Abstract
Molybdenum (Mo) and selenium (Se) are both essential micronutrients for animals and humans. Increasing Mo and Se contents in food crops offers an effective approach to reduce Mo and Se deficiency problems. A hydroponic trial was conducted to investigate the interactions of Mo and Se on uptake, transfer factors (TF shoot ) as well as distribution coefficients (DC) of Mo and Se on Chinese cabbage (Brassica campestris L. ssp. Pekinensis). In Experiment 1 three concentrations of Mo (0.01, 0.1 and 1 mg L−1) and four concentrations of Se (0, 0.01, 0.1 and 1 mg L−1) were arranged with a randomized block design. In Experiment 2, there were three treatments, 0.1 mg L−1 Mo, 0.1 mg L−1 Se and a combination of 0.1 mg L−1 Mo + 0.1 mg L−1 Se. Experiment 1 showed that Se decreased Mo concentrations in shoots and roots. The impact of Mo on Chinese cabbage response to uptake of Se varied, depending on whether the root Se concentration was saturated or not; Experiment 2 showed that there is a strong antagonism between Mo and Se on nutrition uptake when Mo and Se deficiencies persist for long periods; Mo and Se were easily translocated from solution to plants and from roots to shoots. The results will also be of help in cultivating Mo-enriched and Se-enriched crops.
1. Introduction
Selenium (Se) is an essential trace element for human health. Epidemiological studies have shown that a low intake of Se may increase the risk of Keshan disease, Kashin-Beck disease, cardiovascular disease, muscle syndrome, liver disease and many cancers (Liu et al. Citation2004; Arinola and Charles-Davies Citation2008). An important consideration in the agronomic biofortification of molybdenum (Mo) concerns the effect of Se fertilization, because Mo is also an essential trace element for human beings. Similar to Se deficiency, Mo deficiency in the human body may also increase the risk of Keshan disease, Kashin-Beck disease and many cancers (Chen Citation2008). Furthermore, the interaction between Se and Mo in humans was evident.
In China, approximately 2/3 of the land is Se deficient and often results in human Se deficiency in this area (Cao et al. Citation2001). A study by Wang and Cai (Citation2006) on the dietary intake of Se by Chinese people showed that the average intake is only 28–40 µg per day, lower than the reference nutrient intake of 60–75 µg per day (Broadley et al. Citation2006). Although Se is not proven to be essential for plant nutrition, application of Se in the soil is highly recommended as an approach to eliminate Se deficiency in human diet (Stroud et al. Citation2010). In China, for Se supplementation the earliest used approach is Se-enriched tea. Numerous studies have shown that application of Se in soil can promote plant growth, increase activities of antioxidant enzymes (Djanaguiranman et al. Citation2005; Cartes et al. Citation2010) and enhance chlorophyll concentrations (Zhang et al. 2002).
Mo is an essential micronutrient for plants; however, the shortage of Mo in soil can result in Mo deficiency in human beings. According to the National Soil Survey Office of China (Citation1998), Mo deficiencies predominantly exist in the regions of mass crop production areas such as Eastern China, Central China, and Southern China as well as along the Yangtze River Basin. Mo deficiencies have a negative impact on the quality and production of crops. Application of Mo could increase growth, photosynthesis, chlorophyll, antioxidative capacity of plants and so on (Sun et al. Citation2006; Liu et al. 2010b).
Previous studies have focused more on Se-sulfur (S), Mo-S, Se-phosphorus (P) and Mo-P interactions in soil and solution culture, but little attention has been paid to the possible interactions of Mo-Se in soil or in hydroponic culture (Alhendawi et al. Citation2005; Mora et al. Citation2008; Togay et al. Citation2008; Lee et al. Citation2011a). Although Mo and Se do not belong to the same periodic group, they are both absorbed by plants in form of anions. In low soil pH, bioavailability of Mo and Se both trend to decrease (Rashid et al. Citation2002; Eich-Greatorex et al. 2010; Lee et al. Citation2011b). The contents of Mo and Se in soil are closely related to the concentrations of Mo and Se in the mother rocks. In China, large areas of acidic soil and even alkaline soil are Se-deficient, and most of those areas are also Mo-deficient (Liu et al. Citation1982; Wei Citation1990). Therefore, in the growth of Mo- and Se-enriched crops, it is essential to investigate the interactions between Mo and Se.
Some effective measures are required to ensure sufficient Mo and Se supplementation in the food chain; hence Chinese cabbage (Brassica campestris L. ssp. Pekinensis) stands a high chance as an effective supplement of Mo and Se because it has high weather tolerant abilities, it can be grown throughout the year in China and has a high consumption rate. The aims of this study are, therefore, to investigate the uptake of Mo and Se by Chinese cabbage, and to determine whether there are significant interactions between the two elements in plant nutrient absorption and translocation processes.
2. Materials and Methods
2.1 Plant culture in Experiment 1
The Chinese cabbage “wuyueman” seeds were disinfected in 0.5% sodium hypochlorite (NaClO) solution and germinated in the solution culture of deionized water at 25°C for 7 d. After germination, seedlings of similar size were transplanted in plastic pots containing 3 L of nutrient solutions comprised of 492 mg L−1 calcium nitrate (Ca(NO3)2 · 4H2O), 1020 mg L−1 potassium nitrate (KNO3), 230 mg L−1 ammonium phosphate (NH4H2PO4), 490 mg L−1 magnesium sulfate (MgSO4 · 7H2O), 20 mg L−1 ethylenediaminetetraacetic acid-ferric sodium salt (EDTA-Fe), 2.86 mg L−1 boric acid (H3BO3), 1.54 mg L−1 manganese chloride (MnCl2 · 4H2O), 0.22 mg L−1 zinc sulfate (ZnSO4 · 7H2O) and 0.08 mg L−1 copper sulfate (CuSO4 · 5H2O). Three levels of Mo treatment (0.01, 0.1 and 1 mg L−1) and four levels of Se treatment (0, 0.01, 0.1 and 1 mg L−1) were arranged in a randomized block design. Ammonium orthomolybdate ((NH4)2 MoO4 · 4H2O) and sodium selenite (Na2SeO3) of analytical grade were used as Mo and Se fertilizers. Each treatment was replicated four times, and each pot had eight plants. Thirty-nine days after transplanting, samples from different treatments were collected.
2.2 Plant culture in Experiment 2
Seedlings of Chinese cabbage were sown in plastic pots filled with the above-mentioned basal solution (deficient in Mo and Se) for 32 d, and then plants were divided into three groups. The three groups were prepared to form three treatments: 0.1 mg L−1 Mo, 0.1 mg L−1 Se, and a mixture of 0.1 mg L−1 Mo + 0.1 mg L−1 Se. Each treatment was replicated 24 times; four replicates per treatment were harvested 0, 0.5, 1, 2, 4 and 7 d after the treatment period.
2.3 Plant culture environment
One-fourth and half-strength nutrient solution was added in the first and second weeks, respectively, and then full-strength solution was used until harvest. The solution was renewed once every 3 d to maintain the concentration of nutrient solution throughout the experiment. All vessels used in the experiment were soaked in 5% hydrochloric acid (HCl) for one week, and then washed with deionized water at least four times. Deionized water and analytical grade chemical regents were used in the preparation of the nutrient solution to minimize possible contaminations of Mo and Se. The plants were grown in a greenhouse with natural light at a temperature of 15–30°C.
2.4 Mo and Se analysis
Samples for each treatment were collected, sorted into roots and shoots, desorption-tested for ions with a solution containing 0.5 mM calcium chloride (CaCl2) and 2 mM 2-morpholinoethanesulphonic (pH 6.0), washed several times with deionized water, and oven dried at 65°C. Oven-dried samples were weighed and then sieved through a 1-mm nylon sieve for the analysis of Mo and Se.
Mo was assayed by the method of Wan et al. (Citation1988). Plant samples were first carbonized and then ashed at 525°C for 8 h. Mo concentrations were determined using a JP-4000 Oscilloscope Polarograph in 2 mL 0.25 molL−1 sulphuric acid, 1 mL 0.5 molL−1 benzohydroxyacetic acid and 5 mL saturated sodium chlorate solution.
For Se assays, an oven-dried sample was digested with 8 mL ultra-pure nitric acid and 2 mL perchloric acid at 170°C. The acid mixture was heated until white smoke appeared, and then 10 mL 6 molL−1 hydrochloric acid was added until the white smoke appeared again. The residual was then diluted with 25 mL ultra-pure water. Concentration of Se in the solution was analyzed using an 8200 Atomic Fluorescence Spectrometer (GB/T Citation2008).
2.5 Data analysis
Solution-to-shoot transfer factors (TF shoot ) were defined as the ratio of total Mo or Se in shoots to the corresponding concentration of Mo or Se in solution. Nutrient distribution coefficients (DC) were measured as the ratio of total Mo or total Se in shoots to those in roots (Zhu et al. Citation2004).
The data was statistically analyzed using the analysis of variance (ANOVA) procedure provided by SPSS 12.0 software. The main effects of factors and their interactions were assessed by F statistics, and the mean values of each treatment underwent multiple comparisons using the LSD test at the p < 0.05 level.
3. Results
3.1 Effects of Mo and Se on biomass of Chinese cabbage
shows the shoot and root biomass of Chinese cabbage grown in the solution culture at different concentrations of Mo and Se. Shoot biomass was significantly (p < 0.01) affected by Mo and Se concentrations in the solution. Compared with 0.01 mg L−1 Mo treatments, application of 0.1 and 1 mg L−1 solution Mo both increased dry matter of shoots at the four Se levels, and significantly increased (p < 0.05) shoot dry matter at the levels of 0.1 and 1 mg L−1 solution Se. With additional Se, shoot dry matter increased initially and then decreased at three levels of Mo, and the highest shoot dry matter of 13.89 g pot−1 occurred at level 0.1 mg L−1 of Mo and Se solution. Dry matter of roots was influenced by solution Mo concentrations and interactions of Mo and Se (p < 0.05). Application of 0.1 mg L−1 Mo solution increased dry matter of roots at four levels of Se. Root dry matter was significantly increased by application of Mo only in the absence of Se fertilizer.
Table 1. Biomass of Chinese cabbage (Brassica campestris L. ssp. Pekinensis) in different molybdenum (Mo) and selenium (Se) concentrations in solution culture (mean ± SE) (g pot−1, dry matter basis)
3.2 Effects of Mo and Se on the Mo concentrations of Chinese cabbage
In Experiment 1, concentration of Mo in shoots and roots significantly (p < 0.01) increased with an increase of Mo concentration in nutrients solution (). With the concentration of Mo being 1 mg L−1 in solution, the addition of Se significantly (p < 0.05) inhibited the accumulation of Mo in the shoots, but has increased accumulation of Mo in the roots. TF shoot values of Mo varied from about 69 to 470 (), and decreased significantly (p < 0.01) with addition of Mo in the solution. At 1 mg L−1 of Mo concentration, an addition of Se significantly (p < 0.05) reduced the TF shoot values of Mo. In Experiment 2 (), concentrations of Mo in shoots and roots both increased with time over the next 7 d with or without Se treatment, but the differences between the two treatments were not significant.
Figure 1. Molybdenum (Mo) concentrations in shoot and root of Chinese cabbage (Brassica campestris L. ssp. Pekinensis) grown in solution culture with different concentrations of Mo and selenium (Se). Bars indicate standard error (n = 4). Different letters indicate significant differences at p < 0.05.
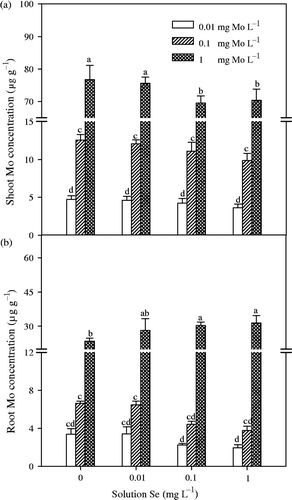
Figure 2. Solution to shoot transfer factors (TF shoot ) for molybdenum (Mo) and selenium (Se) in Chinese cabbage (Brassica campestris L. ssp. Pekinensis) grown at different Mo and Se concentrations in solution culture. Bars mean standard errors (n = 4). Different letters indicate significant differences at p < 0.05.
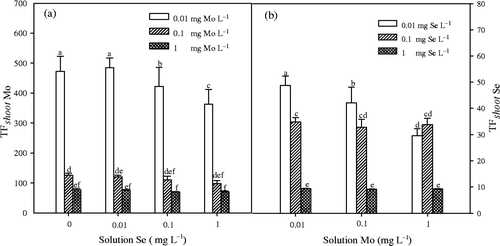
Figure 3. Effects of selenium (Se) treatment on molybdenum (Mo) concentrations in shoot and root of Chinese cabbage (Brassica campestris L. ssp. Pekinensis) in Experiment 2. Bars indicate standard error (n = 4). Different letters indicate significant differences at p < 0.05.
3.3 Effects of Mo and Se on the Se concentrations of Chinese cabbage
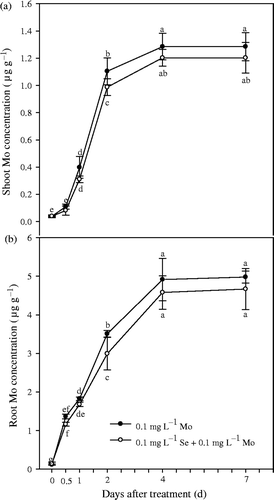
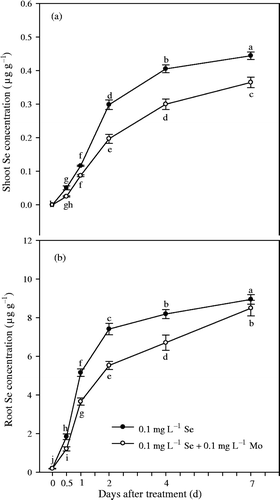
Experiment 1 showed that concentrations of Se in shoots and roots significantly (p < 0.01) increased with the addition of Se in a nutrient solution (). A trace amount of Se was detected in Chinese cabbage grown in solution without additional Se, thus indicating that the plants may have acquired Se from the atmosphere through foliar absorption. Concentration of Mo in the solution had no significant effect on root and shoot Se concentrations at 0.1 and 1 mg L−1 solution Se, but significantly decreased root and shoot Se concentrations at 0.01 mg L−1 solution Se level in Experiment 1. TF shoot values for Se varied from about 9.2 to 48.8 (). Similar to Mo TF shoot values, TF shoot values for Se significantly (p < 0.01) decreased with an increasing solution Se concentration, and were significantly affected by the addition of Mo at 0.01 mg L−1 solution Se. However, it is obvious that Se concentrations in shoots and roots were significantly (p < 0.05) inhibited by application of Mo after Mo and Se deficiencies persisted for 32 d in Experiment 2 ().
Figure 4. Selenium (Se) concentrations in shoot and root of Chinese cabbage (Brassica campestris L. ssp. Pekinensis) grown in solution culture with different concentrations of molybdenum (Mo) and Se. Bars indicate standard error (n = 4). Different letters indicate significant differences at p < 0.05.
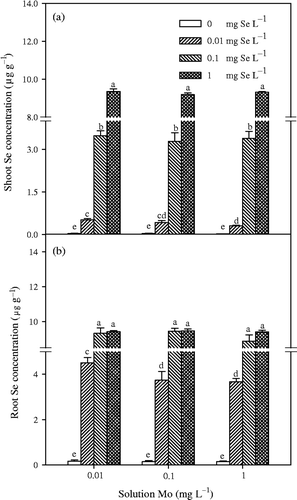
Figure 5. Effects of molybdenum (Mo) treatment on selenium (Se) concentrations in shoot and root of Chinese cabbage (Brassica campestris L. ssp. Pekinensis) in Experiment 2. Bars indicate standard error (n = 4). Different letters indicate significant differences at p < 0.05.
3.4 Effects of Mo and Se on the distribution coefficients of Mo and Se between shoots and roots of Chinese cabbage
Distribution coefficients (DC) of Mo were dramatically (p < 0.01) influenced by solution Mo concentration and interactions of Mo and Se. DC values of Mo ranged from 1.4 to 3.0 (). Compared to 0.01 mg L−1 of Mo treatments, application of 0.1 and 1 mg L−1 of Mo solution both increased DC of Mo at four solution Se levels. At Mo concentrations of 0.01 and 0.1 mg L−1 in the solution, the addition of Se continuously increased the DC of Mo. However, at the 1 mg L−1 concentration of Mo, a decreasing trend with addition of Se was observed.
Table 2. Distribution coefficients (DC) of molybdenum (Mo) and selenium (Se) between shoots and roots of Chinese cabbage (Brassica campestris L. ssp. Pekinensis) in solution culture (mean ± SE)
DC of Se was dramatically (p <0.01) influenced by the concentration of the Se solution. With the increased rates of Se application, DC of Se decreased initially and then increased at three solution Mo levels. DC of Se was decreased by extra application of Mo, and the decreases were more evident in the absence of Se fertilizer.
4. Discussion
4.1 Mo and Se had antagonistic effects on Mo and Se uptakes in Chinese cabbage
Although it is recognized that Mo and Se are closely related biologically in animals (Bogden et al. Citation1986), interactions between Mo and Se fertilizers on plant growth as well as on Mo and Se uptake processes have never been reported. However, a significant antagonistic effect on Mo and Se uptakes between Mo and Se fertilizers was observed in our experiment (). For example, an increase in solution of Se from 0 to 1 mg L−1 decreased root and shoot Mo concentrations at levels of 0.01 and 0.1 mg L−1 of Mo solution in Experiment 1. Furthermore, application of Mo decreased the concentration of Se in roots and shoots at 0.01 mg L−1 solution Se in Experiment 1. These results correspond with previous studies; i.e., several reports have verified that Mo and Se were both transported via sulphate and phosphate transporters and that these anions will compete with one another to interact with the transporters (Kumar and Singh Citation1980; Hopper and Parker Citation1999; Ramos et al. Citation2011).
Table 3. Summary of the analysis of variance showing F-ratio for solution to shoot transfer factors (TF shoot ), molybdenum (Mo) and selenium (Se) concentrations in a double factorial experiment
Mo is transported not only passively via sulfate and phosphate transporters but also actively via single-minded transport proteins MOT1 and AMA1 (Palmgren and Harper Citation1999; Tomatsu et al. Citation2007). Se is only passively taken up by plants, and sulfate and phosphate transporters are believed to act in the process (Terry et al. Citation2000; Li et al. Citation2008). Hence, Mo has a greater impact on Se uptake than Se on Mo. Similarly, Experiment 2 showed that application of Mo significantly decreased plant uptake of Se, while application of Se only slightly inhibited plant uptake of Mo.
4.2 Se decreased Mo concentration and inhibited translocation of excess Mo from roots to shoots in Chinese cabbage
Due to competition, application of Se reduced root and shoot Mo concentrations in Chinese cabbage at ≤ 0.1 mg L−1 solution Mo. Application of Se significantly increased concentration of Mo in root at 1 mg L−1 Mo solution. However, shoot Mo concentrations were dramatically decreased by application of Se at 1 mg L−1 solution Mo. At 1 mg L−1 solution Mo treatments, root Mo contents were approximately 30 mgkg−1. Since plants have a superior capacity for alleviating the toxicity of high Mo (Lu Citation1994), Chinese cabbage biomass was not significantly reduced with increased levels of Mo in this experiment. However, our results implied that the application of Se could still inhibit excess Mo translocation from roots to shoots. Therefore, more Mo was distributed to the root in Chinese cabbage, and Mo nutrition status for the root was also improved. Although results showed that application of Se increased root Mo concentration in 1 mg L−1 solution Mo, the plant total Mo concentrations had been reduced.
4.3 The impact of Mo on Chinese cabbage response to uptake of Se varied, depending on whether the root Se concentration was saturated or not
Application of Mo significantly decreased the concentrations of Se in shoots and roots at 0.01 mg L−1 in Se solution level, but application of Mo did not affect roots and shoots in Se concentrations at 0.1 and 1 mg L−1 Mo solution levels. Results also showed that concentration of Se in roots remained constant at ≥ 0.1 mg L−1 Se solution level, because the Se-binding site was saturated. Previous results also showed that Se accumulation in plants was enhanced disproportionately with increasing Se concentration in the growth environment (Wu et al. Citation1988); e.g., Hu (Citation2011) found that concentration of Se in roots showed a pattern of saturation in relation to the concentration of Se in the soil. It is worth noting that concentrations of Se in shoots and roots in Experiment 1 were not affected by application of Mo at ≥ 0.1 mg L−1 Se solution level. Though the result did not correspond with Experiment 2, this phenomenon is not a surprise. In fact, Experiment 2 indicated that application of Mo delayed the time for Se in root to reach saturation at ≥ 0.1 mg L−1 Se solution levels. Under the pattern of saturation, the absorptive capacity of Se depended on the ability of plant translocation of Se from roots to shoots, while application of Mo did not affect translocation of Se from roots to shoots (). Mo could inhibit plant uptake of Se, but the antagonism would be reduced when plant root Se concentration was saturated. In association with the results ahead, we can deduce that the impact of Mo on Chinese cabbage response to Se uptake varied, depending on whether the root Se concentration was saturated or not.
4.4 Dual supplementation of Mo and Se in Chinese cabbage is feasible
The ultimate goal of this study was to investigate the feasibility of dual fortification of Mo and Se in the human food chain. Hence, increasing fertilizer uptake efficiency is the key to produce Mo-and Se-enriched foods. Root surface area and active absorption area can be used to evaluate capacity of nutrient uptake (Yang et al. Citation2004). It has been reported that application of Se can increase root activity as well as development of the root system (Tang et al. Citation2005), and application of Mo can improve the growth of the taproot and lateral root and increase root total absorption areas (Wang Citation2003). Though there was a negative synergistic effect on TF shoot values of Mo and Se in this experiment, both Mo and Se can be easily translocated from solution to plants. The results indicate that addition of Mo and Se fertilizers on Chinese cabbage were highly effective in raising root nutrient uptake ability. The similar effect of application of Mo and Se fertilizers on plant nutrient uptake and translocation efficiencies were also reported in rapeseed (Brassica napus L.) (Liu et al. 2010a) and spinach (Spinacia oleracea L.) (Zhu et al. Citation2004).
In terms of Mo and Se supplementation of human diets for enhanced nutrition, the accumulation of these two elements in edible parts of Chinese cabbage is very important. Results from this study showed that Mo is easily accumulated in shoots, with shoots to roots DC for Mo ranging from 1.40 to 3.01 (). As concentrations of Se solution increase, the DC for Se were raised at three levels of Mo solution. These results correspond with previous work; e.g., Sors et al. (Citation2005) found that Se can be easily translocated from roots to shoots if plants were fed with sufficient selenate. The DC values for Mo obtained in this study were similar to those reported by Liu (Citation2009), and the DC values for Se are comparable to the results reported by Zayed et al. (Citation1998). Hence, the results suggest that both Mo and Se can be easily translocated from roots to edible parts.
5. Conclusions
This study demonstrates complex interactions between Mo and Se involving both nutrition uptake and translocation. Concentrations of Mo in plants were decreased by solution Se concentrations at 0.01 and 0.1 mg L−1 solution Mo levels, and Se had a reduced effect on plant translocation of Mo from roots to shoots at the higher Mo rate. The impact of Mo fertilization on Chinese cabbage response to uptake of Se varied, depending on whether the root Se concentration was saturated or not. Mo has a greater impact on Se uptake than Se on Mo. There has a strong antagonism between Mo and Se on nutrition uptake, especially when Mo and Se deficiency persist for long periods. Results also showed that both Mo and Se can be easily translocated from solution to plants and from roots to edible parts.
Acknowledgments
The project was sponsored by the Fundamental Research Funds for the Central Universities (2010PY025, 2010PY150) and Innovation Funds of Huazhong Agricultural University (52902-0900206044).
Related Research Data
References
- Alhendawi , RA , Kirkby , EA and Pilbeam , DJ . 2005 . Evidence that sulfur deficiency enhances molybdenum transport in xylem sap of tomato plants . J. Plant Nutr. , 28 : 1347 – 1353 .
- Arinola , OG and Charles-Davies , MA . 2008 . Micronutrient levels in the plasma of Nigerian females with breast cancer . Afr. J. Biotechnol. , 7 : 1620 – 1623 .
- Bogden , JD , Chung , HR , Kemp , FW , Holding , K , Bruening , KS and Naveh , Y . 1986 . Effect of selenium and molybdenum on methylbenzylnitrosamine-induced esophageal lesions and tissue trace metals in the rat . J. Nutr. , 116 : 2432 – 2442 .
- Broadley , MR , White , PJ Bryson , RJ . 2006 . Biofortification of UK food crops with selenium . Proc. Nutr. Soc. , 65 : 169 – 181 .
- Cao , ZH , Wang , XC , Yao , DH , Zhang , XL and Wong , MH . 2001 . Selenium geochemistry of paddy soil in Yangtze River delta . Environ. Int. , 26 : 335 – 339 .
- Cartes , P , Jara , AA , Pinilla , L , Rosas , A and Mora , ML . 2010 . Selenium improves the antioxidant ability against aluminium-induced oxidative stress in ryegrass roots . Ann. Appl. Biol. , 156 : 297 – 307 .
- Chen , BY . 2008: The anti-cancer effects of trace elements molybdenum. Studies of Trace Elements and Health, 25, 68–69
- Djanaguiranman , M , Devi , DD , Shanker , AK , Sheeba , A and Bangarusamy , U . 2005 . Selenium—an antioxidative protectant in soybean during senescence . Plant Soil , 272 : 77 – 86 .
- Eich-Greatorex , S , Krogstad , T and Sogn , TA . 2010 . Effect of phosphorus status of the soil on selenium availability . J. Plant Nutr. Soil Sci. , 173 : 337 – 344 .
- GB/T 21729 2008: Determination of selenium contents in tea. Standard Press of China, Beijing
- Hopper , JL and Parker , DR . 1999 . Plant availability of selenite and selenate as influenced by the competing ions phosphate and sulfate . Plant Soil , 210 : 199 – 207 .
- Hu , FF . 2011: Study on migration effects of selenium in soil-alfalfa-feed-hen system and its mechanism. (PhD thesis). Huazhong Agricultural University, Wuhan, China
- Kumar , V and Singh , M . 1980 . Interactions of sulfur, phosphorus and molybdenum in relation to uptake and utilization of phosphorus by soybean . Soil Sci. , 130 : 26 – 31 .
- Lee , S , Woodard , HJ and Doolittle , JJ . 2011a . Effect of phosphate and sulfate fertilizers on selenium uptake by wheat (Triticum aestivum) . Soil Sci. Plant Nutr. , 57 : 696 – 704 .
- Lee , S , Woodard , HJ and Doolittle , JJ . 2011b . Selenium uptake response among selected wheat (Triticum aestivum) varieties and relationship with soil selenium fractions . Soil Sci. Plant Nutr. , 57 : 823 – 832 .
- Li , HF , McGrath , SP and Zhao , FJ . 2008 . Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite . New Phytol. , 178 : 92 – 102 .
- Liu , HE . 2009: Study on interaction of molybdenum and phosphorus and its mechanism of Brassica napus. (PhD thesis.) Huazhong Agricultural University, Wuhan, China
- Liu , HE , Hu , CX , Hu , XM , Nie , ZJ , Sun , SC , Tan , QL and Hu , FF . 2010a . Interaction of molybdenum and phosphorus supply on uptake and translocation of phosphorus and molybdenum by Brassica napus . J. Plant Nutr. , 33 : 1751 – 1760 .
- Liu , HE , Hu , CX , Sun , SC , Tan , QL , Nie , ZJ and Hu , XM . 2010b . Interactive effects of molybdenum and phosphorus fertilizers on photosynthetic characteristics of seedlings and grain yield of Brassica napus . Plant Soil , 326 : 345 – 353 .
- Liu , Q , Wang , DJ , Jiang , XJ and Cao , ZH . 2004 . Effects of the interactions between selenium and phosphorus on the growth and selenium accumulation in rice(Oryza sativa) . Environ. Geochem. Health , 26 : 325 – 330 .
- Liu , Z , Zhu , QQ , Tang , LH , Xu , JX and Yen , CL . 1982: Geographical distribution of trace elements—deficient soils in China. Acta Pedol. Sin., 19, 209–223
- Lu , JL . 1994 . Plant Nutrition , 2nd , Beijing : China Agricultural Press .
- Mora , ML , Pinilla , L , Ross , A and Cartes , P . 2008 . Selenium uptake and its influence on the antioxidative system of white clover as affected by lime and phosphorus fertilization . Plant Soil , 303 : 139 – 149 .
- National Soil Survey Office of China . 1998 . “ Soils of China ” . In National Soil Survey , Beijing : Chinese Agriculture Press .
- Palmgren , MG and Harper , JF . 1999 . Pumping with plant P-type ATPases . J. Exp. Bot. , 50 : 883 – 893 .
- Ramos , SJ , Rutzke , MA , Hayes , RJ , Faquin , V , Guilherme , L and Li , L . 2011 . Selenium accumulation in lettuce germplasm . Planta , 233 : 649 – 660 .
- Rashid , M , Kang , YM and Sakurai , K . 2002 . Selenium in amorphous iron (hydr)oxide-applied soil as affected by air-drying and pH . Soil Sci. Plant Nutr. , 48 : 243 – 250 .
- Sors , TG , Ellis , DR and Salt , DE . 2005 . Selenium uptake, translocation, assimilation and metabolic fate in plants . Photosynth. Res. , 86 : 373 – 389 .
- Stroud , JL , Li , HF Lopez-Bellido , FJ . 2010 . Impact of sulphur fertilisation on crop response to selenium fertilisation . Plant Soil , 332 : 31 – 40 .
- Sun , XC , Hu , CX and Tan , QL . 2006 . Effects of molybdenum on anti-oxi-dative defense system and membrane lipid peroxidation in winter wheat under low temperature stress . J. Plant Physiol. Mol. Biol. , 2 : 175 – 782 .
- Tang , QY , Wu , YY , Zhou , DZ and Zhou , YF . 2005 . Effect of Se on root activity of soybean . Henan Agric. Sci. , 7 : 42 – 43 .
- Terry , N , Zayed , AM , de Souza , MP and Tarun , AS . 2000 . Selenium in higher plants . Ann. Rev. Plant Physiol. Plant Mol. Biol. , 51 : 401 – 432 .
- Togay , Y , Togay , N and Dogan , Y . 2008 . Research on the effect of phosphorus and molybdenum applications on the yield and yield parameters in lentil . Afr. J. Biotechnol. , 7 : 1256 – 1260 .
- Tomatsu , H , Takano , J , Takahashi , H , Takahashi , AW , Shibagaki , N and Fujiwara , T . 2007 . An Arabidopsis thaliana high-affinity molybdate transporter required for efficient uptake of molybdate from soil . PNAS , 104 : 18807 – 18812 .
- Wan , YH , Liu , XD and Li , ZY . 1988 . Determination of soil available molybdenum and plant molybdenum by polarographic catalytic wave analysis . Chin. J. Soil Sci. , 19 : 43 – 46 .
- Wang , HJ . 2003: The effect of molybdenum on root physiology of rape and two winter wheat cultivars. (Master's thesis.) Huazhong Agricultural University, Wuhan, China
- Wang , JM and Cai , JF . 2006 . Effects of selenium applications on the selenium content, yield and quality of garlic . Chin. Agric. Sci. Bull. , 4 : 342 – 344 .
- Wei , FS . 1990 . Background values of elements in Chinese soil , Beijing : China Environmental and Science press .
- Wu , L , Huang , ZZ and Burau , GR . 1988 . Selenium accumulation and selenium-salt cotolerance in five grass species . Crop Sci. , 28 : 517 – 522 .
- Yang , CM , Yang , LZ , Yang , YX and Zhu , OY . 2004 . Rice root growth and nutrient uptake as influenced by organic manure in continuously and alternately flooded paddy soils . Agric. Water Manage. , 70 : 67 – 81 .
- Zayed , A , Lytle , CM and Terry , N . 1998 . Accumulation and volatilization of different chemical species of selenium by plants . Planta , 206 : 284 – 292 .
- Zhang , C , Wu , YY and Peng , ZK . 2002: Influence of Selenium on the content of chloroplast pigment in rape seedings. J. Hubei Inst. Nationalities (Natural Science Edition), 20, 63–65
- Zhu , YG , Huang , YZ , Hu , Y , Liu , YX and Christie , P . 2004 . Interactions between selenium and iodine uptake by spinach (Spinacia oleracea L.) in solution culture . Plant Soil , 261 : 99 – 105 .