Abstract
Soil respiration in forest plantations can be greatly affected by management practices. Irrigation is necessary for high productivity of poplar plantations in semi-arid northwest China. Moreover, plowing is essential for improving soil quality and reducing evaporation. In the present study, the influences of irrigation and plowing on soil carbon dioxide (CO2) efflux were investigated in poplar plantations in 2007 and 2008. The experiments included three stand age classes receiving three treatments: control, irrigation, and plowing. Mean soil respiration in irrigation treatment stands was 5.47, 4.86, and 4.43 µmol m−2 s−1 in 3-, 8-, and 15-year-old stands, respectively, during the growing season. In contrast, mean soil respiration in control stands was 3.71, 3.83, and 3.98 µmol m−2 s−1 in 3-, 8-, and 15-year-old stands, respectively. During the entire observation period, mean soil respiration in plowing treatment stands increased by 36.2% compared with that in the control stands. Mean soil respiration in irrigation treatment stands was significantly higher than that in the control stands; this was mainly because fine root growth and decomposer activities were greatly depressed by soil drought, since natural precipitation could not meet their water demands. The results also suggest that plowing management can greatly increase soil CO2 emission by modifying soil structure. After plowing, soil bulk density decreased and soil aeration was greatly improved, leading to greater rates of oxidation and mineralization.
Introduction
Soil respiration (SR) is one of the main components of ecosystem respiration (Goulden et al. Citation1996). Small changes in SR may strongly affect soil carbon sequestration (Raich and Schlesinger Citation1992). It is estimated that a global warming of 0.03°C per year will enhance soil carbon dioxide (CO2) efflux, producing a net release of an additional 60 Pg C from the soil to the atmosphere between 1990 and 2050 (Xu and Qi Citation2001). A potential positive feedback between increasing temperature and enhanced SR is now widely accepted (Smith et al. Citation1997; Grace and Rayment Citation1999). Therefore, it is important to obtain accurate estimates of SR and understand the effects of environmental controls on the underlying processes for the global carbon budget.
Soil is the biggest carbon pool of the continental biosphere (Schimel Citation1995) and requires particular attention, especially regarding short-rotation plantation, because it is a major component of durable carbon sequestration, with the aboveground biomass being frequently removed and transformed into wood products within short-time spans. The impact of afforestation on the global carbon budget was addressed in the Kyoto Protocol 1997 on climate change, with particular emphasis on the role of intensively managed forests to function as sinks for atmospheric carbon (Murray et al. Citation2000). Moreover, soil carbon sequestration by forest plantations is considered an effective method for offsetting the emission of CO2 into the atmosphere (Raich and Tufekcioglu Citation2000; Lal Citation2005). Thus, reliable estimates of the soil carbon dynamics of forest plantations are required to represent forest–atmosphere interactions and mitigate climate change.
Soil moisture is one of the main factors controlling SR (Raich and Schlesinger Citation1992; Davidson et al. Citation1998). Low or high soil water content may depress SR either by drought stress to microbial and root activities or by limited oxygen availability due to reduced air diffusion (Rey et al. Citation2002). In general, in intensively managed forests, cultivation practices can influence soil moisture and temperature to a large extent, consequently affecting soil CO2 efflux. Therefore, there is considerable interest to determine the effects of forest management on soil carbon balance and storage (Davidson et al. Citation1998). Such an understanding will be helpful for improving soil carbon sequestration in forest plantations by using suitable management practices.
However, most studies focus on the influences of management practices on biomass productivity (Laureysens et al. Citation2004; Swamy et al. Citation2006), and little research has been done to investigate their effects on soil CO2 emission in Populus plantations (O’Neill et al. Citation2002). Populus species, which are characterized by rapid growth and easy adaptability, are cultivated worldwide and play an increasingly important role as a source of carbon sequestration (Bradshaw et al. Citation2000). Cultural management in poplar plantations usually includes irrigation, rotation selection, and plowing. Because the high productivity of poplar plantations depends heavily on water availability (Puri and Thompson Citation2003; Monclus et al. Citation2006) and because low soil moisture can limit soil CO2 evolution, particularly in arid or semi-arid ecosystems (Orchard and Cook Citation1983; Conant et al. Citation2004), we hypothesized that irrigation practices greatly affect SR in poplar plantations. Moreover, plowing is useful for weed control and improving soil quality in Populus plantations. However, plowing may loosen the soil structure and increase the exposure of soil organic matter. Previous research suggests that soil disturbance can greatly stimulate soil CO2 efflux (Fortin et al. Citation1996; Curtin et al. Citation2000; Jabro et al. Citation2008). Therefore, it is necessary to obtain sufficient information on the influences of irrigation and plowing on SR if we are to improve the soil carbon storage of poplar plantations. In the present study, we measured SR, fine root biomass, and soil properties in order to (1) compare the variations in SR between control and irrigation (plowing) treatment, (2) determine the effects of irrigation on SR, and (3) determine the effects of plowing on SR.
Material and Methods
Study site description
The study area is located at the Pingyuan Forestry Farm, Yili River Valley, northwest China (81°09′E, 43°45′N, 660 m above sea level). The climate is continental semi-arid with a mean annual temperature of 8.4°C, ranging from −12.2°C in January to 22.7°C in July. The mean annual precipitation is approximately 203.8 mm. The frost-free period lasts from May to September. Annual sunshine is up to 2800 h. The groundwater is typically 2.0–2.3 m below the soil surface. The soil type is Sierozem originating from a parent material of loess. Soil texture is typical sandy loam with an organic matter content of 12.6 to 25.2 g kg−1, total nitrogen of 1.1 to 2.3 g kg−1, and pH 8–8.5.
Pingyuan Forestry Farm is an experimental center for afforestation in northwest China and covers roughly 345 ha. Most of the poplar plantations are established on lands formerly growing wheat (Triticum aestivum) and have undergone intensive managements, such as irrigation, fertilization (40 and 120 kg N ha−1 in years one and five, respectively), and weed control. Water resources originating from the Yili River are abundant in the farm area, which provides an advantage for irrigation and plantation development. Because of weed control management, herbaceous vegetation such as Carex liparocarpos, Bromus japonicus, and Achnatherum splendens is very sparse.
Experimental design
A clone of Populus deltoids cv. was selected for this study because it is a typical clone used for afforestation in local farms. Three stand age classes representative of three developmental stages including 3-, 8-, and 15-year-old stands were chosen in this study. We considered each age class separately in a randomized block design. Each age class had three blocks. In order to investigate the effects of irrigation and plowing on SR, we established the following experiments separately: control versus irrigation and control versus plowing in each block. Each treatment was randomly replicated three times. The treatment plots were 20 m × 30 m. To minimize sources of error, the plots were set up on sites with similar soil types, landscape positions, parent materials, and prior land-use history. The sample trees were selected away from the borders of each stand to minimize any edge effects. Forest stands were established with 3.0 m × 4.0 m spacing. The stand characteristics are shown in .
Table 1 Characteristics of poplar plantation stands (July 2007). Data are means ± standard errors†
Through the growth seasons of 2007 and 2008, irrigation plots received an average of 350 m3 ha−1 water monthly. Irrigation treatment lasted five days, and water was supplied at intervals during each day. Trees were irrigated five times from May to September. In contrast, the control plots received no additional water other than precipitation (the precipitation was 93.8 and 84.0 mm for the growth seasons of 2007 and 2008, respectively, based on the Dynamet meteorological station in the forestry farm). To avoid the influences of excessive soil water and heterogeneity of soil water distribution on soil CO2 emission, measurements were taken two days after irrigation. In the plowing treatment, soil was plowed to a soil depth of 20 cm using a rototiller in May 2007. In order to obtain steady SR rates, measurements were taken two days after plowing treatment. If there was a rainfall event, the measurements in the controls were obtained a week after rainfall.
Measurement of soil respiration and soil conditions
Soil respiration was measured using a Li-6400 portable CO2 infrared gas analyzer equipped with a Li-6400-09 chamber (Li-Cor Inc, Lincoln, NE). The polyvinyl chloride (PVC) soil collars were inserted 2–3 cm below the surface into the soil after litter was removed; these collars were left in the study field throughout the measurement period. To account for within-plot variability, 6–8 soil collars were randomly established: three of them situated at the base of a tree and the others between two planting rows. Measurements were taken 48 h after the collars were installed. Soil respiration was measured monthly from May to September of 2007 and 2008. Within each month, all of the soil collars were examined from 10:00 a.m. to 14:00 p.m. To minimize measurement errors, measurements were taken only on sunny days without high winds.
Soil temperature was monitored simultaneously with SR measurement using a Constantan thermocouple penetration probe (Li-6000-09 TC; Li-Cor Inc.) inserted to soil depths of 5, 10, and 15 cm by the collars. Six soil samples (0–20 and 20–40 cm deep) near the collars were collected in each plot and oven-dried at 105°C for over 24 h to a constant mass to determine the soil water content.
Soil and root sampling
During the growth seasons of 2007 and 2008, the fine root (<2 mm in diameter) biomass of three depths (0–10, 10–20, and 20–40 cm) was estimated monthly on SR measurement days. A soil auger (8 cm in diameter) with a sharpened edge was pushed into the soil to collect fine roots in the vicinity of the soil collars. Eighteen fine root samples from three depths were obtained monthly from each plot. In July 2007, coarse root (2–10 mm in diameter) biomass was randomly obtained from the three soil pits (1 m3) in each control plot. All roots in the samples were washed and separated by hand, oven-dried at 80°C for over 24 h, and weighed using an electric balance (±0.01 g). To estimate bulk density, six soil samples were randomly collected from each plot (0–10, 10–20, and 20–40 cm deep) using a steel core (100 cm3 in volume). In July 2007, six composite soil samples each consisting of nine random core samples from two depths (0–10 cm and 10–20 cm) were collected from each plot. Soil samples were passed through a 2-mm sieve and subsequently analyzed in the laboratory to determine the physical and chemical properties of the soil. Soil carbon content was analyzed using the Walkley–Black wet oxidation method, and soil nitrogen was measured using the Kjeldahl method.
Statistical analysis
All data were analyzed using SPSS 15.0 (SPSS, Chicago, IL). An analysis of covariance (ANCOVA) test with soil temperature and water content as covariates was used to compare SR among age classes. A paired t-test was conducted to determine significant differences in SR between control and irrigation (plowing) treatment. Analysis of variance (ANOVA) was used to examine the effects of stand age and irrigation (plowing) as well as their interactions. An exponential model was used to fit the relationship between SR and soil temperature, and a linear model was used to represent the relationship between SR and soil moisture. Multiple regression analyses were carried out to evaluate the relationship between soil respiration and relevant factors. Significant differences for all statistical tests were evaluated at α = 0.05. The models used are as follows:
Results
Soil respiration in irrigation treatment and control
Throughout the growing season, the SR of three age classes in irrigation treatment exhibited a distinct seasonal pattern: increasing from May to June, peaking in July, and then rapidly decreasing in September (). With all measurements pooled together, the mean SR rates under irrigation were 5.47, 4.86, and 4.43 µmol m−2 s−1 in 3-, 8-, and 15-year-old stands, respectively. Soil respiration exhibited a wide seasonal range: 3.90–7.64, 3.56–7.46, and 2.60–7.03 µmol m−2 s−1 in 3-, 8-, and 15-year-old stands, respectively. The results of ANCOVA revealed significant differences in SR among the three stand age classes (P < 0.01). During the observation period, the largest SR occurred in the youngest plantation and SR was 19.0% lower in 15-year-old stands.
Figure 1 Comparison of soil respiration (SR) among control, plowing treatment, and irrigation treatment (a, b, and c represent 3-, 8-, and 15-year-old stands, respectively) during the growing seasons of 2007 and 2008. Vertical bars indicate standard error. n = 22.
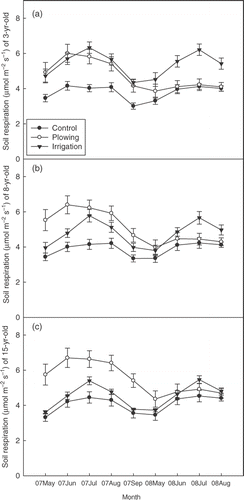
In the control, the SR of the 3- and 8-year-old stands gradually increased from May to June, then tended to level off, and decreased in September (). Meanwhile, the SR in the 15-year-old stands exhibited a clear seasonal pattern peaking in July. In general, soil CO2 efflux in the controls was greatly depressed, especially in July, compared with that in the irrigated stands. With all measurements pooled together, mean efflux rates in the controls during the growing season were 3.71, 3.83, and 3.98 µmol m−2 s−1 in 3-, 8-, and 15-year-old stands, respectively. Without irrigation, the greatest SR occurred in the oldest stands. There were no significant differences in SR between irrigation treatment and controls in the 15-year-old stands (P > 0.05). However, in 3-year-old stands, the SR in the irrigation treatment was significantly higher than that in the controls (P < 0.01).
With measurements in irrigated and control stands pooled together, the results of ANOVA indicate that the main effects of irrigation and stand age on SR are all significant (P < 0.01) (). In addition, the results revealed a significant interaction (P < 0.01) between irrigation and stand age. Furthermore, the SR of 3-year-old stands was more sensitive to soil water availability.
Table 2 Results of analysis of variance (ANOVA) on the effects of stand age, irrigation (plowing), and their interactions on soil respiration
Soil respiration in the plowed stands
Soil respiration of three age classes in the plowing treatment stands increased sharply in the beginning of the growing season of 2007, peaked in June, and remained relatively high in July and August (). In comparison, the seasonal pattern of SR of the plowing treatment greatly differed from those of the control in 2007 (). However, the difference in SR between the plowing treatment and control gradually decreased in the subsequent sampling months. In June 2008, there was no significant difference between the two treatments ().
With all measurements pooled together, the mean efflux rates in the plowing treatment stands were 4.82, 5.26, and 5.62 µmol m−2 s−1 in 3-, 8- and 15-year-old stands, respectively. The mean SR of the plowing treatment stands was significantly higher than that of the controls (P < 0.01). The results of ANOVA indicated that plowing had a significant effect on SR (). In addition, it was noteworthy that the largest difference in SR between the two treatments occurred in June 2007 shortly after the plowing. The SR in the plowing treatment stands increased by 54.4% in June 2007 compared with that in the control.
Soil temperature and water content
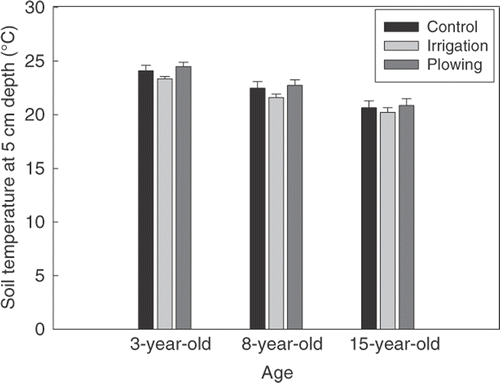
During the growing season, soil temperature at 5 cm depth (T) of the three treatments exhibited a similar seasonal pattern, with the maximum occurring in July. With all measurements pooled together, the average values of T were 22.37°C, 21.02°C, and 22.63°C in the control, irrigation treatment, and plowing treatment stands, respectively (). There was no significant difference in soil temperature between control and treatment stands, although the T of plowing treatment stands was slightly higher than those of the other groups.
Figure 2 Soil temperature at 5 cm depth for control, irrigation treatment, and plowing treatment of three stand age classes, averaged over the experimental period. Vertical bars indicate standard error. n = 158.
At the seasonal scale, the SR in the irrigation treatment stands closely followed the changes in soil temperature. The exponential model indicated that T could explain roughly 73–79% of variation in SR (3-year-olds: SR = 2.126e 0.041 t , n = 160, R 2 = 0.732, P < 0.01; 8-year-olds: SR = 1.594e 0.043 t , n = 156, R 2 = 0.775, P < 0.01; 15-year-olds: SR = 1.231e 0.062 t , n = 154, R 2 = 0.791, P < 0.01). However, in the control stands, higher soil temperature in the summer months was related to the lower SR, and T explained only 36–45% of the variation in SR.
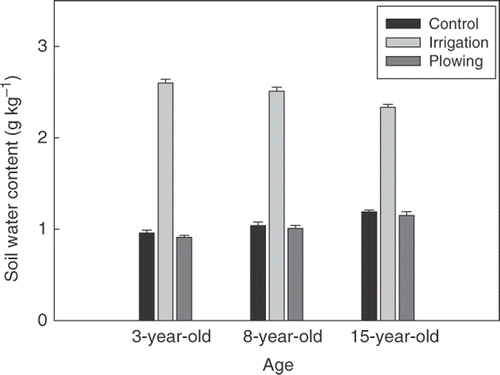
Because of monthly irrigation, soil water contents (W) in the irrigation treatment stands were 26.3%, 25.1%, and 23.3% in 3-, 8-, and 15-year-old stands, respectively (). Soil moisture in the irrigation treatment fluctuated relatively little during the growing season. Soil water content in the control stands usually remained below 15.8% during the growing season, with average values of 9.5%, 10.4%, and 11.9% in 3-, 8-, and 15-year-old stands, respectively. Soil water content in the plowing treatment was slightly lower than that in the control. Soil water content reached the minimum in July.
Figure 3 Soil water content at 0–20 cm depth for control, irrigation treatment, and plowing treatment of three stand age classes, averaged over the experimental period. Vertical bars indicate standard error. n = 156.
In the controls, SR tended to increase with increasing soil moisture; the linear model showed that soil water content was closely related to SR (3-year-olds: SR = 2.670 + 0.109W, n = 162, R 2 = 0.663, P < 0.01; 8-year-olds: SR = 2.599 + 0.131W, n = 148, R 2 = 0.627, P < 0.01; 15-year-olds: SR = 2.875 + 0.119W, n = 154, R 2 = 0.682, P < 0.01). In the irrigation treatment stands, correlation analysis revealed that soil water content was weakly related to SR (P > 0.05).
Soil conditions and fine root biomass
At 0–20 cm depth, soil carbon contents averaged by three age classes were 14.82, 14.75, and 14.56 g kg−1 in the control, irrigation treatment, and plowing treatment stands, respectively. Mean soil nitrogen concentrations at 0–20 cm depth were 1.33, 1.32, and 1.28 g kg−1 in the control, irrigation treatment, and plowing treatment stands, respectively. There were no significant differences in soil carbon contents between control and treatment stands (P > 0.05). In comparison, there was an obvious decrease in soil nitrogen content in plowing treatment. Soil pH exhibited a narrow range from 8.29 to 8.36 across all treatments ().
Soil bulk density increased with soil depth (), with mean values of 1.22, 1.23, and 1.18 g cm−3 at 0–10 cm deep in the control, irrigation treatment, and plowing treatment stands, respectively. Soil bulk density greatly decreased after plowing, and there were significant differences between control and plowed stands (P < 0.05).
Table 3 Soil bulk density and fine root biomass in control and treatment stands of poplar plantations in 2007 and 2008†
Fine root growth exhibited a distinct seasonal pattern. In the irrigation treatment stands, fine root biomass at 0–10 cm depth averaged over the growing season was 144.2, 126.1, and 113.3 g m−2 in 3-, 8-, and 15-year-old stands, respectively (). In the control stands, fine root biomass at 0–10 cm depth was 57.2, 71.4, and 75.6 g m−2 in 3-, 8-, and 15-year-old stands, respectively (). There was a significant difference in fine root biomass between the control and irrigated stands (P < 0.01).
To investigate the relationship between SR and relevant factors, multiple stepwise regression was performed between SR and T, W, fine root biomass at 0–10 cm depth (FR), soil carbon and nitrogen contents, pH, and soil bulk density among the three treatments. The results suggested that T, W, and FR were important controlling factors, roughly explaining 67–85% of variation in SR (). In the irrigation treatment stands, T could explain roughly 76.6% of the variation in SR averaged by three age classes and fine root biomass could explain an additional 8.6% (). In the control, the linear model indicated that W could explain roughly 65.7% of the variation in SR averaged by the three age classes; the correlation coefficient of the regression model was improved by 5.9% when fine root biomass was included (). However, the regression model by T, W, and FR was not significant for predicting SR in the plowing treatment stands.
Table 4 Regression relationship between soil respiration (SR) and relevant factors by pooling data from all sampling months in 2007 and 2008†
Discussion
Influence of irrigation on soil respiration
In general, the availability of water is closely related to high biomass production in poplar plantations (Puri and Thompson Citation2003; Monclus et al. Citation2006). Furthermore, soil moisture is essential for root development and microbial activity, and consequently has a great impact on SR (Beukes Citation1984; Davidson et al. Citation1998). However, the effect of soil moisture on SR has not yet been quantified at the forest stand level, and researchers have produced widely different results among climatic regions (Raich and Schlesinger Citation1992; Davidson et al. Citation1998; Maier and Kress Citation2000; Gaumont-Guay et al. Citation2006a).
Conant et al. (Citation2004) reported that SR tended to increase with increasing soil moisture; and SR under wetter conditions was 2.5 times greater than that under drier conditions. Gaumont-Guay et al. (Citation2006b) observed that daily SR increased rapidly from 3.6 to 9.0 µmol m−2 s−1 following heavy rainfall. In another study, Gaumont-Guay et al. (Citation2006a) also found that SR was greatly depressed when soil moisture exceeded 25–30% in aspen stands. In the present study, it was observed that SR in the control stands remarkably declined (), especially in July when soil moisture reached a minimum. During the study period, SR in the control stands decreased by 22.0% compared with that in the irrigated stands. This implies that soil moisture is an obvious limiting factor of SR. Because root and decomposer activities could be greatly inhibited by insufficient soil water availability, SR in the control stands was greatly depressed.
Soil water deficiency may inhibit root growth, increase mortality of fine roots, and promote suberization of root tips (Beukes Citation1984), which can eventually depress root respiration (Burton et al. Citation1998). Previous studies indicate that fine root growth and production decline with decrease in soil moisture in poplar clones (Dickman et al. Citation1996; Pregitzer and Friend Citation1996). Dickman et al. (Citation1996) observed that the fine root proliferation of poplar clones apparently occurred in soil profiles with high soil water availability. Coyle and Coleman (Citation2005) found that the root biomass of two cottonwood clones responded positively to irrigation treatment. In the present study, fine root biomass clearly decreased with decrease in soil water. In the controls, fine root biomass at 0–10 cm depth decreased by 46.3% () compared with that in the irrigated stands; this was partly responsible for the decrease in root respiration in the control stands. Previous studies show that root biomass tends to be positively correlated with root respiration (Ewel and Cropper Citation1987; Burton et al. Citation1998; Saiz et al. Citation2006). Burton et al. (Citation1998) reported that root respiration associated with fine root growth decreased because of soil water shortage. Saiz et al. (Citation2006) found that the root respiration of spruce forests decreased with stand age, which was concordant with the decrease in fine root biomass. Ewel and Cropper (Citation1987) found that soil CO2 efflux in a 29-year-old pine plantation was 35% higher than that in 9-year-old pines, which was mainly due to the higher root biomass in the older plantation. In the present study, the predictive capacity of regression models could be improved by 6–9% when fine root biomass was included, which suggested that fine root also had influence on SR besides soil water content.
In addition, the strong inhibition of SR observed in the control stands might be partly due to a reduction in microbial activity under soil drought conditions (Davidson et al. Citation1998; Borken et al. Citation2002; Gaumont-Guay et al. 2006a). Orchard and Cook (Citation1983) state that lower soil moisture can depress SR by limiting microbial contact with available substrate and the activity of microorganisms. Gaumont-Guay et al. (Citation2006a) found that the increase in SR was associated with enhanced microbial decomposition due to the replenishment of soil water in aspen stands. In the present study, mean soil water content in the control stands was 10.6%, which approximately corresponded to the permanent wilting point in the top soil layer. Therefore, microbial respiration might be greatly depressed because of soil water limitation.
Furthermore, the SR of the three age classes responded differently to soil water shortage. Unlike the SR of other age stands, the SR in 3-year-old stands significantly decreased by 32.2% in the control, compared with that in the irrigation treatment. This was mainly because the root systems in 3-year-old stands were relatively small and confined to the shallow soil layer (Coyle and Coleman Citation2005). When soil water availability decreased, they had difficulties in absorbing soil water from deeper soil layers. However, in the older plantations, root systems could reach deep soil layers to make use of groundwater. Therefore, the SR in 3-year-old stands was more sensitive to soil water shortage. In addition, the higher soil temperatures in the younger stands due to less canopy coverage possibly accelerated soil vapor evaporation (), which might eventually exacerbate the soil water shortage.
Therefore, soil CO2 efflux increased dramatically with irrigation, resulting mainly from increased fine root growth and carbon mineralization. In addition, SR was closely related to soil temperature in the irrigation treatment, suggesting that soil temperature exerts a dominant control on SR only when sufficient soil moisture availability is ensured.
Plowing increases soil carbon dioxide efflux
Plowing is often used to improve soil aeration and control weeds in poplar plantations. Plowing affects SR mainly by influencing soil microclimate and structure. Although many studies have focused on the effects of plowing on SR in croplands and grasslands (Fortin et al. Citation1996; Dao Citation1998; Curtin et al. Citation2000; Al-Kaisi and Yin Citation2005; Jabro et al. Citation2008; Koga and Tsuji Citation2009), little is known about its effects on SR in forest plantations.
Previous studies suggest that soil disturbance can greatly stimulate soil CO2 emission (Curtin et al. Citation2000; Jabro et al. Citation2008). Dao (Citation1998) reported that soil CO2 efflux under tillage was twice as great as that under no-till conditions in a wheat crop (Triticum aestivum L.). In a grain crop system in eastern Canada, soil CO2 efflux under conventional tillage was 75 g C m−2 greater than that under a no-till condition (Fortin et al. Citation1996). Matsumoto et al. (Citation2008) observed that SR with conventional tillage was 2.1 Mg C ha−1 higher than that of no tillage in the first year after plowing. In the present study, the SR of the three age classes was greatly stimulated by plowing treatment. On average, the SR in plowed stands increased by 36.2% compared with that in the controls. Furthermore, a great pulse of CO2 emission occurred shortly after the plowing, which mainly resulted from quickly stimulated microbial activity due to enhanced carbon availability. After soil disturbance due to plowing, soil bulk density decreased and soil aeration improved greatly (), which was helpful for soil CO2 effusion. In addition, more organic carbon within the soil profile was exposed, leading to greater rates of oxidation and mineralization (Logan et al. Citation1991; Al-Kaisi and Yin Citation2005). Therefore, soil CO2 emission can be greatly accelerated by plowing treatment.
With the depletion of labile substrates, the differences in SR between control and plowed stands tend to decrease. Schlesinger and Andrews (Citation2000) suggest that soil organic matter content declines after soil is disturbed through plowing. As a result, SR in plowed stands may decrease after an initial increase. In the present study, differences in SR between plowing treatment and control stands decreased gradually after an initial pulse of CO2 emission. A year after plowing in June 2008, there was no significant difference in SR between plowing treatment and control stands.
Plowing can also increase soil temperature as a result of decreased residue accumulation on the soil surface (Curtin et al. Citation2000; Al-Kaisi and Yin Citation2005). In the present study, soil temperature in plowing treatment stands was slightly higher than that in the controls (); this was also partly responsible for the higher SR in these stands.
Acknowledgments
This work was jointly supported by the Project of the Key Laboratory of Earth Surface Processes and Resource Ecology (2009-TD-03), Beijing Normal University, and the Natural Science Foundation of China (Grants 40625015, 30770413). The authors would like to thank Drs Jirui Gong, Zhiwei Ge, Xin You, and Qingwei Duan and the Pingyuan Forestry Farm for their work and assistance in the field experiments.
Related Research Data
References
- Al-Kaisi , MM and Yin , X . 2005 . Tillage and crop residue effects on soil carbon and carbon dioxide emission in corn-soybean rotations . J. Environ. Qual. , 34 : 437 – 445 .
- Beukes , DJ . 1984 . Apple root distribution as affected by irrigation at different soil water levels on two soil types . J. Am. Soc. Hortic. Sci. , 109 : 723 – 728 .
- Borken , W , Xu , YJ , Davidson , EA and Beese , A . 2002 . Site and temporal variation of soil respiration in European beech, Norway spruce, and Scots pine forests . Glob. Change Biol. , 8 : 1205 – 1216 .
- Bradshaw , HD Jr , Ceulemans , R , Davis , J and Stettler , R . 2000 . Emerging model systems in plant biology: Poplar (Populus) as a model forest tree . J. Plant Growth Regul. , 19 : 306 – 313 .
- Burton , AJ , Pregitzer , KS , Zogg , GP and Zak , DR . 1998 . Drought reduces root respiration in sugar maple forests . Ecol. Appl. , 8 : 771 – 778 .
- Conant , RT , Dalla-Betta , P , Klopatek , CC and Klopatek , JM . 2004 . Controls on soil respiration in semiarid soils . Soil Biol. Biochem. , 36 : 945 – 951 .
- Coyle , DR and Coleman , MD . 2005 . Forest production responses to irrigation and fertilization are not explained by shifts in allocation . For. Ecol. Manage. , 208 : 137 – 152 .
- Curtin , D , Wang , H , Selles , F , McConkey , BG and Campbell , CA . 2000 . Tillage effects on carbon fluxes in continuous wheat and fallow rotations . Soil Sci. Soc. Am. J. , 64 : 2080 – 2086 .
- Dao , TH . 1998 . Tillage and crop residue effects on carbon dioxide evolution and carbon storage in a Paleustoll . Soil Sci. Soc. Am. J. , 62 : 250 – 256 .
- Davidson , EA , Belk , E and Boone , RD . 1998 . Soil water content and temperature as independent of confounded factors controlling soil respiration in a temperate mixed hardwood forest . Glob. Change Biol. , 4 : 217 – 227 .
- Dickman , DI , Nguyen , PV and Pregitzer , KS . 1996 . Effects of irrigation and coppicing on above-ground growth, physiology and fine-root dynamics of two field-grown hybrid poplar clones . For. Ecol. Manage. , 80 : 163 – 174 .
- Ewel , KC and Cropper , WP . 1987 . Soil CO2 evolution in Florida slash pine plantation I . Changes through time. Can. J. Forest Res. , 17 : 325 – 329 .
- Fortin , MC , Rochette , P and Pattey , E . 1996 . Soil carbon dioxide fluxes from conventional and no-till small grain cropping systems . Soil Sci. Soc. Am. J. , 60 : 1541 – 1547 .
- Gaumont-Guay , D , Black , TA , Griffis , TJ , Barr , AG , Jassal , RS and Nesic , Z . 2006a . Interpreting the dependence of soil respiration on soil temperature and water content in a boreal aspen stand . Agric. For. Meteorol. , 140 : 220 – 235 .
- Gaumont-Guay , D , Black , TA , Griffis , TJ , Barr , AG , Morgenstern , K , Jassal , RS and Nesic , Z . 2006b . Influence of temperature and drought on seasonal and interannual variations of soil, bole and ecosystem respiration in a boreal aspen stand . Agric. For. Meteorol. , 140 : 203 – 219 .
- Goulden , ML , Munger , JW , Fan , SM , Daube , BC and Wofsy , SC . 1996 . Exchange of carbon dioxide by a deciduous forest: response of interannual climate variability . Science , 271 : 1576 – 1578 .
- Grace , J and Rayment , M . 1999 . Respiration in the balance . Nature , 404 : 819 – 820 .
- Jabro , JD , Sainju , U , Stevens , WB and Evans , RG . 2008 . Carbon dioxide flux as affected by tillage and irrigation in soil converted from perennial forages to annual crops . J. Environ. Manage. , 88 : 1478 – 1484 .
- Koga , N and Tsuji , H . 2009 . Effects of reduced tillage, crop residue management and manure application practices on crop yields and soil carbon sequestration on an Andisol in northern Japan . Soil Sci. Plant Nutr. , 55 : 546 – 557 .
- Lal , R . 2005 . Forest soils and carbon sequestration . For. Ecol. Manage. , 220 : 242 – 258 .
- Laureysens , I , Bogaert , J , Blust , R and Ceulemans , R . 2004 . Biomass production of 17 poplar clones in a short-rotation coppice culture on a waste disposal site and its relation to soil characteristics . For. Ecol. Manage. , 187 : 295 – 309 .
- Logan , TJ , Lal , R and Dick , WA . 1991 . Tillage systems and soil properties in North America . Soil Tillage. Res. , 20 : 241 – 270 .
- Maier , CA and Kress , LW . 2000 . Soil CO2 evolution and root respiration in 11-year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability . Can. J. For. Res. , 30 : 347 – 359 .
- Matsumoto , N , Paisancharoen , K and Hakamata , T . 2008 . Carbon balance in maize fields under cattle manure application and no-tillage cultivation in northeast Thailand . Soil Sci. Plant Nutr. , 54 : 277 – 288 .
- Monclus , R , Dreyer , E , Villar , M , Delmotte , FM , Delay , D , Petit , JM , Barbaroux , C , Thiec , DL , Bréchet , C and Brignolas , F . 2006 . Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoids × Populus nigra . New Phytol. , 169 : 765 – 777 .
- Murray , BC , Prisley , SP , Birdsey , RA and Sampson , NR . 2000 . Carbon sinks in the Kyoto Protocol . J. Forestry , 98 : 6 – 11 .
- O’Neill , KP , Kasischke , ES and Richter , DD . 2002 . Environmental controls on soil CO2 flux following fire in black spruce, white spruce, and aspen stands of interior Alaska . Can. J. For. Res. , 32 : 1525 – 1541 .
- Orchard , VA and Cook , FJ . 1983 . Relationship between soil respiration and soil moisture . Soil Biol. Biochem. , 15 : 447 – 453 .
- Pregitzer , KS and Friend , AL . 1996 . “ The structure and function of Populus root systems ” . In Biology of Populus and its Implications for Management and Conservation , Edited by: Stettler , RF , Bradshaw , HD Jr , Heilman , PE and Hinckley , TM . 331 – 354 . Ottawa : NRC Research Press .
- Puri , S and Thompson , FB . 2003 . Relationship of water to adventitious rooting in stem cuttings of Populus species . Agrofor. Syst. , 58 : 1 – 9 .
- Raich , JW and Schlesinger , WH . 1992 . The global carbon-dioxide flux in soil respiration and its relationship to vegetation and climate . Tellus , 44B : 81 – 99 .
- Raich , JW and Tufekcioglu , A . 2000 . Vegetation and soil respiration: correlations and controls . Biogeochemistry , 48 : 71 – 90 .
- Rey , A , Pegoraro , E and Tedeschi , V . 2002 . Annual variation in soil respiration and its components in a coppice oak forest in Central Italy . Glob. Change Biol. , 8 : 851 – 866 .
- Roberts , WP and Chan , KY . 1990 . Tillage-induced increases in carbon dioxide from the soil . Soil Tillage. Res. , 17 : 143 – 151 .
- Saiz , G , Byrnew , EA , Butterbach-bahl , K , Kiese , R , Blujdea , V and Farrell , EP . 2006 . Stand age-related effects on soil respiration in a first rotation Sitka spruce chronosequence in central Ireland . Glob. Change Biol. , 12 : 1 – 14 .
- Schimel , DS . 1995 . Terrestrial ecosystems and the carbon cycle . Glob. Change Biol. , 1 : 77 – 91 .
- Schlesinger , WH and Andrews , JA . 2000 . Soil respiration and the global carbon cycle . Biogeochemistry , 48 : 7 – 20 .
- Smith , P , Powlson , DS , Glendining , MJ and Smith , JU . 1997 . Potential for carbon sequestration in European soils: preliminary estimates for five scenarios using results from long-term experiments . Glob. Change Biol. , 3 : 67 – 79 .
- Swamy , SL , Mishra , A and Puri , S . 2006 . Comparison of growth, biomass and nutrient distribution in five promising clones of Populus deltoides under an agrisilviculture system . Bioresource Technol. , 97 : 57 – 68 .
- Xu , M and Qi , Y . 2001 . Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California . Glob. Change Biol. , 7 : 667 – 677 .