsely to between ∼5 and 1 kcal BP at Tlaeeng Pass, further north along the Escarpment (Hanvey and Marker, Citation1992). In the Eastern Cape Drakensberg to the region’s far south, various valley sediments have yielded 14C dates of interest. Overbank flood sediments and paleosols atop high-energy river gravels were all deposited sometime prior to ∼2.4 kcal BP at Kilchurn, while at nearby Athol bedded gravels were deposited earlier than ∼2.35 kcal BP (Lewis, Citation2005:41). When this fluvial activity began is unclear, but Lewis (Citation2005) highlights the existence of deep slopewash deposits and paleosols post-dating ∼3.2 kcal BP at Kopshoring and evidence of gully erosion occurring after ∼2.8 kcal BP at Tiffindell (Rosen et al., Citation1999). Geomorphological proxies of temperature depressions are more elusive, but Grab (Citation2000:186) wonders whether, given their fresh appearances, periglacial stone-banked lobes on Popple Peak, Njesuthi Summit and Thabana Ntlenyana (all 3400–3300 m asl) were “active during Late Holocene cold periods”. Reinforcing this is a minimum age of a protalus rampart on Thabana Ntlenyana of cal AD 248–348, demonstrating that some periglacial landforms were indeed generated as recently as the late Holocene (Grab and Mills, Citation2011).
4.2. Ecological and dietary shifts at Likoaeng
Archaeological data also suffer from temporal discontinuities, but with adequate preservation they, too, can shed valuable light on prevailing environmental conditions during phases of deposition. Probably the best record of Neoglacial environments in the mountain core comes from the site of Likoaeng in the southern Lesotho Highlands (Mitchell et al., Citation2011; Parker et al., Citation2011). Multiple proxies – phytoliths, δ13C of sediment organic matter (δ13CSOM), charcoals and faunal remains – contribute to a picture of pronounced ecological changes to highland Lesotho’s Senqu Valley during the third millennium BP (). They show that the site’s earliest occupational phase (Phase A), with 14C ages spanning 3.4–2.9 kcal BP, was deposited when the local landscape was not dissimilar to today – a C4 (likely Themeda triandra)-dominated grassland with minor contributions of C3 elements. This contrasts sharply with the subsequent phase (Phase B), deposited during the height of the Neoglacial ∼2.9–2.16 kcal BP ().
Figure 8. Selected palaeoenvironmental proxies (Parker et al., Citation2011) and faunal indices (Stewart and Mitchell, Citation2018a) from the archaeological sequence at Likoaeng, highland Lesotho, discussed in the text: (a) percentages of phytolith short cell morphotypes diagnostic of different grass types (Pooid C3, Panicoid C4 and Chloridoid C4); (b) percentages of bulliform phytolith morphotypes, argued to be preferentially laid down under conditions of enhanced humidity (Sangster and Parry, Citation1969; Andrejko and Cohen, Citation1984); (c) Likoaeng phytolith aridity index (Iph% = Chloridoid / Chloridoid + Panicoid), which provides a measure of xeric versus mesic grassland composition (Diester-Haas et al., Citation1973); (d) Likoaeng phytolith dicotyledon/Poaceae (D/P) ratio, a measure of woody (ligneous dicotyledon morphotypes) to grassy (Poaceae morphotypes) vegetation abundance and thus community openness (1 to 0, with 1 representing maximum woodland coverage and 0 none; Alexandre et al. Citation1997); (e) Likoaeng δ13C of sediment organic matter (δ13CSOM); (f) Likoaeng percentage proportions of C3 (versus C4) from δ13CSOM values (% of C3 plants = (δ13C −2 + 12.5)/−0.14; Wedin et al., Citation1995; after Bousman, Citation1991); (g) Likoaeng fish/mammal index (Σ NISP all fish / Σ [NISP all fish + NISP all mammals; 1 to 0, with 1 representing only fish and 0 only mammals]; Stewart and Mitchell, Citation2018a); (h) Likoaeng large ungulate/mammal index (Σ NISP all large ungulates / Σ [NISP all large ungulates + NISP all other mammals]; 1 to 0, with 1 representing only large ungulates and 0 only other mammals). The age/depth model employed is that found in Parker et al., Citation2011. The chronological boundaries of the late Holocene Neoglacial sensu lato can be seen in shaded blue, with the environmental and zooarchaeological inflection point that we identify at ∼2.6 ka highlighted.
![Figure 8. Selected palaeoenvironmental proxies (Parker et al., Citation2011) and faunal indices (Stewart and Mitchell, Citation2018a) from the archaeological sequence at Likoaeng, highland Lesotho, discussed in the text: (a) percentages of phytolith short cell morphotypes diagnostic of different grass types (Pooid C3, Panicoid C4 and Chloridoid C4); (b) percentages of bulliform phytolith morphotypes, argued to be preferentially laid down under conditions of enhanced humidity (Sangster and Parry, Citation1969; Andrejko and Cohen, Citation1984); (c) Likoaeng phytolith aridity index (Iph% = Chloridoid / Chloridoid + Panicoid), which provides a measure of xeric versus mesic grassland composition (Diester-Haas et al., Citation1973); (d) Likoaeng phytolith dicotyledon/Poaceae (D/P) ratio, a measure of woody (ligneous dicotyledon morphotypes) to grassy (Poaceae morphotypes) vegetation abundance and thus community openness (1 to 0, with 1 representing maximum woodland coverage and 0 none; Alexandre et al. Citation1997); (e) Likoaeng δ13C of sediment organic matter (δ13CSOM); (f) Likoaeng percentage proportions of C3 (versus C4) from δ13CSOM values (% of C3 plants = (δ13C −2 + 12.5)/−0.14; Wedin et al., Citation1995; after Bousman, Citation1991); (g) Likoaeng fish/mammal index (Σ NISP all fish / Σ [NISP all fish + NISP all mammals; 1 to 0, with 1 representing only fish and 0 only mammals]; Stewart and Mitchell, Citation2018a); (h) Likoaeng large ungulate/mammal index (Σ NISP all large ungulates / Σ [NISP all large ungulates + NISP all other mammals]; 1 to 0, with 1 representing only large ungulates and 0 only other mammals). The age/depth model employed is that found in Parker et al., Citation2011. The chronological boundaries of the late Holocene Neoglacial sensu lato can be seen in shaded blue, with the environmental and zooarchaeological inflection point that we identify at ∼2.6 ka highlighted.](/cms/asset/35daa8ec-daab-4843-ab95-1b32f5a7ae87/ttrs_a_2244923_f0008_oc.jpg)
Phase B witnessed a marked increase in cool-adapted C3 Pooid grass phytoliths at the expense of Panicoids and Chloridoids (both C4; Parker et al., Citation2011; (a)). Corroborated by the presence of phytoliths of Erica and other heathland taxa, as well as Protea and Leucosidea sericea charcoals, this apparent cooling signature is mirrored in the more negative δ13CSOM values (−21.0‰ to −18.9‰) relative to Phase A (−17.9‰ to −16.7‰) ((e)). C3 grasses in today’s Maloti Mountains, as discussed above, dominate at altitudes exceeding 2700–2100 m (depending on aspect). Given Likoaeng’s location deep in the Senqu Valley at an elevation of ∼1700 m, the Neoglacial therefore saw vegetation belts fall by at least 400 m. Altitudinal depressions of this magnitude are estimated to have been triggered by MAT reductions of ∼2.5°C relative to present-day (Parker et al., Citation2011). As elsewhere, accompanying this cooling is evidence of heightened humidity: frequencies of bulliform phytoliths – morphotypes argued to precipitate in wet conditions (Sangster and Parry, Citation1969; Andrejko and Cohen, Citation1984) – reach their highest values of the Likoaeng sequence ((b)), while the phytolith aridity index (Iph%; Diester-Haas et al., Citation1973) in Phase B hits its lowest (Parker et al., Citation2011; (c)).
Beyond contrasting with the occupational phases above and below, Phase B at Likoaeng evidences important ecological shifts within the Neoglacial itself. In keeping with the regional trend, the local environment responded to progressively colder and more humid conditions. While proportions of C3 Pooid grass phytoliths increase from 12% in Phase A to nearly 30% in lower Phase B, for example, after ∼2.6 kcal BP (upper Phase B) they rise sharply to 54% (Parker et al., Citation2011; (a)). At the same time there is a drop in woody (ligneous dicotyledon) morphotypes, with the index of tree cover density (dicotyledon/Poaceae or D/P ratio; Alexandre et al., Citation1997) falling from a sequence high of 0.20 in lower Phase B to 0.04 immediately after ∼2.6 kcal BP ((d)). This drastic reduction in trees and bushes, after what was a relatively well-wooded earlier Neoglacial, corresponds with the lowest phytolith aridity index of the sequence (Iph%) (Parker et al., Citation2011; (c)). Temperatures after ∼2.6 kcal BP were apparently sufficiently cold, therefore, for alpine grasses to proliferate in the Senqu Valley at the expense of woody taxa. The latter were either outcompeted or, very possibly, reduced under intense pressure from browsers. The δ13CSOM record bottoms out at ∼2.6 kcal BP with a value (−21.05‰; (e)) suggesting grass cover in the vicinity of Likoaeng reached >70% C3 ((f)). The subsequent uptick after this time likely stems from the resulting reductions in woody contributions to the C3 signal, although an eco-topographic lag effect cannot be ruled out (e.g. Alexander et al., Citation2018).
It is against this ecological backdrop that forager subsistence at Likoaeng underwent profound changes, evidenced in the site’s faunal assemblages. The shift, in the broadest terms, was one from an emphasis on mammalian game to one on fish. In Phase A, mammal bones comprise over 90% of the total faunal assemblage. This changed drastically during the Neoglacial apex (Phase B), when frequencies of mammal remains plunged relative to those of fish (Mitchell et al., Citation2011; (g)). From being less common in the earliest levels (Phase A and lowermost B), fish start overwhelming mammals by ∼2.8 kcal BP (Layer XIV; NISP = 808 to 118) before increasing exponentially ∼2.6 kcal BP to eclipse them by a ratio of 21:1 (Layer XIII; NISP = 19,138 to 414) (Stewart and Mitchell, Citation2018a:181; (g)). The species of fish targeted also changed at this time, from a relatively even mix of Orange River mudfish (Labeo capensis) and smallmouth yellowfish (Labeobarbus aeneus) to an almost singular focus on the former above Layer XIII (Mitchell et al., Citation2011:1234; Stewart and Mitchell, Citation2018a:184–185). Of the mammalian prey that declined, the most conspicuous are the large ungulates (i.e. size class 3 or above; Brain, Citation1981). Whereas the sequence’s five lowest archaeology-bearing levels (Layers XVII–XIII) yielded a total large ungulate NISP of 300, less than one-sixth this amount (NISP = 46) was recovered from the overlying 12 layers combined (Layers XII–I). Particularly striking is near-total disappearance of eland – the site’s only large bovid identifiable to species – after ∼2.6 kcal BP (Layer XII and above) (Plug et al., Citation2003; (h)).
Stewart and Mitchell (Citation2018a) have argued that expansions of alpine grasses were deleterious to large game populations, forcing longer range out-migrations in winter and constraining opportunities for seasonal (summertime) hunting. With encounter rates reduced, foragers were obliged to expand their diets to lower-ranked aquatic resources, which have been shown to offer hunter-gatherers viable intensification options (e.g. Broughton, Citation1994, Citation1997; Zangrando, Citation2009; Jerardino, Citation2010, Citation2012; Toso et al., Citation2021). Indeed, they may represent the only such option when primary productivity is low. Binford (Citation2001), in his ethnographic sample of 339 foraging societies worldwide, noted that those inhabiting areas with effective temperatures (ET) < 12.75°C subsist mostly on aquatic fauna or terrestrial game (rather than plantfoods), but that only the former can sustain enhanced population densities. Given that present-day ETs in the Maloti-Drakensberg do not exceed 14°C, and with much of the highlands <13°C (http://www.worldclim.com), the MAT drop of ∼2.5°C estimated for the Neoglacial would have pressed the region below this “terrestrial plant dependence threshold” (Binford, Citation2001). Faced with dwindling large game and unproductive alpine flora on which to fall back, afromontane foragers during the Neoglacial emphasised freshwater fish and other smaller food packets. High artefact densities, moreover, are suggestive of afromontane foragers converging on productive stretches of the deep valley bottoms along which resources had become concentrated, encouraging a degree of population packing in river corridors (Stewart and Mitchell, Citation2018a).
4.3. A new approach to painting?
Given foragers’ preoccupation with resource control and otherworldly negotiation, the magnitude of the ecological and dietary changes just described raises questions about concomitant ontological shifts. In the previous section, we saw that adjacent to Likoaeng is a highly suggestive rock art panel involving eland and fish, interpreted as expressing concerns with influencing rain and the resources dependent on it (Challis et al., Citation2008). The Likoaeng paintings remain undated, but the dietary shifts from eland to fish registered in the Neoglacial levels of the adjacent excavation might suggest the paintings relate to events at that time. A rare opportunity thus arises to gain a fuller understanding of cultural change by bridging the parietal and excavated archaeological records, but how to seize it?
Fortunately, a handful of direct 14C dates now exist that help illuminate the ontological response to Neoglacial conditions in the Maloti-Drakensberg. We argue that this both involved and necessitated an elaboration of pre-existing artistic practice.
Specifically, the recent work of Bonneau and colleagues (Citation2017a, Citation2017b, Citation2022; cf. Bonneau et al., Citation2011) has produced dates for shaded polychrome eland antelope, owing to the carbon black paint in their manufacture. The samples were taken from images in the Maclear region in the southern Maloti-Drakensberg (). As a whole, it appears that shaded polychrome eland entered the archaeological record during the Neoglacial, between ∼2.9–2.3 kcal BP (; ). As we have seen, Mazel (Citation2009:89) notes that shaded polychromes (eland as well as other animals, such as rhebok, bushpig and aardvark – and possibly fish: ) appear at the same point in the relative sequence across multiple research areas. Oxalate crusts underlying images produced dates of ∼4 kcal BP (Mazel and Watchman, Citation2003), which might in general terms corroborate a Neoglacial date. However, Mazel places shaded polychromes nearer the 2 kcal BP mark, in accordance with his ongoing hypothesis that they were produced in response to stress relating to the arrival of African agropastoralists (e.g. Mazel, Citation2009:103–7). While shaded polychromes may very well disappear because of contact (Mazel, Citation2009:104; cf. Challis and Sinclair-Thomson, Citation2022), the new direct dates place their inception between 500 and 1000 years before such arrivals (pace Mazel, Citation2022). Questions raised by Jolly (Citation2020) regarding the veracity of these dates have been forcibly rebuffed (Bonneau et al. in Jolly, Citation2020).
Figure 9. Sites in the Eastern Cape Drakensberg with directly dated shaded polychrome eland paintings. Carbon black was identified and isolated with pretreatment protocols to remove atmospheric carbon, and directly dated using AMS 14C, yielding results shown in . Images courtesy of Adelphine Bonneau and David Pearce.

Figure 10. Shaded polychrome fish at Likholong ha Piti on a tributary of the Senqu in southern Lesotho (see also Hobart and Smits, Citation2002; Hobart, Citation2003).Image and enhancement by authors.

Table 1. The four direct dates obtained for shaded polychrome eland in the Maloti-Drakensberg, after Bonneau et al. Citation2017a and supplementary material.
Rather than catalysed through culture contact, this new, embellished manner of depiction can best be understood, we suggest, as another cultural response to the Neoglacial, one visible in the socio-ontological domain. Viewed through the lens of San idiom, the placing of shaded polychromes in their chronological, environmental and excavated archaeological context allows us to make sense of this technique, and why it came about at this time. Especially during a phase when the environment was changing, the forager response would have been one of propriety and negotiation. If access to resources was achieved only on condition of correct behaviour, so respect toward the entities – rain, fish, eland – would have been enhanced. Conversely, the paintings so contextualised help complete the archaeological and palaeoenvironmental picture presented above.
5. DISCUSSION: ONTOLOGICAL ADAPTATIONS IN A VERTICAL WORLD
Afromontane foragers during the Neoglacial adjusted dynamically as their world in the high Maloti-Drakensberg changed around them. Responding to broader subcontinental climatic trends, local environments opened and closed windows onto resources whose successful capture demanded flexibility not only in diet, settlement and technology, but in social and ontological realms as well. Over the span of a millennium, societies went from year-round hunters of eland (pre-Neoglacial) to seasonal mass-harvesters of cyprinid fish (late Neoglacial), with an intervening phase when both resources were emphasised (early Neoglacial). This transition from terrestrial to aquatic resources placed premiums on logistical aptitude, requiring sharp seasonal timing and heightened spiritual precautions. Foraging, social and spiritual strategies were developed to capitalise on seasonal rainfall regimes, vegetation growth and concordant movement of prey animals. We hypothesise that the Neoglacial in the Maloti-Drakensberg witnessed a spiritual intensification married to that found in the faunal record. We also suggest that ideas about eland and fish, while originating before the Neoglacial in a pre-existing “grammar” (Barnard, Citation1988), coalesced in new ways that were both salient and enduring. Indeed, the impacts of these events on the Mountain San were profound, influencing highland foodways and artistic traditions into historic times.
5.1. Early Neoglacial (∼3–2.6 kcal BP)
In the early Neoglacial, high-altitude alpine vegetation began to move downslope with enhanced cooling (Parker et al., Citation2011). Like many mountain systems, surface area in the Maloti-Drakensberg decreases monotonically with elevation (Elsen and Tingley, Citation2015). Past depressions in vegetation belts would have resulted, therefore, in unpalatable alpine grasses covering larger proportions of the landscape than they do today. However, this expansion of C3 grasses would have also resulted in much more of the mountains experiencing the short “burst” of higher grazing potential in the growing season. Eland are known for taking advantage of tight seasonal windows by congregating in very large numbers (e.g. Hillman, Citation1987, Citation1988; Keeping et al., Citation2018). Thus, while overall mammal biomass on the landscape was likely lower, for a brief window in summer large terrestrial fauna would have been plentiful – particularly the locally migratory and mountain-loving eland antelope.
Mitchell et al. (Citation2011) hypothesise that Likoaeng’s immense quantities of (often very large) fish suggests that its inhabitants aggregated there to exploit seasonal fish runs. We concur, but maintain that this scenario more likely characterised the late Neoglacial and after. In the early Neoglacial, we believe that eland was the target resource that motivated such aggregations. Seasonal aggregations so focused would have included locals on the Senqu and, via regional social networks (Stewart et al., Citation2020; Mitchell and Stewart, CitationIn press), probably bands of friends and relatives from elsewhere. Indeed, eland herds – congregating at altitude from surrounding lowland zones to consume the tender new C3 grass shoots – would have exerted a heavy draw on nonlocal foragers. To maximise the narrow hunting window, foragers may well have anticipated it by several weeks or more, readying their hunting gear for the arrival of up-migrating eland (cf. Binford, Citation1978). Rivers rising with spring rains would have precipitated spawning runs of fish, presenting those assembled with ready sources of protein and fat (Mitchell et al., Citation2011). That early Neoglacial foragers at Likoaeng were fishing from late spring to mid-summer is suggested by the relatively even representation, as we have seen, of Orange River mudfish and smallmouth yellowfish in these levels (Plug et al., Citation2010).
This historical trajectory thus entailed evolving encounters with two very different prey types – eland and cyprinid fish – which, during the early Neoglacial, came to share one trait: both were warm-season migrants into the high mountain zone, the first rains triggering their up-valley ascents. The arrival of the rains, the subsequent running of the fish, and finally the arrival of the herds would have created conditions in which a number of actions – according to San idiom – were necessary. Because the high mountain pastures create a seasonal pulse of flora and fauna, so people’s subsistence and spiritual strategies pulsed to converge here. Specifically, the need to influence the rain and the game would have become critical, with ritual specialists probably working together as they do today when bands aggregate, as well as holding healing dances to dissipate tensions (Katz, Citation1982; Wiessner, Citation2002).
As people came together in the highland summer for the eland hunt, their behaviour in this sense echoed that of the eland themselves (Vinnicombe Citation1976; for large summer aggregations of eland see Melton and Snyman, Citation1989). Eland venture upwards in the San idiom because they are called by |Kaggen and fed by !Khwa – the rain. Both deities are involved in these aggregations, giving them supernatural power (both danger and raw potential). Because human society mimicked eland society in this way, we postulate that relations brokered between humans and non-humans were likely to have been that much more closely and well negotiated.
The response to the more temporally-focused eland aggregation would, in the San idiom, have required a mediation with the forces on the landscape including the rain which brought the new grass, and therefore the animals. People summon/tame and “cut” the rain’s animal so that the veld is regenerated and eland arrive. Never is this more important than in this time-pressed context. Concentrating efforts on influencing the rain (!Khwa) for the grass to shoot and the fish to run, in anticipation of the arrival of the eland in a narrowing window of opportunity, entailed a tightening of focus and a heightening of ritual action in an environmental ontology well suited to such adaptation. The spiritual links between eland, fish and rain would have become emphasised, the three things mutually reinforcing each other during this time of plenty. Once the game started arriving and the “glut” of highland hunting began, further ritual mediation was required to control the dangerous levels of supernatural potency – the superabundance and the social ills that can follow. Therefore, as well as wishing to make animals behave “nicely” in the hunt, there would have been a need to coerce people to behave nicely as well. Indeed, the former cannot be achieved without the latter (Challis and Skinner, Citation2021; Skinner and Challis, Citation2022).
Depictions of eland – already central to the forager canon of rock art – receive lavish attention in the early Neoglacial, as the dated record from Maclear attests. We hypothesise that the enhanced care afforded these images reflects these concerns and may mark the beginning of the hyper-attentive focus on eland for which the Mountain San are well-known (Vinnicombe Citation1976). While other species are rendered with the shaded polychrome technique (including rhebok, bushpig and fish), these are rare, whereas for eland this portrayal becomes the standard. Indeed, shaded polychrome rain’s animals, whether bovid or serpentine, may well relate to the same era. Fish traps, nets and the act of fishing are usually painted in monochrome, but probably on account of their linear format – being usually too thin to “carry” shading. As we have seen, moreover, they often occur in close proximity to and in the same panels as shaded polychrome eland. Further to the observation that rain’s animals, fish and eland are associated, we ask whether this was in fact the origin of equating rain with eland – did the eland become one of the rain’s animals at this time? The pre-existing grammar is flexible enough to accommodate major dietary transitions, but at the same time this transition may have precipitated precedents of Mountain San ontology.
5.2. Later Neoglacial (∼2.6–2 kcal BP)
After ∼2.6 kcal BP, rainfall increased and became more evenly distributed throughout the year, with greater amounts falling in winter as snow (Fitchett et al., Citation2016b; Herbert and Fitchett, Citation2022). Colder temperatures encouraged even more intensive downwards incursions of “sour” C3 alpine grasses at the expense of more palatable (“sweet”) C4 Themeda-dominated communities. Crucially, multiple archives also suggest that woody vegetation declined as well, placing strain on the browsing antelopes that had come to dominate the region’s ungulate communities after the mid-Holocene (Plug and Engela, Citation1992; Plug et al., Citation2003; Plug and Mitchell, Citation2008).
As we have seen, Likoaeng at this time registers a dramatic downturn in eland remains, a pattern at odds with that of fish, which continue to dominate most of the rest of the sequence. It might be tempting to relate this purely to changes in site function in an otherwise unaltered resource landscape. For example, during its depositional history Likoaeng experienced a major geomorphological transformation that also could have affected how or when people used it. Specifically, it switched from a rockshelter to an open-air site when accrued sediments filled and then buried the former (Layer XI; Mitchell et al., Citation2006:83). However, the shifts in ungulate and fish frequencies occurred prior to this – that is, when the site was still a shelter.
Rather than a change in site function, we interpret the crash in eland numbers as signalling a downturn in their abundance on the landscape. Today, the primary limiting factor on populations of wild antelope in uKhahlamba-Drakensberg parks, whether endemic or reintroduced, is food quality. While never high due to the region’s extremely leached soils, the nutritive quality of even palatable forage like Themeda triandra plunges during the cold-season (Mentis, Citation1978; Rowe-Rowe, Citation1983). Eland are highly unselective mixed-feeders that eat mostly grasses in spring and summer, but in winter are dependent on browse. For this reason, as we have seen, they disperse widely and migrate to low-lying, well-wooded valleys (Melton and Snyman, Citation1989). In the late Neoglacial, with higher annual rainfall, colder temperatures and longer-lasting snow leading to widespread alpine grasses with few woody plants atop intensely leached soils, highland environments were likely unsuitable for eland populations, even in summer. Obliged to migrate further for winter forage, their return migrations during the growing season evidently reached up less frequently into highland Lesotho.
As mentioned, by the late Holocene eland were apparently the only very large ungulate present in the highlands (Plug et al., Citation2003), a pattern at odds with sites below the Escarpment where other large game, though rarer than in the mid-Holocene, nevertheless persisted (Mazel, Citation1984; Opperman, Citation1987, Citation1996). For highland foragers, losing eland – their only large game species – would have thus been particularly serious. Perhaps unsurprisingly, then, after eland populations began eschewing the highlands the trend towards enhanced fishing continued. The representation of fish species also narrowed sharply to a taxon that spawns earlier in the year (Orange River mudfish). This suggests a concomitant change in the season of site use, from extended occupations spanning spring to mid-summer before ∼2.6 kcal BP to shorter, springtime-focussed ones after that time. Thus, the intensified focus on aquatic resources (and other small food parcels) went hand-in-hand with a change in foragers’ annual schedules to a different timetable: with no hunting glut to incentivize longer (spring-summer) occupation, late Neoglacial aggregations focused on the beginning of a short fish run, in spring.
The crash in eland numbers might otherwise be interpreted as a trigger for stress, and a seasonal abandonment. Yet we suspect something of the reverse in that the robust ontology of the San, shaped as it was over the millennia, allowed these afromontane foragers to pivot, as conditions altered, from terrestrial to subaquatic game. Images of fish traps, nets and spearfishing from boats occur across the southern Maloti-Drakensberg on both sides of the Escarpment. The southern Maloti-Drakensberg also exhibit the highest concentration of rain’s animal depictions (Dowson, Citation1994, Citation1995, Citation1998). It is more than coincidence that fishing and rain’s animals come together in this region. The change in climatic and faunal data must have had an ontological counterpart, and we hypothesise that this will be seen in a change in spiritual emphasis, from negotiating primarily with the entities of the uppermost tiers of the cosmos to those associated with the lower places.
5.3. A Cosmos Compressed
The high Maloti-Drakensberg – above the cloud and shrouded in mist, where elands are in droves like cattle, and where fish run up the rivers into the sky – are a manifestation of the San cosmos (Lewis-Williams and Challis, Citation2011). This conceptualisation would have been thrown into sharp relief in the early Neoglacial, when the vertical axis mundi became “compressed”. Descending vegetation belts brought eland from higher-altitude grazing grounds to within closer range of reconfigured, valley-centred human settlement systems and their hunting parties, but only for a short time. For local (and perhaps visiting) groups aggregated in highland valleys to take advantage of this glut of game, timing was everything. Successful hunting demanded presence and preparation, themselves dependent on similarly time-sensitive, aquatic resources: the rains arriving, the rivers rising and the fish running. The reliance on fish to sustain preparatory aggregations of hunters and their families thus brought these two mountain resources together both physically and, by extension, ontologically.
Specialist tamers of the game and of the rain (of elands and snakes) would have been expected to ensure the arrival of rains and associated resources. Equally crucial was the need to dissipate the inevitable tensions of the aggregated families, as well as deal with the superabundance of spiritual power present in the aggregated eland herds. San testimony suggests that all such tasks were undertaken in altered states of consciousness (e.g. in the sky or underwater), during which relations with those entities were brokered. Because of the anticipation, the tension and the need to broker “nice” relations with one another, with the animals and with the spirits, we have hypothesised a summer burst of ritual activity, visible in the inception of concentrated attention on eland paintings via the shaded polychrome technique. The exertion of influence over rain’s animals would have been the first priority to ensure precipitation and the rising of the rivers. Underwater cosmology reflected in rock art depicts not only the manifestations of !Khwa, but the specialists themselves influencing the movements of the fish (). Fish, in turn, enabled hunting parties to anticipate the arrival of the eland. In this way, the realms of the cosmos above and the cosmos below were compressed, creating an ontological fusion of fish and eland () – both of which came to be rain’s things. This, we suggest, helped ease the subsequent dietarily substitution of the latter with the former when eland populations ceased congregating in the highlands. It follows that this ontological flux will be reflected in fish-related concerns in paintings, direct dates for which are thus a major research priority.
Figure 11. From the Eastern Cape Drakensberg, the upper painting (a) depicts an underwater encounter between a !Khwa-ka-!gi:xa tamer-of-the-rain and two dark !Khwa-ka-xoro rain’s animals surrounded by fish. The rain’s animals are superimposed on two bichrome eland, not by chance, because the eland is the quintessence of supernatural potency and itself came to symbolise the rain. In (b) we see Patricia Vinnicombe’s re-drawing of (part of) a “fishing scene” on the Tsoelike, not far from Sehlabathebe. A.J. Goodwin (Citation1949) noted the “fishing scenes” at Kenegha Poort and Mpongweni on the south-eastern side of the Drakensberg Escarpment and in 1960, Vinnicombe, already familiar with the Drakensberg “scenes”, began finding images of fishing in high Maloti (Vinnicombe, Citation1960). The human figure standing amongst the fish she thought must have been unrelated. Thanks to her own (1976), Lewis-Williams’s (Citation1981:112) and others’ later research, what we now know of San cosmology allows us to see the human figure underwater, controlling the movements of the subaquatic game. Images from ARADA, Rock Art Research Institute.
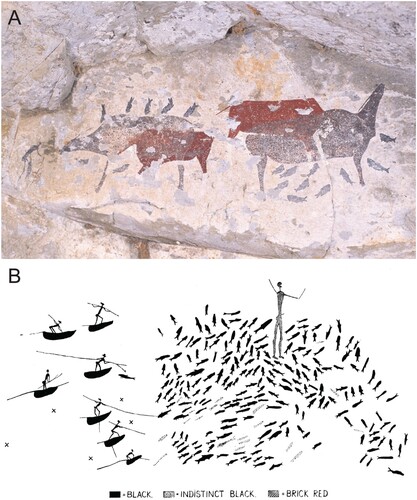
6. CONCLUSION
Few regions of the world boast richer visual and material archives of hunter-gatherer lifeways than the Maloti-Drakensberg Mountains. The region’s first professional archaeologists, Patricia Vinnicombe and Patrick Carter, understood both their individual importance and the power of coupling them. While still far too few, direct dates from specific paintings present a long-awaited opportunity to do this, and so begin constructing the three-ply cable braiding the region’s ethnography, rock art and excavated archaeology (cf. Ouzman and Wadley, Citation1997). In this paper, we have hypothesised several interlocking phenomena with repercussions for the last ∼3 kcal BP in the high Maloti-Drakensberg. Integrating cultural remains from rock walls and sedimentary deposits with New Animistic readings of critical San testimonies, we see an overall spiritual intensification during the Neoglacial that mirrors the evidence for dietary intensification (Stewart and Mitchell, Citation2018a). This process played out in two distinct phases divided by a noticeable inflection point ∼2.6 kcal BP.
In the early Neoglacial, the target of social aggregations in the highlands were eland populations who themselves were congregating there to exploit the short summer burst of shooting C3 grasses. By this time, large ungulates had become rare across the Maloti-Drakensberg except for eland, which came to prominence (Carter, Citation1978:187; Opperman, Citation1987:85; Plug and Mitchell, Citation2008:48). The earliest shaded polychromes were painted, displaying an enhanced degree of care and attention that reflects San idioms of correct behaviour that, we argue, relates to a summer burst of ritual behaviour necessary at such aggregations. Fishing also ramped up, with an extended spring-summer season geared towards feeding the groups amassed for the hunt. Triggering the in-migrations of both fish and eland was early seasonal rainfall, and ritual practitioners specialised in taming the rain worked towards the dual goal of enticing game and raising the rivers to bring fish. One consequence was the ontological fusion of rain, eland and fish, a conceptual blurring which facilitated the subsequent shift in emphasis from terrestrial to subaquatic resources when eland vacated the highlands in the later Neoglacial (after ∼2.6 kcal BP). Direct dates are needed to assess whether the onset of fish-related concerns in paintings mark this shift, but clear associations between paintings of fish – in some areas enhanced in shaded polychrome () – with rain animals ( and ) ties them unambiguously to seasonal rainfall and thus concerns about spawning (Challis et al., Citation2008).
Placing directly dated shaded polychromes (Bonneau et al., Citation2017a,Citation2017b, Citation2022) in their full archaeological context, our model seeks to account for when and why the eland became the Mountain San’s large game animal par excellence (Vinnicombe Citation1976). Indeed, those dates show that painting of said shaded polychromes did not cease with the Neoglacial, and there is clear evidence that eland remained ontologically central to Mountain San into historical times (Orpen, Citation1874). Eland feature in Qing’s creation story as god’s favourite animal “in droves like cattle” where he gives it special prominence; he further speaks of the eland and the rain in the same context of negotiation and influence. At present, however, we do not know how and why these ideas proliferated and endured over the past two millennia, nor what explains their variable expression across the region. Many questions remain. To what extent, for example, were ontological associations formed under one set of ecological circumstances still adaptive after conditions changed? Does the profusion of large, shaded polychrome eland along the Escarpment (e.g. (c,d)) stem from preservation bias or attempts to influence up-migrating eland? Do occasional shaded paintings in other parts of the subcontinent (Pager, Citation1973; Mazel, Citation2009:111) speak to similar concerns as shaded polychromes in the Maloti-Drakensberg? Answers to these and other questions must await further data, not least from Neoglacial-aged sites in highland Lesotho other than Likoaeng. That this open-air site survived on the banks of the upper Senqu, a river segment known for its violent floods and erosive flushing of highland sediments, is extraordinarily fortunate and something on which we have admittedly tried to capitalise.
There is a trope in anthropological archaeology that heightened stress results in heightened ritual (e.g. Johnson, Citation1982; Jochim, Citation1983; Alberti, Citation2014). Typically, however, such claims lack specificity as to which stressors may have been filtered through what socio-cultural mechanisms and thus, in the absence of ethnographic scaffolding (Conkey, Citation1985; Hayden, Citation1987; Hitchcock and Ebert, Citation1989; Turpin, Citation1990), are deficient in explanatory power. Rather than viewing it as a Pavlovian response, our model can and does include the particulars of when, why and in what forms heightened ritual occurred. The ontological shifts that we infer, moreover, speak less to a society in trouble than to one “doing particularly well” (cf. Johnson, Citation1982:406) That is to say, they are not a response to stress but an adaptation to change. This society’s success lies in the inherent mutability of the San’s pre-existing spiritual grammar, an attribute that lends their spiritual system marked durability (Guenther, Citation1999, Citation2020b).
Resilience returns us to the concept of environmental ontology. Ecologically-induced transformations of relationships between people, resources and the supernatural entities they embody, all rooted in landscape, are both spiritual and inherently adaptive. Though hunter-gatherer religious life cannot be reduced to single characteristics, one common feature is that it can serve to promote – and thus cannot be separated from – survival (e.g. Biesele, Citation1993; Whallon, Citation2011). To deny that hunter-gatherer religion provides means to an end is to suggest that indigenous epistemologies are not pragmatic – indeed relational ontologies “function” to serve the purposes of the hunter, as Willerslev (Citation2004:646) observes. In our case, supernatural re-negotiations eased the ecological changes and logistical challenges faced by upland communities via the creation of new ontological associations from a pre-existing grammar. The outcomes of these adaptations are both material and intangible, infusing key aspects of ethnographically-attested Mountain San ideology and its expression through a spectacular new mode of painting. Direct dates for shaded polychrome eland and all rock art imagery are critical to the long-pursued project of bridging the parietal and excavated archaeological records, but they are not enough. Employing the lens of environmental ontology affords archaeologists the inferential tools necessary for making synchronous archives mutually intelligible, rendering hunter-gatherer pasts – like shaded polychromes themselves – in fuller colour.
ACKNOWLEDGMENTS
We wish to thank Peter Mitchell and Adrian Parker for generously making the Likoaeng palaeoenvironmental and zooarchaeological data available. Donald Guy and the late Rethabile Mokhachane provided key insights into contemporary eland migration and aggregation behaviours in the Underberg-Sehlabathebe region. Three anonymous reviewers made useful comments that helped strengthen this paper, for which we are grateful. We thank the Kingdom of Lesotho’s Department of Culture for ongoing permitting and support. With deep appreciation we also acknowledge the seminal work of Patricia Vinnicombe and Patrick Carter, whose initial vision of an intertwined parietal and excavated archaeological record in the Maloti-Drakensberg we hope is finally beginning to be realised.
DISCLOSURE STATEMENT
No potential conflict of interest was reported by the authors.
REFERENCES
- Alberti, G. 2014. Modeling group size and scalar stress by logistic regression from an archaeological perspective. PLoS ONE 9 (3): e91510.
- Alexander, J., Chalmandrier, L., Lenoir, J., Burgess, T., Essl, F., Haider, S., Kueffer, C., McDougall, K., Milbau, A., Nuñez, M.A. and Pauchard, A. 2018. Lags in the response of mountain plant communities to climate change. Global Change Biology 24(2): 563–579.
- Alexandre, A., Meunier, J., Lezine, A., Vincens, A. & Schwartz, D. 1997. Phytoliths: indicators of grassland dynamics during the late Holocene in intertropical Africa. Palaeogeography, Palaeoclimatology, Palaeoecology 136: 213–229.
- Ambrose, D. 2006. Mammals: Including Annotated Species Checklist. House 9 Publications. Roma, National University of Lesotho.
- Andrejko, M. & Cohen, A. 1984. Scanning electron microscopy of silicophytoliths from the Okefenokiee swamp-marsh complex. In Cohen, A.D., Casagrande, D.J., Andrejko, M.J. & Best, G.R. (Eds.) The Okefenokiee Swamp: Its Natural History, Geology and Geochemistry. Los Alamos, Wetland Surveys. pp. 468–491.
- Arbousset, T. 1991. Missionary Excursion into the Blue Mountains. Ambrose, D. & Brutsch, A. (Eds.). Morija, Morija Archives.
- Arbousset, T. & Daumas, F. 1846. Narrative of an Exploratory Tour to the North-East of the Colony of the Cape of Good Hope. Cape Town, Robertson.
- Arthington, A., Rall, J. & Kennard, M. 1999. Specialist report: fish. Final report to the Lesotho Highlands Development Authority.
- Badenhorst, S., Mitchell, P., Arthur, C. & Capelli, C. 2019. Late Holocene fauna from Moshebi’s Shelter, a Later Stone Age site in Lesotho. Southern African Humanities 32 (1): 83–107.
- Barnard, A. 1988. Structure and fluidity in Khoisan religious ideas. Journal of Religion in Africa 18 (3): 216–236.
- Bawden, M. & Carroll, D. 1968. The Land Resources of Lesotho. London, United Kingdom Directorate of Overseas Surveys.
- Biesele, M. 1974. A note on the beliefs of modern Bushmen concerning the Tsodilo Hills. Newsletter of the SWA Scientific Society 15: 1–3.
- Biesele, M. 1993. Women Like Meat: The Folklore and Foraging Ideology of the Kalahari Ju/'hoan. Johannesburg, Witwatersrand University Press; Bloomington, Indiana University Press.
- Binford, L. 1978. Dimensional analysis of behavior and site structure: learning from an Eskimo hunting stand. American Antiquity 43: 330–361.
- Binford, L.R. 2001. Constructing Frames of Reference. Berkeley, University of California Press.
- Bird-David, N. 1999. “Animism” revisited: personhood, environment, and relational epistemology. Current anthropology 40 (S1): S67–S91.
- Bleek, D. 1933. Beliefs and customs of the |Xam Bushmen, part VI. Rain-Making. Bantu Studies 7 (4): 375–392.
- Bleek, D. 1935. Customs and beliefs of the/Xam Bushmen: part VII: Sorcerors [sic]. Bantu Studies 9: 1–47.
- Bleek, D. 1956. A Bushman Dictionary. New Haven, American Oriental Society.
- Bleek, W. & Lloyd, L. 1911. Specimens of Bushman Folklore. London, G. Allen & Company.
- Bonneau, A., Brock, F., Higham, T., Pearce, D. & Pollard, A. 2011. An improved pretreatment protocol for radiocarbon dating black pigments in San rock art. Radiocarbon 53 (3): 419–428.
- Bonneau, A., Pearce, D., Mitchell, P., Didier, L., Nic Eoin, L., Higham, T., Lamothe, M. & Arthur, C. 2022. Characterization and dating of San rock art in the Metolong catchment, Lesotho: a preliminary investigation of technological and stylistic changes. Quaternary International 611-612: 177–189.
- Bonneau, A., Pearce, D., Mitchell, P., Staff, R., Arthur, C., Mallen, L., Brock, F. & Higham, T. 2017a. The earliest directly dated rock paintings from southern Africa: new AMS radiocarbon dates. Antiquity 91 (356): 322–333.
- Bonneau, A., Staff, R., Higham, T., Brock, F., Pearce, D. & Mitchell, P. 2017b. Successfully dating rock art in southern Africa using improved sampling methods and new characterization and pretreatment protocols. Radiocarbon 59 (3): 659–677.
- Boshoff, A. & Kerley, G. 2013. Historical Incidence of the Larger Mammals in the Free State Province (South Africa) and Lesotho. Port Elizabeth, Centre for African Conservation Ecology; Nelson Mandela Metropolitan University.
- Boshoff, A. & Kerley, G. 2015. Lost herds of the Highveld: evidence from the written, historical record. African Journal of Wildlife Research 45 (3): 287–300.
- Boshoff, A., Landman, M. & Kerley, G. 2016. Filling the gaps on the maps: historical distribution patterns of some larger mammals in part of southern Africa. Transactions of the Royal Society of South Africa 71 (1): 23–87.
- Bousman, C.B. 1991. Holocene paleoecology and Later Stone Age hunter-gatherer adaptations in the South African interior plateau. Ph.D. Dissertation, Southern Methodist University.
- Brain, C. 1981. The Hunters of the Hunted? An Introduction to African Cave Taphonomy. Chicago, University of Chicago Press.
- Broughton, J. 1994. Late Holocene resource intensification in the Sacramento Valley, California: the vertebrate evidence. Journal of Archaeological Science 21: 501–514.
- Broughton, J. 1997. Widening diet breadth, declining foraging efficiency, and prehistoric harvest pressure: ichthyofaunal evidence from the Emeryville Shellmound, California. Antiquity 71: 845–862.
- Cambray, J. 1985. Observations on spawning of Labeo capensis and Clarias gariepinus in the regulated lower Orange River, South Africa. South African Journal of Science 81 (6): 318–321.
- Carbutt, C. 2019. The Drakensberg Mountain Centre: a necessary revision of southern Africa's high-elevation centre of plant endemism. South African Journal of Botany 124: 508–529.
- Carbutt, C. & Edwards, T. 2001. Cape elements on high-altitude corridors and edaphic islands: historical aspects and preliminary phytogeography. Systematics and Geography of Plants 71: 1033–1061.
- Carbutt, C. & Edwards, T. 2004. The flora of the Drakensberg alpine centre. Edinburgh Journal of Botany 60 (03): 581–607.
- Carbutt, C. & Edwards, T. 2006. The endemic and near-endemic angiosperms of the Drakensberg Alpine Centre. South African Journal of Botany 72 (1): 105–132.
- Carbutt, C. & Edwards, T. 2015. Reconciling ecological and phytogeographical spatial boundaries to clarify the limits of the montane and alpine regions of sub-Sahelian Africa. South African Journal of Botany 98: 64–75.
- Carter, P.L. 1970. Late Stone Age exploitation patterns in southern Natal. The South African Archaeological Bulletin 25 (98): 55–58.
- Carter, P. 1978. The Prehistory of Eastern Lesotho. Doctoral Thesis, University of Cambridge.
- Challis, S. 2019. High and mighty: a San expression of excess potency control in the high-altitude hunting grounds of southern Africa. Time and Mind 12 (3): 169–185.
- Challis, S., Hollmann, J. & McGranaghan, M. 2013. ‘Rain snakes’ from the Senqu River: new light on Qing's commentary on San rock art from Sehonghong, Lesotho. Azania: Archaeological Research in Africa 48 (3): 331–354.
- Challis, S., Mitchell, P. & Orton, J. 2008. Fishing in the rain: control of rain-making and aquatic resources at a previously undescribed rock art site in highland Lesotho. Journal of African Archaeology 6 (2): 203–218.
- Challis, S. & Sinclair-Thomson, B. 2022. The impact of contact and colonization on indigenous worldviews, rock art, and the history of southern Africa: “the disconnect”. Current Anthropology 63 (S25): S91–S127.
- Challis, S. & Skinner, A., 2021. Art and influence, presence and navigation in southern African forager landscapes. Religions 12(12): 1099.
- Clarke, M. 1888. Unexplored Basuto Land. Proceedings of the Royal Geographical Society and Monthly Record of Geography 10: 519–30.
- Conkey, M. 1985. Ritual communication, social elaboration, and the variable trajectories of Paleolithic material culture. In Price, T.D. & Brown, J.A. (Eds.) Prehistoric Hunter-Gatherers: The Emergence of Complexity. Orlando, Academic Press. pp. 299–323.
- Deacon, J. 1988. The power of a place in understanding Southern San rock engravings. World Archaeology 20 (1): 129–40.
- Deacon, J. 1997. My heart stands in the hill': rock engravings in the Northern Cape. Kronos: Journal of Cape History 24 (1): 18–29.
- Descola, P. 2012. Beyond nature and culture: the traffic of souls. HAU: Journal of Ethnographic Theory 2 (1): 473–500.
- Diester-Haas, L., Schrader, H.J. & Thiede, J. 1973. Sedimentological and paleoclimatological investigations of two pelagic ooze cores off Cape Barbas, North-West AfricaMeteor Forschungsergebnisse: Reihe C, Geologie und Geophysik 16: 19–66.
- Dowson, T. 1994. Reading art, writing history: rock art and social change in southern Africa. World Archaeology 25 (3): 332–345.
- Dowson, T. 1995. Hunter-gatherers, traders and slaves: the ‘Mfecane’ impact on Bushmen, their ritual and their art. In Hamilton, C. (Ed.) The Mfecane Aftermath: Reconstructive Debates in Southern African History. Johannesburg, Witwatersrand University Press. pp. 51–70.
- Dowson, T. 1998. Rain in Bushman belief, politics and history: the rock art of rain-making in the south-eastern mountains, southern Africa. In Chippindale, C. & Taçon, P.S.C. (Eds.) The Archaeology of Rock-Art. Cambridge, Cambridge University Press. pp. 73–89.
- Du Plessis, S. 1969. The Past and Present Geographical Distribution of the Perissodactyla and Artiodactyla in Southern Africa. Masters Thesis, University of Pretoria.
- Ellery, W., Scholes, R. & Scholes, M. 1995. The distribution of sweetveld and sourveld in South Africa's grassland biome in relation to environmental factors. African Journal of Range & Forage Science 12 (1): 38–45.
- Elsen, P. & Tingley, M. 2015. Global mountain topography and the fate of montane species under climate change. Nature Climate Change 5: 772–776.
- Fitchett, J., Grab, S., Bamford, M. & Mackay, A. 2016a. A multi-disciplinary review of late Quaternary palaeoclimates and environments for Lesotho. South African Journal of Science 112: 24–32.
- Fitchett, J., Grab, S., Bamford, M. & Mackay, A. 2016b. A multi-proxy analysis of late Quaternary palaeoenvironments, Sekhokong Range, eastern Lesotho. Journal of Quaternary Science 31: 788–798.
- Fitchett, J., Mackay, A., Grab, S. & Bamford, M. 2017. Holocene climatic variability indicated by a multi-proxy record from southern Africa’s highest wetland. The Holocene 27: 638–650.
- Gaigher, I., Hamman, K. & Thorne, S. 1980. The distribution, conservation status and factors affecting the survival of indigenous freshwater fishes in the Cape Province. Koedoe 23 (1): 57–88.
- Gerber, R., Smit, N. & Wagenaar, J. 2012. Age, growth rate and size at sexual maturity of Labeobarbus aeneus (Teleostei: Cyprinidae) in the Middle Vaal River, South Africa. African Journal of Aquatic Science 37 (1): 49–58.
- Germond, R. 1967. Chronicles of Basutoland. Morija, Morija Sesuto Book Depot.
- Goodwin, A. 1949. A fishing scene from East Griqualand. The South African Archaeological Bulletin 4 (14): 51–53.
- Grab, S. 1994. Thufur in the Mohlesi Valley, Lesotho, southern Africa. Permafrost and Periglacial Processes 5 (2): 111–118.
- Grab, S. 1997. Thermal regime for a thufa apex and its adjoining depression, Mashai Valley, Lesotho. Permafrost and Periglacial Processes 8 (4): 437–445.
- Grab, S. 2000. Stone-banked lobes and environmental implications, high Drakensberg, southern Africa. Permafrost and Periglacial Processes 11 (3): 177–187.
- Grab, S. & Deschamps, C. 2004. Geomorphological and geoecological controls and processes following gully development in alpine mires, Lesotho. Arctic, Antarctic, and Alpine Research 36 (1): 49–58.
- Grab, S. & Mills, S. 2011. Quaternary slope processes and morphologies in the upper Sehonghong Valley, eastern Lesotho. Proceedings of the Geologists' Association 122: 179–186.
- Grab, S. & Nash, D. 2022. “But what silence! No more gazelles … ”: occurrence and extinction of fauna in Lesotho, southern Africa, since the late Pleistocene. Quaternary International 611–612: 91–105.
- Grab, S. & Nüsser, M. 2001. Towards an integrated research approach for the Drakensberg and Lesotho mountain environments: a case study from the Sani plateau region. South African Geographical Journal 83: 64–68.
- Guenther, M. 1999. Tricksters and Trancers: Bushman Religion and Society. Bloomington, Indiana University Press.
- Guenther, M. 2015. Therefore their parts resemble humans, for they feel that they are people. Hunter Gatherer Research 1 (3): 277–315.
- Guenther, M. 2020a. Human-Animal Relationships in San and Hunter-Gatherer Cosmology, Volume I: Therianthropes and Transformation. Cham, Palgrave Macmillan.
- Guenther, M. 2020b. Human-Animal Relationships in San and Hunter-Gatherer Cosmology, Volume II: Imagining and Experiencing Ontological Mutability. Cham, Palgrave Macmillan.
- Halbmayer, E. 2012. Debating animism, perspectivism and the construction of ontologies. Indiana 29: 9–23.
- Hallowell, I. 1960. Ojibwa ontology, behavior, and world view. In Diamond, S. (Ed.) Culture in History: Essays in Honor of Paul Radin. New York, Columbia University Press. pp. 19–52.
- Hanvey, P. & Marker, M. 1992. Present-day periglacial microforms in the Lesotho Highlands: implications for present and past climatic conditions. Permafrost and Periglacial Processes 3: 353–361.
- Harris, G., Thirgood, S., Hopcraft, J., Cromsigt, J. & Berger, J. 2009. Global decline in aggregated migrations of large terrestrial mammals. Endangered Species Research 7 (1): 55–76.
- Harvey, G. 2010. Animism rather than shamanism: new approaches to what shamans do (for other animals). In Schmidt, B. & Huskinson, L. (Eds.) Spirit Possession and Trance: New Interdisciplinary Perspectives. London, Bloomsbury. pp. 16–34.
- Hayden, B. 1987. Alliances and ritual ecstasy: human responses to resource stress. Journal for the Scientific Study of Religion 26: 81–91.
- Herbert, A. & Fitchett, J. 2022. Synthesising the pollen records for the Drakensberg-Maloti through quantitative modelling. Quaternary International 611-612: 77–86.
- Hillman, J. 1987. Group size and association patterns of the common eland (Tragelaphus oryx). Journal of Zoology 213 (4): 641–663.
- Hillman, J. 1988. Home range and movement of the common eland (Taurotragus oryx Pallas 1766) in Kenya. African Journal of Ecology 26 (2): 135–148.
- Hitchcock, R. & Ebert, J. 1989. Modeling Kalahari hunter-gatherer subsistence and settlement systems: implications for development policy and land use planning in Botswana. Anthropos 84: 47–62.
- Hobart, J. 2003. An old fashioned approach to a modern hobby: fishing in the Lesotho Highlands. In Mitchell, P., Haour, A. & Hobart, J. (Eds.) Researching Africa’s Past. New Contributions from British Archaeologists. Oxford University School of Archaeology Monograph 57. Oxford, Oxford University Press. pp. 44–53.
- Hobart, J. & Smits, L. 2002. Fishing in the Lesotho highlands. Antiquity 76 (291): 39–40.
- Hoff, A. 1997. The water snake of the Khoekhoen and/Xam. The South African Archaeological Bulletin 52 (165): 21–37.
- Hoff, A. 1998. The water bull of the/Xam. The South African Archaeological Bulletin 53 (168): 109–124.
- Hogg, A., Heaton, T., Hua, Q., Palmer, J., Turney, C., Southon, J., Bayliss, A., Blackwell, P., Boswijk, G., Bronk Ramsey, C., Pearson, C., Petchey, F., Reimer, P., Reimer, R. & Wacker, L. 2020. SHCal20 Southern Hemisphere Calibration, 0–55,000 Years cal BP. Radiocarbon 62 (4): 759–778.
- Hollmann, J. 2007. The ‘cutting edge’ of rock art: motifs and other markings on Driekuil Hill, North West Province, South Africa. Southern African Humanities 19 (1): 123–151.
- Hollmann, J. 2022. Customs and Beliefs of the |Xam. 2nd ed, New York and Johannesburg, NYU Press and Wits University Press.
- Holmgren, K., Karl ́en, W., Lauritzen, S., Lee-Thorp, J., Partridge, T., Piketh, S., Rebinski, P., Stevenson, C., Svanered, O. & Tyson, P. 1999. A 3000-year high-resolution stalagmitebased record of palaeoclimate for northeastern South Africa. The Holocene 9: 295–309.
- Holmgren, K., Lee-Thorp, J., Cooper, G., Lundblad, K., Partridge, T., Scott, L., Sithaldeen, R., Talma, A. & Tyson, P. 2003. Persistent millennial-scale climatic variability over the past 25,000 years in southern Africa. Quaternary Science Reviews 22: 2311–2326.
- Hornborg, A. 2006. Visiting the six worlds: Shamanistic journeys in Canadian Mi’kmaq cosmology. Journal of American Folklore 119 (473): 312–336.
- How, M. 1962. The Mountain Bushmen of Basutoland. Pretoria, van Schaik.
- Hydén, L. 2002. The influence on summer rainfall in the Lesotho Lowlands from Indian Ocean SSTs. Hydrology Research 33 (4): 305–318.
- Jacot Guillarmod, A. 1971. Flora of Lesotho (Basutoland). Lehre, Verlag Von Cramer.
- Jerardino, A. 1995. Late Holocene Neoglacial episodes in southern South America and southern Africa: a comparison. The Holocene 5 (3): 361–368.
- Jerardino, A. 2010. Large shell middens in Lamberts Bay, South Africa: a case of hunter-gatherer resource intensification. Journal of Archaeological Science 37: 2291–2302.
- Jerardino, A. 2012. Large shell middens and hunter-gatherer resource intensification along the west coast of South Africa: the Elands Bay case study. The Journal of Island and Coastal Archaeology 7: 76–101.
- Jochim, M. 1983. Palaeolithic cave art in ecological perspective. In Bailey, G. (Ed.) Hunter-Gatherer Economy in Prehistory: A European Perspective. Cambridge, Cambridge University Press. pp. 212–219.
- Johnson, G. 1982. Organizational structure and scalar stress. In Renfrew, C., Rowlands, M. & Segraves-Whallon, B. (Eds.) Theory and Explanation in Archaeology. Cambridge, MA, Academic Press. pp. 389–421.
- Jolly, P. 2020. New methods for direct dating of southern African rock paintings–Testing the methods. The South African Archaeological Bulletin 75 (213): 161–164.
- Jubb, R. 1966. Notes on the popular angling yellowfishes and distributions outside their native range. Piscator 68: 120–125.
- Katz, R. 1982. Boiling Energy: Community Healing among the Kalahari Kung. Cambridge, MA, Harvard University Press.
- Keeping, D., Keitsile, A. & Burger, J. 2018. Eland in Botswana's Kalahari: now the largest free-ranging population in Africa? Gnusletter 35: 21–26.
- Killick, D. 1963. An account of the plant ecology of the Cathedral Peak area of the Natal Drakensberg. Memoirs of the Botanical Survey of South Africa 34: 1–178.
- Killick, D. 1978. The afro-alpine region. In Werger, M.J.A. (Ed.) Biogeography and Ecology of Southern Africa. The Hague, Junk. pp. 515–560.
- Killick, D. 1990. A Field Guide to the Flora of the Natal Drakensberg. Johannesburg, Jonathan Ball.
- Kinahan, J. 2017. The Dancing Kudu: women’s initiation in the Namib Desert during the second millennium AD. Antiquity 91(358): 1043–1057.
- Knight, J. & Grab, S. 2014. Lightning as a geomorphic agent on mountain summits: evidence from southern Africa. Geomorphology 204: 61–70.
- Lee-Thorp, J., Holmgren, K., Lauritzen, S., Linge, H., Moberg, A., Partridge, T., Stevenson, C. & Tyson, P. 2001. Rapid climate shifts in the southern African interior throughout the mid to late Holocene. Geophysical Research Letters 28: 4507–4510.
- Lewis-Williams, D. 1981. Believing and Seeing: Symbolic Meanings in Southern San Rock Paintings. London, Academic Press.
- Lewis-Williams, D. 1998. Quanto?: the issue of ‘many meanings’ in southern African San rock art research. The South African Archaeological Bulletin 53 (168): 86–97.
- Lewis-Williams, D. 2000. Stories That Float from Afar: Ancestral Folklore of the San of Southern Africa. Cape Town, David Philip.
- Lewis-Williams, D. 2013. Qwanciqutshaa, a 19th-century southern San mythological being: who is he and what does his name mean? South African Archaeological Bulletin 68 (197): 79–85.
- Lewis-Williams, D. & Biesele, M. 1978. Eland hunting rituals among northern and southern San groups: striking similarities. Africa 48 (2): 117–134.
- Lewis-Williams, D., Blundell, G., Challis, W. & Hampson, J. 2000. Threads of light: re-examining a motif in southern African San rock art. The South African Archaeological Bulletin 55 (172): 123–136.
- Lewis-Williams, D. & Challis, S. 2011. Deciphering Ancient Minds: The Mystery of San Bushman Rock Art. London, Thames & Hudson.
- Lewis-Williams, D. & Dowson, T. 1989. Images of Power: Understanding Bushman Rock Art. Johannesburg, Southern Book Publishers.
- Lewis-Williams, D. & Pearce, D. 2004. Southern African San rock painting as social intervention: a study of rain-control images. African Archaeological Review 21 (4): 199–228.
- Lewis, C. 2005. Late glacial and Holocene palaeoclimatology of the Drakensberg of the Eastern Cape, South Africa. Quaternary International 129: 33–48.
- Lodder, J., Hill, T.R. & Finch, J.M. 2018. A 5000-yr record of Afromontane vegetation dynamics from the Drakensberg Escarpment, South Africa. Quaternary International 470A: 119–129.
- Mallen, L. 2005. Linking sex, species and a supernatural snake at Lab X rock art site. South African Archaeological Society Goodwin Series 9: 3–10.
- Marker, M. 1994. Sedimentary sequences at Sani Top, Lesotho highlands, southern Africa. The Holocene 4: 406–412.
- Marshall, L. 1969. The medicine dance of the !Kung Bushmen. Africa 39 (4): 347–381.
- Marshall, L. 1976. The!Kung of Nyae Nyae. Cambridge, MA, Harvard University Press.
- Marshall, L. 1999. Nyae Nyae! Kung Beliefs and Rites. Cambridge, MA, Peabody Museum Press.
- Mayet, N., Knight, J. & Grab, S.W. 2016. Spatial and temporal patterns of lightning strikes in the eastern Lesotho Highlands, southern Africa. South African Geographical Journal 98 (2): 321–336.
- Mazel, A. 1984. Diamond 1 and Clarke's Shelter: report on excavations in the northern Drakensberg, Natal, South Africa. Annals of the Natal Museum 26 (1): 25–70.
- Mazel, A. 2009. Unsettled times: shaded polychrome paintings and hunter-gatherer history in the southeastern mountains of southern Africa. Southern African Humanities 21 (1): 85–115.
- Mazel, A. 2022. Mountain living: the Holocene people of the uKhahlamba-Drakensberg, South Africa. Quaternary International 611: 190–210.
- Mazel, A. & Watchman, A. 2003. Dating rock paintings in the uKhahlamba-Drakensberg and the Biggarsberg, KwaZulu-Natal, South Africa. Southern African Humanities 15 (1): 59–73.
- McGranaghan, M. 2012. Foragers on the Frontiers: The |Xam Bushmen of the Northern Cape, South Africa, in the Nineteenth Century. Doctoral Thesis, University of Oxford.
- McGranaghan, M. 2014a. ‘He who is a devourer of things’: monstrosity and the construction of difference in |Xam Bushman oral literature. Folklore 125 (1): 1–21.
- McGranaghan, M. 2014b. ‘Different people’ coming together: representations of alterity in |Xam Bushman (San) narrative. Critical Arts 28 (4): 670–688.
- McGranaghan, M. 2016. The death of the agama lizard: the historical significances of a multi-authored rock-art site in the Northern Cape (South Africa). Cambridge Archaeological Journal 26 (1): 157–179.
- McGranaghan, M. & Challis, S. 2016. Reconfiguring hunting magic: Southern Bushman (San) perspectives on taming and their implications for understanding rock art. Cambridge Archaeological Journal 26 (4): 579–599.
- McGranaghan, M., Challis, S. & Lewis-Williams, D. 2013. Joseph Millerd Orpen's’ A Glimpse into the Mythology of the Maluti Bushmen': a contextual introduction and republished text. Southern African Humanities 25: 137–166.
- Melton, D. & Snyman, A. 1989. The influence of farmland on eland in the Natal Drakensburg. South African Journal of Wildlife Research 19 (1): 26–30.
- Mentis, M. 1978. Population limitation in grey rhebuck and oribi in the Natal Drakensberg. Lammergeyer 26: 19–28.
- Mentis, M. & Huntley, B. 1982. Description of the Grassland Biome Project. South African National Scientific Programmes Report No. 62. Pretoria, CSIR.
- Mguni, S. 2009. Natural and supernatural convergences: trees in southern African rock art. Current Anthropology 50 (1): 139–148.
- Mitchell, P. 2002. The Archaeology of Southern Africa. Cambridge, Cambridge University Press.
- Mitchell, P. 2009. Gathering together a history of the people of the Eland: towards an archaeology of Maloti-Drakensberg hunter-gatherers. In Mitchell, P.J. & Smith, B.W. (Eds.) The Eland’s People: New Perspectives in the Rock Art of the Maloti-Drakensberg Bushmen. Johannesburg, Wits University Press. pp. 99–139.
- Mitchell, P., Plug, I. & Bailey, G. 2006. Spatial patterning and site occupation at Likoaeng, an open-air hunter-gatherer campsite in the Lesotho Highlands, southern Africa. Archeological Papers of the American Anthropological Association 16 (1): 81–94.
- Mitchell, P., Plug, I., Bailey, G., Charles, R., Esterhuysen, A., Thorp, J., Parker, A. & Woodborne, S. 2011. Beyond the drip-line: a high-resolution open-air Holocene hunter-gatherer sequence from highland Lesotho. Antiquity 85 (330): 1225–1242.
- Mitchell, P. & Stewart, B. In press. Enveloping oneself in others: semiotic, spatial, and temporal dimensions of ostrich eggshell bead use in southern Africa. In Nowell, A. & Collins, B. (Eds.) Culturing the Body: Prehistoric Perspectives on Identity and Sociality. New York, Berghahn.
- Morris, C., Taintoi, N. & Boleme, S. 1993. Classification of the eastern alpine vegetation of Lesotho. African Journal of Range & Forage Science 10 (1): 47–53.
- Mucina, L. & Rutherford, M. 2006. The vegetation of South Africa, Lesotho and Swaziland. Pretoria, South African National Biodiversity Institute.
- Mulder, P.F.S. 1973. Aspects of the ecology of Labeo capensis and Labeo umbratus in the Vaal River. Zoologica Africana 8 (1): 15–24.
- Mulder, N. & Grab, S.W. 2009. Contemporary spatio-temporal patterns of snow cover over the Drakensberg. South African Journal of Science 105: 228–233.
- Mullen, A. 2018. Re-investigating significantly differentiated figures in the rock art of the South-eastern mountains. Masters Dissertation, University of the Witwatersrand.
- Nash, D.J. & Meadows, M.E. 2012. Africa. In Metcalf, S.E. & Nash, D. (Eds.) Quaternary Environmental Change in the Tropics. Chichester, John Wiley & Sons. pp. 79–150.
- Neumann, F.H., Botha, G.A. & Scott, L. 2014. 18,000 years of grassland evolution in the summer rainfall region of South Africa: evidence from Mahwaqa Mountain, KwaZulu-Natal. Vegetation History and Archaeobotany 23: 665–681.
- O’Connor, T.G. & Bredenkamp, G.J. 1997. Grassland. In Cowling, R.M., Richardson, D.M. & Pierce, S.M. (Eds.) Vegetation of Southern Africa. Cambridge, Cambridge University Press. pp. 215–257.
- Opperman, H. 1987. The Later Stone Age of the Drakensberg Range and its Foothills. Oxford, British Archaeological Reports.
- Opperman, H. 1996. Excavation of a Later Stone Age deposit in Strathalan Cave A, Maclear District, northeastern Cape, South Africa. In Pwiti, G. & Soper, R. (Eds.) Aspects of African Archaeology. Harare, University of Zimbabwe Press. pp. 335–342.
- Orpen, J. 1874. A glimpse into the mythology of the Maluti Bushmen. The Cape Monthly Magazine 9: 1–13.
- Ouzman, S. & Wadley, L. 1997. A history in paint and stone from Rose Cottage Cave, South Africa. Antiquity 71 (272): 386–404.
- Pager, H. 1971. Ndedema: A Documentation of the Rock Paintings of the Ndedema Gorge. Graz, Akademische Druck- u. Verlagsanstalt.
- Pager, H. 1973. Shaded rock-paintings in the Republic of South Africa, Lesotho, Rhodesia and Botswana. The South African Archaeological Bulletin 28 (109–110): 39–46.
- Parker, A.G., Lee-Thorp, J.A. & Mitchell, P.J. 2011. Late Holocene Neoglacial conditions from the Lesotho Highlands, southern Africa: phytolith and stable carbon isotope evidence from the archaeological site of Likoaeng. Proceedings of the Geologists' Association 122: 201–211.
- Plug, I. 1997. Late Pleistocene and Holocene hunter–gatherers in the eastern highlands of South Africa and Lesotho: a faunal interpretation. Journal of Archaeological Science 24 (8): 715–727.
- Plug, I. 2003. Appendix D: Report on Pitsaneng fauna. In Hobart, J. (Ed.) Forager-Farmer Relations in South-eastern Southern Africa: A Critical Reassessment. Doctoral thesis. University of Oxford. pp. 425–428.
- Plug, I. 2008. Fish bone sizes as estimators of standard lengths of three southern African freshwater species with application to archaeological samples: a preliminary investigation. South African Journal of Science 104 (1): 71–75.
- Plug, I. 2017. Middle and Later Stone Age hunters and their prey in southern Africa. In Albarella, U., Rizzetto, M., Russ, H., Vickers, K. & Viner-Daniels, S. (Eds.) The Oxford Handbook of Zooarchaeology. Oxford, Oxford University Press. pp. 385–395.
- Plug, I. & Badenhorst, S. 2001. The distribution of macromammals in Southern Africa over the past 30 000 Years as reflected in animal remains from archaeological sites. Transvaal Museum Monograph No 12. Pretoria, Transvaal Museum.
- Plug, I. & Engela, R. 1992. The macrofaunal remains from recent excavations at Rose Cottage Cave, Orange Free State. The South African Archaeological Bulletin 47: 16–25.
- Plug, I. & Mitchell, P. 2008. Fishing in the Lesotho highlands: 26,000 years of fish exploitation, with special reference to Sehonghong Shelter. Journal of African Archaeology 6 (1): 33–55.
- Plug, I., Mitchell, P. & Bailey, G. 2003. Animal remains from Likoaeng, an open-air river site, and its place in the post-classic Wilton of Lesotho and eastern Free State, South Africa. South African Journal of Science 99 (3): 143–152.
- Plug, I., Mitchell, P. & Bailey, G. 2010. Late Holocene fishing strategies in southern Africa as seen from Likoaeng, highland Lesotho. Journal of Archaeological Science 37 (12): 3111–3123.
- Quinlan, T. & Morris, C.D. 1994. Implications of changes to the transhumance system for conservation of the mountain catchments in eastern Lesotho. African Journal of Range & Forage Science 11 (3): 76–81.
- Rakotsoane, F. 1996. Religion of the ancient Basotho with special reference to “water snake”. Masters Thesis, University of Cape Town.
- Rakotsoane, F. 2008. Traces of snake worship in Basotho culture. Journal for the Scientific Study of Religion 21 (1): 53–70.
- Rosen, D.Z., Lewis, C.A. & Illgner, P.M. 1999. Palaeoclimatic and archaeological implications of organic-rich sediments at Tiffindell Ski Resort, near Rhodes, Eastern Cape Province, South Africa. Transactions of the Royal Society of South Africa 54 (2): 311–321.
- Rowe-Rowe, D. 1983. Habitat preferences of five Drakensberg antelopes. South African Journal of Wildlife Research 13 (1): 1–8.
- Rowe-Rowe, D. & Scotcher, J. 1986. Ecological carrying capacity of the Natal Drakensberg for wild ungulates. South African Journal of Wildlife Research 16 (1): 12–16.
- Russell, T. 2000. The Application of the Harris Matrix to San Rock Art at Main Caves North, Kwazulu-Natal. The South African Archaeological Bulletin 55 (171): 60–70.
- Sangster, A. & Parry, D. 1969. Some factors in relation to bulliform cell silicification in the grass leaf. Annals of Botany 33: 315–323.
- Schmidt, S. 1979. The rain bull of the South African Bushmen. African Studies 38 (2): 201–224.
- Schrijvershof, J. 2015. Biology and ecology of fishes of the Senqu River, Lesotho. Masters Thesis, North-West University (South Africa).
- Schulze, R. 1979. Hydrology and Water Resources of the Drakensberg. Pietermaritzburg, Natal Town and Regional Planning Commission.
- Schwabe, C. 1995. Alpine mires of the eastern highlands of Lesotho. In Cowan, G.I. (Ed.) Wetlands of South Africa. Pretoria, Department of Environmental Affairs and Tourism. pp. 33–40.
- Scott, L., Holmgren, K., Talma, A., Woodborne, S. & Vogel, J. 2003. Age interpretation of the Wonderkrater spring sediments and vegetation change in the savanna biome, Limpopo province, South Africa. South African Journal of Science 99: 484–488.
- Sene, K., Jones, D., Meigh, J. & Farquharson, F. 1998. Rainfall and flow variations in the Lesotho Highlands. International Journal of Climatology 18 (3): 329–345.
- Shortt-Smith, K. 1963. Basutoland. In Harrison, A., Shortt-Smith, K., Yates, J., Jubb, R., Rushby, G. & Flamwell, C. (Eds.) Fresh-Water Fish and Fishing in Africa. Johannesburg, Nelson and Sons. pp. 55–72.
- Skelton, P. 2001. The Freshwater Fishes of Southern Africa. Cape Town, Struik.
- Skinner, A. 2017. The Changer of Ways: Rock Art and Frontier Ideologies on the Strandberg, Northern Cape, South Africa. Masters Thesis, University of the Witwatersrand.
- Skinner, A. 2021. Politics of identity in Maloti-Drakensberg rock art research. In Oxford Research Encyclopedia of Anthropology. doi:10.1093/acrefore/9780190854584.013.528
- Skinner, A. 2022a. A survey of social ecologies in the freshwater biomes of highland Lesotho and the adjacent Eastern Cape, South Africa. Quaternary International 611-612: 220–229.
- Skinner, A. 2022b. ‘Things of the outside teach me’: identity transfer and contextual transformation as expressions of persistent, syncretic cosmology in traditional spiritual and medicinal practice in the south-central Maloti-Drakensberg, southern Africa. Azania: Archaeological Research in Africa 57 (1): 121–146.
- Skinner, A. & Challis, S. 2022. Fluidities of personhood in the idioms of the Maloti-Drakensberg, past and present, and their use in incorporating contextual ethnographies in southern African rock art research. Time and Mind 15 (2): 101–141.
- Skinner, J. & Chimimba, C. 2005. The Mammals of the Southern African Subregion, 3rd edition. Cape Town, Cambridge University Press.
- Snyman, H.A., Ingram, L.J. & Kirkman, K.P. 2013. Themeda triandra: a keystone grass species. African Journal of Range & Forage Science 30 (3): 99–125.
- Staples, R. & Hudson, W.K. 1938. An Ecological Survey of the Mountain Area of Basutoland. London, Crown Agents for the Colonies.
- Stewart, B.A. In press. Space, Food and Magic: The Coastal Hunter-Herders of Dunefield Midden (South Africa). Los Angeles, Cotsen Institute of Archaeology Press.
- Stewart, B.A. & Mitchell, P. 2018a. Beyond the shadow of a desert: aquatic resource intensification on the roof of southern Africa. In Lemke, A. (Ed.) Foraging in the Past: Archaeological Studies of Hunter-Gatherer Diversity. Boulder, University Press of Colorado. pp. 159–208.
- Stewart, B.A. & Mitchell, P. 2018b. Late Quaternary palaeoclimates and human-environment dynamics of the Maloti-Drakensberg region, southern Africa. Quaternary Science Reviews 196: 1–20.
- Stewart, B.A., Zhao, Y., Mitchell, P., Dewar, I., Gleason, J. & Blum, J. 2020. Ostrich eggshell bead strontium isotopes reveal persistent macroscale social networking across late Quaternary southern Africa. Proceedings of the National Academy of Sciences 117: 6453–6462.
- Swallow, B., Brokken, R., Motsamai, M., Sopeng, L. & Storey, G. 1987. Livestock Development and Range Utilization in Lesotho. Institute of Southern African Studies Research Report 18. Maseru, Lesotho Ministry of Agriculture.
- Tainton, N. 1981. Veld and Pasture Management in South Africa. Pietermaritzburg, Shuter & Shooter.
- Talma, A.S. & Vogel, J. 1992. Late Quaternary palaeotemperatures derived from a speleothem from Cango Caves, Cape Province, South Africa. Quaternary Research 37: 203–213.
- Tanner, A. 1979. Bringing Home Animals. London, Hurst.
- Tómasson, T., Cambray, J. & Jackson, P. 1984. Reproductive biology of four large riverine fishes (Cyprinidae) in a man-made lake, Orange River, South Africa. Hydrobiologia 112: 179–195.
- Toso, A., Hallingstad, E., McGrath, K., Fossile, T., Conlan, C., Ferreira, J., da Rocha Bandeira, D., Giannini, P.C.F., Gilson, S.P., de Melo Reis Bueno, L. & Bastos, M.Q.R. 2021. Fishing intensification as response to Late Holocene socio-ecological instability in southeastern South America. Scientific Reports 11 (1): 23506.
- Turpin, S. 1990. Rock art and hunter-gatherer archaeology: a case study from SW Texas and northern Mexico. Journal of Field Archaeology 17 (3): 263–281.
- van Wyk, A. & Smith, G.F. 2001. Regions of Floristic Endemism in Southern Africa: A Review with Emphasis on Succulents. Pretoria, Umdaus Press.
- van Zinderen Bakker, E. & Werger, M. 1974. Environment, vegetation and phytogeography of the high-altitude bogs of Lesotho. Plant Ecology 29: 37–49.
- Vinnicombe, P. 1960. The recording of rock paintings in the upper reaches of the Umkomaas, Umzimkulu and Umzimvubu rivers. South African Journal of Science 56 (1): 11–14.
- Vinnicombe, P. 1972. Myth, Motive, and Selection in Southern African Rock Art. Africa 42 (3): 192–204.
- Vinnicombe, P. 1976. People of the Eland: Rock Paintings of the Drakensberg Bushmen as a Reflection of their Life and Thought. Pietermaritzburg, University of Natal Press.
- Viveiros de Castro, E. 2004. Exchanging perspectives: the transformation of objects into subjects in Amerindian ontologies. Common Knowledge 10 (3): 463–484.
- Vogel, J.C. 1983. Isotopic evidence for the past climates and vegetation of southern Africa. Bothalia 14 (3/4): 391–394.
- Wallis, R. 2009. Re-enchanting rock art landscapes: animic ontologies, nonhuman agency and rhizomic personhood. Time and Mind 2 (1): 47–69.
- Wedin, D.A., Tieszen, L.L., Dewey, B. & Pastor, J. 1995. Carbon isotope dynamics during grass decomposition and soil organic matter formation. Ecology 76 (5): 1383–1392.
- Whallon, R. 2011. An introduction to information and its role in hunter-gatherer bands. In Hitchcock, R., Whallon, R. & Lovis, W. (Eds.) Information and Its Role in Hunter-Gatherer Bands. Los Angeles, Cotsen Institute of Archaeology Press. pp. 1–28.
- Wiessner, P. 2002. Hunting, healing, and hxaro exchange: a long-term perspective on !Kung (Ju/’hoansi) large-game hunting. Evolution and Human Behavior 23: 407–436.
- Willerslev, R. 2004. Not animal, not not-animal: hunting, imitation and empathetic knowledge among the Siberian Yukaghirs. Journal of the Royal Anthropological Institute 10: 629–652.
- Witelson, D. 2022. Rock art and archaeology? The problem of ‘integration’ in southern African Later Stone Age research. In Hampson, J., Challis, S. & Goldhahn, J. (Eds.) Powerful Pictures: Rock Art Research Histories around the World. Oxford, Archaeopress. pp. 105–115.
- Wylie, A. 1989. Archaeological cables and tacking: the implications of practice for Bernstein's ‘Options beyond objectivism and relativism’. Philosophy of the Social Sciences 19 (1): 1–18.
- Zangrando, A. 2009. Is fishing intensification a direct route to hunter-gatherer complexity? A case study from the Beagle Channel region (Tierra del Fuego, southern South America). World Archaeology 41: 589–608.