
Abstract
The objective of this study was to compare the carbon (C) balance of mowed grassland that received either manure slurry plus synthetic fertilizers (slurry plots) or sawdust-containing composted farmyard manure plus synthetic fertilizers (FYM plots). Dairy slurry (65.8–66.4 Mg ha–1 year–1) or FYM (36.5–39.2 Mg ha–1 year–1) was applied to a maximum based on a safe potassium content that met the potassium requirement for forage grass production. Ammonium sulfate and superphosphate were additionally supplied to meet the nitrogen (N) and phosphorus requirements. The C inputs to the soil through manure + plant residues (losses during harvesting) and the C outputs from the soil (i.e., respiration) through decomposition of soil organic matter (Rs) + applied manure (Rm) + plant residues (Rp) were monitored for 2 years. Rs and Rm were estimated by using a closed-chamber method with root exclusion. Rp was estimated by using a litter-bag method. Rs averaged 8.2 Mg-C ha–1 per 2 years. Rm accounted for 84% of the applied slurry C (5.5 Mg-C ha–1 per 2 years), and 23% of applied FYM C (9.0 Mg-C ha–1 per 2 years). Rp accounted for 68–70% of plant-residue C (1.0–1.1 Mg-C ha–1 per 2 years) in the slurry and FYM plots. The C balance of the grassland, estimated as C inputs through manure + plant residues minus C outputs through Rs + Rm + Rp, was – 7.0 Mg-C ha–1 per 2 years in the slurry plots and –1.0 Mg-C ha–1 per 2 years in the FYM plots. Thus, FYM application produced a smaller decrease in the soil organic C by 6.0 Mg-C ha–1 per 2 years in comparison with the slurry application. The decomposability of the applied manure therefore depended on the storage conditions, which have an important effect on the C balance (i.e., increment of SOC). Plant residues (i.e., harvest loss) contributed less than FYM to the C balance.
1. INTRODUCTION
Since the industrial revolution, global anthropogenic carbon (C) emissions have totaled an estimated 270 ± 30 Pg due to fossil fuel combustion, and 136 ± 55 Pg due to land use changes and crop cultivation (Lal Citation2004). Conversion of perennial-dominated, native ecosystems into arable land usually reduces soil organic C (SOC) by reducing plant-derived C inputs, creating soil conditions (e.g., moisture, aeration, temperature) that promote the decomposition of SOC, and exposing SOC that is physically protected in soil aggregates to microbial activity (Gregorich et al. Citation1998). Conversely, the conversion of arable land into permanent pasture is considered to have C sequestration potential (Soussana et al. Citation2004). Management practices to rebuild soil organic matter (SOM) must increase the supply of OM to the soil, reduce the tillage-induced decomposition of SOM, or both (Paustian et al. Citation1997; Smith and Powlson Citation2000).
Management techniques intended to increase fodder production potentially increase SOM by increasing the supply of dead plant material to the soil (Ammann et al. Citation2007; Conant et al. Citation2001). Manure, an organic fertilizer, supplies nutrients to crops while improving the structure of soil aggregates (Bhogal et al. Citation2011; Diacono and Montemurro Citation2010). The rate of manure mineralization after application depends on its storage conditions (Harada et al. Citation1993). The labile organic matter in manure slurry (hereafter, “slurry”) decomposes quickly, releases mineral N rapidly, and its C content returns to the atmosphere (Angers et al. Citation2007). In contrast, the persistent organic matter in composted farmyard manure (FYM) decomposes slowly, releases mineral N gradually, and increases SOM (Shimizu et al. Citation2014a, Citation2014b).
Plant residues are a substantial source of OM for the soil. The introduction of improved grass species with higher productivity could therefore increase SOM (Conant Citation2010; Smith et al. Citation2008). These residues enter the soil as dead plant material, senescent root tissue, sloughed root cells, and root exudates (Gregorich and Janzen Citation1996). In mowed grasslands, plant residues from the aboveground biomass derive mostly from losses during harvesting due to shattering of the leaves and stems into pieces too small to be collected (Rees Citation1982). These harvest losses were estimated to be 12–14% of the dry matter yield of grass (Rotz and Muck Citation1994). The subsequent decomposition of the plant residues supplied by improved pasture species is faster than that of the residues from traditional species (Kirita et al. Citation2002).
Heterotrophic respiration is a critical ecological process because it releases SOC into the atmosphere in the form of CO2. Soil respiration represents the combined activity of the soil microorganisms that decompose litter and SOM (i.e., heterotrophic respiration) and the respiration of plant-assimilated C by roots and associated microorganisms (i.e., root respiration; Singh et al. Citation2011). Heterotrophic respiration can be distinguished from root respiration by measuring the CO2 fluxes of soil from which roots have been excluded (Hanson et al. Citation2000). The rate of heterotrophic respiration depends on soil temperature and moisture, as well as on the quality and quantity of SOM (Davidson and Janssens Citation2006; Zhou et al. Citation2009).
It is crucial to understand the effect of fertility management using livestock manure on the C balance of grasslands, yet insufficient information is available on the effect of different manure types on the C balance. The objectives of the present study were to provide some of this information by (1) quantifying the C inputs to the soil through manure (slurry or FYM) and plant residues and the C outputs from the soil through decomposition (i.e., respiration, R) of soil organic matter (Rs), applied manure (Rm), and plant residues (Rp) in grassland plots that received slurry or FYM plus synthetic fertilizers; and (2) estimating the C balances of the grassland. On the basis of these results, we discuss the effects of manure type on the C balance and the implications for pasture management.
2. MATERIALS AND METHODS
2.1. Study site
Our field experiment was carried out in a mowed grassland at the NARO Institute of Livestock and Grassland Science in Nasu, Japan (36°55′N, 139°55′E, 320 m a.s.l.). The ungrazed grassland had been managed as a meadow for more than 30 years. The grassland was last renovated by cultivating the soil with a plow and a rotovator, followed by seeding of orchardgrass (Dactylis glomerata L.) at a rate of 30 kg ha−1 in August 2006 (Mori and Hojito Citation2007). White clover (Trifolium repens L.) was removed by hand periodically from the study plots. The soil was derived from volcanic ash; Kurashima et al. (Citation1993) classified it as a loamy Entic Humudept, mixed, mesic (Soil Survey Staff Citation2014). The amounts of C and N of the top 30 cm of the soil just before the application of the fertilizer treatments were 126 ± 8 Mg C ha−1 and 7.5 ± 0.3 Mg N ha−1 (n = 3, mean ± SEM). The C and N contents of the top 5 cm of the soil (i.e., the bulk of the rooting zone) were 66.0 ± 3.8 g C kg–1 and 4.19 ± 0.28 g N kg–1 (n = 3, mean ± SEM). The 30-year averages for precipitation and temperature were 1561 mm year–1 and 12.0°C (NGRI Citation2001). There was generally no snow cover during the study. The minimum and maximum mean monthly temperatures were 1.2°C (January) and 23.7°C (August).
2.2. Fertility management
In March 2008, nine plots (each 4 m × 4 m) were established: three with no supplemental N fertilizer (control plots), three with slurry plus synthetic fertilizers (slurry plots), and three with composted FYM plus synthetic fertilizers (FYM plots), in a randomized block design. Slurry (65.8–66.4 Mg ha–1 year–1) or sawdust-containing FYM (36.5–39.2 Mg ha–1 year–1) obtained from nearby commercial dairy farms was applied so that the annual potassium supply from either roughly covered the annual potassium requirement for forage grass production without exceeding the limit that would be safe for livestock ( and ). Supplemental ammonium sulfate and superphosphate were applied to provide any additional annual N and phosphorus (P) requirements. Details of the application of manure and synthetic fertilizers are presented in Mori and Hojito (Citation2015).
Table 1 Manure application rates and their chemical and physical properties
2.3. Measurements of Rs and Rm
We used the root exclusion approach to estimate Rs and Rm (Hanson et al. Citation2000; Prolingheuer et al. Citation2014). In each plot, we established a root-exclusion subplot (1 m × 1 m) by excavating the soil, removing roots and stubble to a depth of 30 cm, lining the holes with water-permeable root barrier sheets (BKS0812, Toyobo, Tokyo, Japan), and replacing the soil.
From March 2008 to March 2010, we measured the CO2 fluxes in the root-exclusion subplots 90 times using a cylindrical stainless-steel chamber (40 cm in diameter, 30 cm in height). The concentration of CO2 in the chamber headspace was monitored at 5-s intervals using a non-dispersive infrared sensor (GMP343 + MI70, Vaisala, Helsinki, Finland). The measurements were performed between 08:00 and 10:00 to minimize the effect of diurnal variation in CO2 fluxes on the cumulative CO2 emission. The CO2 flux measurements were conducted 1, 3, 7, 10, and 14 days after fertilization, and then once a week (once every 2–4 weeks during winter).
The CO2 flux was determined by assuming a linear increase in the CO2 concentrations between 1 and 3 min after the chamber was sealed (Matsuura et al. Citation2011). The CO2 fluxes were calculated as follows:
where F is the CO2 flux (kg-C ha–1 d–1), ρ is the density of CO2 (0.536 kg-C m–3 at 273 K), V is the volume of the chamber headspace (m3), A is the area of the chamber’s base (m2), ∆c/∆t is the temporal change in the CO2 concentration (m3 m–3 h–1), and T is the air temperature (K). The cumulative CO2 emission was calculated by integration of the area under the CO2 flux curve.
Rs was assumed to be the cumulative CO2 emission in the control plots. Rm was estimated by subtracting Rs from the cumulative CO2 emission in the slurry and FYM plots. Rs was defined as the decomposition of SOM but excluding the manure and plant residues supplied during the 2-year period of this field experiment.
2.4. Measurements of dry matter yield and the amount of plant residues
The plots were harvested four times each year: on 27 May, 9 or 10 July, 1 September, and 4 or 5 November. Grass yields were determined by cutting the aboveground biomass in a 1.3 m × 4.0 m area of each plot at a height of 5 cm above the soil surface using a reciprocating mower (HTK8070, STAR Machinery, Chitose, Japan).
The amount of plant residues in the slurry and FYM plots was measured as follows. A quadrate (50 cm × 50 cm) was fixed to the soil surface in each plot using stainless-steel pins. At each harvest, plant residues that resulted from defoliation since the previous harvest and from each new harvest were collected by hand the same day. The plant residues were dried at 70°C for 3 days, and their C concentrations were determined by means of dry combustion using a C-N analyzer (JM1000CN, J-Science Lab Co. Ltd., Kyoto, Japan). The dry matter yield was multiplied by the C and N concentrations to determine the total content per unit area.
2.5. Measurement of Rp
Rp was estimated by using the litter-bag method (Kirita et al. Citation2002). On 25 March 2008, more than 120 nylon-mesh bags (15 cm × 20 cm, 1-mm mesh) were each filled with 8.0 g of air-dried orchardgrass residues and fixed to the surface of the control plots using stainless-steel wires. The C and N contents of the residues were 438 ± 3 g-C kg–1 and 21 ± 1 g-N kg–1 (n = 6, mean ± SEM). Every month between 25 March 2008 and 23 March 2010, five bags were randomly collected from each plot. The plant residues remaining in each bag were dried at 70°C for 3 days and weighed.
The weight loss over time was assumed to be due solely to the decomposition of plant residues. Under these assumptions, we developed a decomposition curve, and used it to estimate the residues remaining on 21 March 2010 (i.e., the end of the field experiment). We then calculated the sum of plant-residue C that remained undecomposed on 21 March 2010 (Lr) as:
where Li is the weight of plant-residue C that had fallen by the ith harvest (g), LBi is the weight of plant residues in a litter bag at the ith harvest (g), and LBe is the weight of plant residues in a litter bag on 21 March 2010.
Rp was estimated by subtracting Lr from the cumulative amount of plant-residue C from the eight harvests.
2.6. Measurement of plant biomass
Plant biomass (i.e., roots and stubble) in the slurry and FYM plots were measured three times: on 26 March 2008, 23 March 2009, and 23 March 2010. The aboveground biomass inside a quadrat (25 cm × 50 cm) was collected by cutting stubble at the soil surface with sharp scissors. Next, litter and manure remaining on the soil surface were removed by hand. The top 30 cm of the soil, which contains the majority of the belowground biomass, was collected using a shovel and a trowel. Finally, the belowground biomass was separated from the soil under a stream of water. The samples were disaggregated into roots and stubble, dried at 70°C for 3 days, and weighed, and their C contents were determined.
2.7. Statistical analyses
Statistical analyses were performed using STATISTICA 6.1 software (StatSoft Inc., Tulsa, OK, USA). Two-way ANOVA was used to examine the effects of treatments and years on the annual CO2 emission, the annual yield, and the annual amount of plant residues. The Kruskal–Wallis test was used to test for significant interannual differences in the amounts of roots and stubble in the slurry and FYM plots.
3. RESULTS
3.1. Carbon dioxide flux
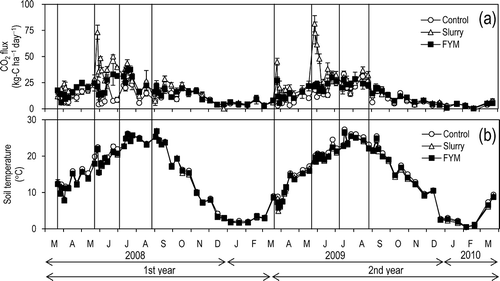
The CO2 fluxes from the control, slurry, and FYM plots ranged from –2 to 33, from 0–81, and from 0–38 kg C ha–1 day–1, respectively (). Sharp CO2 flux peaks were observed following slurry applications, but only slight increases after FYM applications. The change in soil temperature explained most of the seasonal variation in CO2 fluxes (), however, the correlation coefficient between soil temperature and CO2 fluxes in the slurry plots (R2 = 0.29, P < 0.001) was smaller than that in the control (R2 = 0.51, P < 0.001) and FYM plots (R2 = 0.62, P < 0.001) owing to the sharp flux peaks of CO2.
Figure 1 Temporal changes in (a) CO2 flux, (b) temperature at a depth of 5 cm below the soil surface. Values are means ± SEM (n = 3). Vertical lines indicate fertilizer applications.
3.2. Rs and Rm
ANOVA showed a significant difference in the annual CO2 emission among treatments (F = 11.26, P = 0.002, ). No significant interannual difference was observed (F = 2.46, P = 0.143), and there was no significant treatment × year interaction (F = 0.24, P = 0.793). The annual CO2 emission in slurry plots was significantly greater than that in control and FYM plots (P = 0.002, P = 0.069, Scheffé’s test). No difference was observed between the control and FYM plots (P = 0.142, Scheffé’s test).
Table 2 Mineral nitrogen supplied from synthetic fertilizer, slurry, and FYM
Table 3 CO2 emission from grassland, Rs and Rm
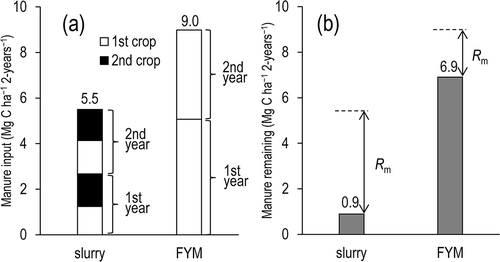
Rs totaled 8.2 Mg-C ha–1 per 2 years (). On the one hand, out of the applied slurry C (5.5 Mg-C ha–1 per 2 years), Rm accounted for 4.6 Mg-C ha–1 per 2 years, suggesting that 84% of the slurry C decomposed in 2 years (, ). On the other hand, out of the applied FYM C (9.0 Mg-C ha–1 per 2 years), Rm accounted for 2.1 Mg C ha–1 per 2 years, suggesting that 23% of the FYM C decomposed in 2 years. Consequently, the amount of manure C that remained undecomposed until the end of the field experiment on 21 March 2010 was estimated to be 0.9 Mg-C ha–1 in the slurry plots and 6.9 Mg-C ha–1 in the FYM plots.
Figure 2 (a) Manure input for each crop and (b) manure that remained on 21 March 2010. FYM, farmyard manure; Rm, manure decomposition.
3.3. Dry matter yield and amount of plant residues
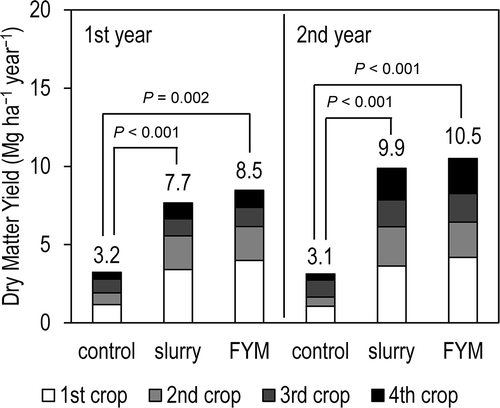
The annual yields (dry matter basis) in the slurry and FYM plots were comparable to the standard annual yields in our region (8–10 Mg ha–1 year–1; Tochigi Prefectural Government Citation2002). In the slurry and FYM plots, the yield of the first crop was greater than those of the second, third, and fourth crops (). ANOVA showed that the annual yield differed significantly among the treatments (F = 167.9, P < 0.001) and years (F = 20.2, P = 0.001), with a significant treatment × year interaction (F = 5.8, P = 0.017). In each of the first and second years, no significant difference was observed between the slurry and FYM plots (P = 0.515 and P = 0.289, respectively; Scheffé’s test).
Figure 3 Dry matter yields (Scheffé’s test). FYM, farmyard manure.
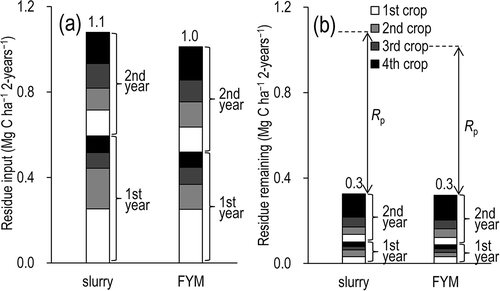
The annual amount of plant residue inputs () represented 12–19% of the annual yield (). ANOVA showed that the annual input of plant residues did not differ significantly between the slurry and FYM plots (F = 0.486, P = 0.506). No significant interannual difference was observed (F = 1.916, P = 0.204), and there was no significant interaction term (F = 0.740, P = 0.415).
Figure 4 (a) Plant residue inputs and (b) amount of plant residues remaining until the end of the study (21 March 2010). FYM, farmyard manure. Rp, decomposition of plant residue.
3.4. Rp
The litter-bag experiment revealed that a total of 74% and 92% of the plant residues decomposed during the first and second years, respectively (). The residues decomposed most rapidly during summer and more slowly in winter. From the decomposition curve (), we can estimate the proportion of the plant residues fallen during the first year that remained until the end of the field experiment: 13% of the residues from the first crop, 17% from the second crop, 21% from the third crop, and 27% from the fourth crop remained undecomposed on 21 March 2010. Similarly, of the plant residues fallen during the second year, 29% from the first crop, 34% from the second crop, 40% from the third crop, and 75% from the fourth crop remained undecomposed on 21 March 2010.
Figure 5 Proportion of the plant-residue mass in the litter bags remaining at various times during the study period. Values are means ± SEM (n = 5). Arrows represent the harvest times for the1st to 4th crops.
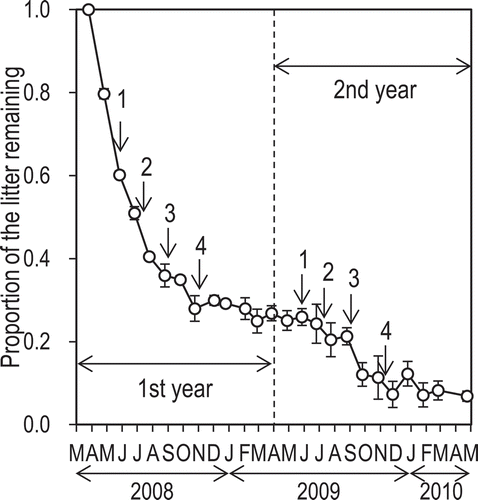
On this basis, the sum of plant-residue C that remained undecomposed until the end of the field experiment (i.e., 21 March 2010) was 0.3 Mg-C ha–1 in both the slurry plots and the FYM plots (, ). Consequently, of the plant-residue C that fell in the slurry plots (1.1 Mg-C ha–1 per 2 years) and FYM plots (1.0 Mg-C ha–1 per 2 years), Rp accounted for 0.8 and 0.7 Mg-C ha–1 per 2 years, respectively, suggesting that 70% and 68% of plant-residue C decomposed during the 2 years of this field experiment.
3.5. Plant biomass
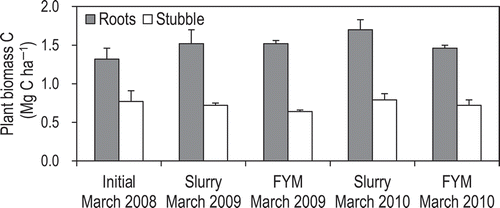
The amount of root C was 1.7–2.4 times that of the stubble C (). There were no significant interannual differences in the amount of root C (P = 0.638 for both treatments) or stubble C (P = 0.165 for both treatments) in the slurry or FYM plots. Thus, the amounts of root C and stubble C in both plots appear to have remained unchanged during the 2 years of this field experiment. Therefore, we did not account for either in our calculation of the C balance of the grassland (Section 3.6).
Figure 6 Carbon contents in the roots and stubble. Values are means ± SEM (n = 3). FYM, farmyard manure.
3.6. Carbon balance of the grassland
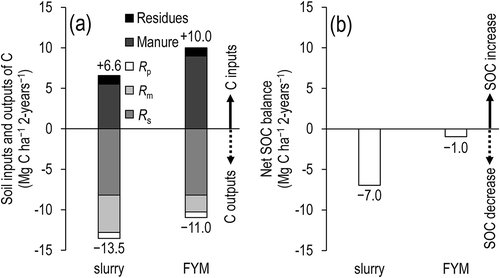
The C balance of the grassland, calculated as C inputs (manure + plant residues) minus C outputs (Rs + Rm + Rp), was –7.0 Mg-C ha–1 per 2 years in the slurry plots and –1.0 Mg-C ha–1 per 2 years in the FYM plots (, ). Thus, FYM application produced a smaller decrease in SOC by 6.0 Mg-C ha–1 per 2 years in comparison with the slurry application.
Figure 7 (a) Soil inputs and outputs of C to the soil and (b) net SOC balance. FYM, farmyard manure; Rp, decomposition of plant residues; Rm, decomposition of manure; Rs, decomposition of soil organic matter; SOC, soil organic carbon.
4. DISCUSSION
The organic C input in the FYM plots was greater than that in the slurry plots (), but Rm in the FYM plots was smaller than that in the slurry plots (). Consequently, a substantially greater amount of FYM C than slurry C remained in the soil (), suggesting that the conditions under which manure was stored at the farm was an important factor that controlled the resulting C balance (). On the one hand, readily available organic C in the slurry, which had been stored under relatively anaerobic conditions, was quickly decomposed after its application to the grassland (). On the other hand, that in FYM, which had been stored under relatively aerobic conditions, could have decomposed before its application to the grassland (i.e., during the composting process). In addition, sawdust was applied during the composting of FYM, and the sawdust contains lignin, which decomposes more slowly than other components derived from dead vegetation.
Manure and synthetic fertilizer applications indirectly increase the C input to grassland by stimulating plant growth (Ryals and Silver Citation2013). In this study, a large part of the aboveground primary production (i.e., more than 80%) was harvested (). Consequently, the C input through plant residues () was much smaller than that through manure (). The amount of newer plant residues remaining was greater than that of older plant residues (), but 68–70% of plant residues had decomposed after 2 years, suggesting that plant residues contributed less than FYM to the C balance (i.e., the change in SOC; ). The annual rate of decomposition of the plant residues (74% year–1, ) was comparable to that of a mixture of improved pasture species (~70% year–1), but larger than that of Japanese silver grass (Miscanthus sinensis; ~40% year–1) owing to differences in the composition of the OM (i.e., more crude fiber in the Japanese silver grass) in the plant residues (Kirita et al. Citation2002).
There was no significant difference in annual yield between the slurry and FYM plots (), suggesting that fertility management in both treatments worked acceptably well (). From a different perspective, this also suggests that the growth and turnover rates of roots were similar in the slurry and FYM plots. Root-derived C is considered to be retained in soils more efficiently than leaf-derived C (Schmidt et al. Citation2011; Soussana et al. Citation2010). Fine roots are the most dynamic portion of a plant’s root system and a major source of SOM (Solly et al. Citation2013), however, belowground C cycling remains unclear. In our calculation of the C balance, we did not account for the input of organic C to the soil through root turnover, so the C balance (i.e., the SOC increase) could have been underestimated in both treatments (). However, the input of organic C to the soil through root turnover is likely to be similar in both treatments.
Rs (4.0–4.2 Mg-C ha–1 year–1; ) was comparable to the values previously measured in Japanese grassland (3.6–3.9 Mg-C ha–1 year–1; Shimizu et al. Citation2009). We measured Rs by using the root-exclusion approach (Hanson et al. Citation2000). However, this approach disturbs the soil aggregates, exposing more of the protected SOM to microbial decomposition. In addition, direct impacts of raindrops on the soil surface in the absence of protection from plants could promote physical disruption of the soil aggregates, also stimulating decomposition of SOM (Six et al. Citation2004). Therefore, we may have overestimated Rs in comparison with the values for an intact grassland soil (). However, direct measurement of Rs of intact grassland soil is confounded by the effects of plant growth, so it is difficult to solve this problem.
Despite this problem, it was possible to compare the C balances of the slurry and FYM plots because the organic C input to the soil through root turnover can be considered to be similar in the two treatments, so we assumed identical Rs in the two treatments (). In addition, the annualized C balances for the slurry plots (–3.5 Mg-C ha–1 year–1; ) and FYM plots (–0.5 Mg-C ha–1 year–1; ) were similar to previously measured values in mowed Japanese grasslands that respectively received only synthetic fertilizer (–2.9 to –3.8 Mg-C ha–1 year–1) or FYM plus synthetic fertilizer (0.0–0.3 Mg C ha–1 year–1), respectively (Shimizu et al. Citation2009). These results collectively suggest that slurry C had limited capacity for maintaining SOC in comparison with FYM C (Angers et al. Citation2010).
A literature search showed that grassland C sequestration reaches on average 0.05 Mg-C ha–1 year–1 according to inventories of SOC stocks, and 0.22 Mg-C ha–1 year–1 according to C flux balance (Soussana et al. Citation2010). The annualized C balances obtained in the present study were comparable with these values. In a cutting regime, a large part of primary production is harvested from grassland as hay or silage, however, these OM exports may be compensated for by OM imports through manure application (Soussana et al. Citation2010). Manure application in an appropriate manner is favorable management option to increase SOC and effectively managing livestock waste. (Hirata et al. Citation2013). Our results suggest that the types of manure, from different storage conditions, had significant effect on the C balance.
The amount of root biomass increased slightly but not significantly during the 2 years of the study (), and although it is possible that the additional nutrients increased root growth, statistical limitations related to the numbers and time scale of the sampling prevent us from confirming this hypothesis. The amount of roots in a relatively young grassland (i.e., less than 15 years after renovation) could increase (Whitehead et al. Citation1990), but accurate estimation of the annual change in root biomass can be obtained only from long-term field experiments with adequate replication, owing to the high temporal and spatial variability in this parameter. To complete our assessment of the C balance of the grassland, it will be necessary to perform further research on the C inputs to the soil through the growth and turnover of roots.
5. CONCLUSIONS
FYM application produced a smaller decrease in the C balance of the grassland, which we calculated as C inputs (manure + plant residues) minus C outputs (Rs + Rm + Rp); the decrease amounted to 6.0 Mg C ha–1 per 2 years less than in the treatment with slurry application. The decomposability of the applied manure therefore depended on the storage conditions, which have an important effect on the C balance (i.e., increment of SOC). Plant residues (i.e., harvest loss) contributed less than FYM to the C balance, but additional research will be required to determine the contribution of C inputs through growth and turnover of roots.
ACKNOWLEDGMENTS
This work was financially supported by the Ministry of Agriculture, Forestry and Fisheries of Japan through a research project entitled “Development of technologies for mitigation and adaptation to climate change in Agriculture, Forestry and Fisheries.” We thank Prof. Ryusuke Hatano, Hokkaido University, and Dr. Kazuyuki Yagi, National Institute for Agro-Environmental Sciences, for their valuable discussion of the experimental design.
REFERENCES
- Ammann C, Flechard CR, Leifeld J, Neftel A, Fuhrer J 2007: The carbon budget of newly established temperate grassland depends on management intensity. Agric. Ecosyst. Environ., 121, 5–20. doi:10.1016/j.agee.2006.12.002
- Angers DA, Chantigny MH, MacDonald JD, Rochette P, Côté D 2010: Differential retention of carbon, nitrogen and phosphorus in grassland soil profiles with long-term manure application. Nutr. Cycl. Agroecosyst., 86, 225–229. doi:10.1007/s10705-009-9286-3
- Angers DA, Rochette P, Chantigny MH, Lapierre H 2007: Use of 13C abundance to study short-term pig slurry decomposition in the field. Soil Biol. Biochem., 39, 1234–1237. doi:10.1016/j.soilbio.2006.12.028
- Bhogal A, Nicholson FA, Young I, Sturrock C, Whitmore AP, Chambers BJ 2011: Effects of recent and accumulated livestock manure carbon additions on soil fertility and quality. Eur. J. Soil Sci., 62, 174–181. doi:10.1111/j.1365-2389.2010.01319.x
- Conant RT 2010: Challenges and opportunities for carbon sequestration in grassland systems: a technical report on grassland management and climate change mitigation. pp. 1–57, Food and Agriculture Organization of the United Nations, Rome.
- Conant RT, Paustian K, Elliott ET 2001: Grassland management and conversion into grassland: effects on soil carbon. Ecol. Appl., 11, 343–355. doi:10.1890/1051-0761(2001)011[0343:GMACIG]2.0.CO;2
- Davidson EA, Janssens IA 2006: Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 440, 165–173. doi:10.1038/nature04514
- Diacono M, Montemurro F 2010: Long-term effects of organic amendments on soil fertility. A review. Agron. Sustain. Dev., 30, 401–422. doi:10.1051/agro/2009040
- Gregorich EG, Greer KJ, Anderson DW, Liang BC 1998: Carbon distribution and losses: erosion and deposition effects. Soil Till. Res., 47, 291–302. doi:10.1016/S0167-1987(98)00117-2
- Gregorich EG, Janzen HH 1996: Storage of soil carbon in the light fraction and macroorganic matter. In Structure and Organic Matter Storage in Agricultural Soils, Eds.Carter MR, Stewart BA, pp. 167–190. CRC Press, Boca Raton.
- Hanson PJ, Edwards NT, Garten CT, Andrews JA 2000: Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry, 48, 115–146. doi:10.1023/A:1006244819642
- Harada Y, Haga K, Osada T, Koshino M 1993: Quality of compost produced from animal wastes. Jpn. Agric. Res. Q, 26, 238−246.
- Hirata R, Miyata A, Mano M et al. 2013: Carbon dioxide exchange at four intensively managed grassland sites across different climate zones of Japan and the influence of manure application on ecosystem carbon and greenhouse gas budgets. Agr. For. Meteorol., 177, 57–68. doi:10.1016/j.agrformet.2013.04.007
- Hokkaido Prefectural Agricultural Experiment Stations and Hokkaido Animal Research Center (HPAES and HARC) 2004: Method for manure utilization. In Handbook of Animal Waste Management and Utilization in Hokkaido 2004, Eds. Ohara M, Takeda Y, Omura K, pp. 64–67. Hokkaido Prefectural Agricultural Experiment Stations and Hokkaido Animal Research Center, Sapporo. (in Japanese)
- Kirita H, Shiyomi M, Takahashi S, Yamamoto Y 2002: Relationship between decomposition of pasture plant dead materials and meteorological factors. Grassl. Sci., 48, 95–99.
- Kurashima K, Ota T, Kusaba T, Amano Y, Yamamoto K, Kimura T, Kondo H, Saito G 1993: Classification and characteristics of soils at the National Grassland Research Institute. Misc. Publ. Natl. Grassl. Res. Inst., 3, 1–47. (in Japanese)
- Lal R 2004: Soil carbon sequestration to mitigate climate change. Geoderma, 123, 1–22. doi:10.1016/j.geoderma.2004.01.032
- Matsuura S, Mori A, Hojito M, Kanno T, Sasaki H 2011: Evaluation of a portable chamber system for soil CO2 efflux measurement and the potential errors caused by internal compensation and water vapor dilution. J. Agric. Meteorol., 67, 127–137. doi:10.2480/agrmet.67.3.7
- Mori A, Hojito M 2007: Grassland renovation increases N2O emission from a volcanic grassland soil in Nasu, Japan. Soil Sci. Plant Nutr., 53, 812–818. doi:10.1111/j.1747-0765.2007.00202.x
- Mori A, Hojito M 2015: Effect of dairy manure type and supplemental synthetic fertilizer on methane and nitrous oxide emissions from a grassland in Nasu, Japan. Soil Sci. Plant Nutr., 61, 347–358. doi:10.1080/00380768.2014.981676
- National Grassland Research Institute (NGRI) 2001: Meteorological observation in National Grassland Research Institute. Misc. Publ. Natl. Grassl. Res. Inst., 4, 3–5. (in Japanese)
- Paustian K, Andrén O, Janzen HH, Lal R, Smith P, Tian G, Tiessen H, Van Noordwijk M, Woomer PL 1997: Agricultural soils as a sink to mitigate CO2 emissions. Soil Use Manage., 13, 230–244. doi:10.1111/j.1475-2743.1997.tb00594.x
- Prolingheuer N, Scharnagl B, Graf A, Vereecken H, Herbst M 2014: On the spatial variation of soil rhizospheric and heterotrophic respiration in a winter wheat stand. Agric. For. Meteorol., 195–196, 24–31. doi:10.1016/j.agrformet.2014.04.016
- Rees DVH 1982: A discussion of sources of dry matter loss during the process of haymaking. J. Agric. Engng. Res., 27, 469–479. doi:10.1016/0021-8634(82)90085-3
- Rotz CA, Muck RE 1994: Changes in forage quality during harvest and storage. In Forage Quality, Evaluation, and Utilization, Ed. Fahey GC, pp. 828–868. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison.
- Ryals R, Silver W 2013: Effects of organic matter amendments on net primary productivity and greenhouse gas emissions in annual grasslands. Ecol. Appl., 23, 46–59. doi:10.1890/12-0620.1
- Schmidt MWI, Torn MS, Abiven S et al. 2011: Persistence of soil organic matter as an ecosystem property. Nature, 478, 49–56. doi:10.1038/nature10386
- Shiga H, Ohyama N, Maeda K, Suzuki M 1985: An evaluation of different organic materials based on their decomposition pattern in paddy soils. Res. Bull. Natl. Agric. Res. Cetr., 5, 1–19. (in Japanese with English summary)
- Shimizu M, Hatano R, Arita T et al. 2014a: Farmyard manure application mitigates greenhouse gas emissions from managed grassland in Japan. In Sustainable Agroecosystems in Climate Change Mitigation, Ed. Oelbermann M, pp. 115–132. Wageningen Academic Publishers, Wageningen.
- Shimizu M, Hatano R, Arita T et al. 2014b: Mitigation effect of farmyard manure application on greenhouse gas emissions from managed grasslands in Japan. In Soil Carbon, Eds. Hartemink AE, McSweeney K, pp. 313–325. Springer International Publishing, Cham.
- Shimizu M, Marutani S, Desyatkin AR, Jin T, Hata H, Hatano R 2009: The effect of manure application on carbon dynamics and budgets in a managed grassland of Southern Hokkaido, Japan. Agric. Ecosyst. Environ., 130, 31–40. doi:10.1016/j.agee.2008.11.013
- Singh BP, De Courcelles VR, Adams MA 2011: Soil respiration in future global change scenarios. In Soil Health and Climate Change, Eds. Singh BP, Cowie AL, Chan KY, pp. 131–153. Springer Berlin Heidelberg, Berlin.
- Six J, Bossuyt H, Degryze S, Denef K 2004: A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res., 79, 7–31. doi:10.1016/j.still.2004.03.008
- Smith P, Martino D, Cai Z et al. 2008: Greenhouse gas mitigation in agriculture. Philos. Trans. R. Soc. B, 363, 789–813.
- Smith P, Powlson DS 2000: Considering manure and carbon sequestration. Science, 287, 428–429. doi:10.1126/science.287.5452.427e
- Soil Survey Staff 2014: Keys to Soil Taxonomy, 12th ed., pp. 1–360. USDA Natural Resources Conservation Service, Washington, DC.
- Solly E, Schöning I, Boch S, Müller J, Socher SA, Trumbore SE, Schrumpf M 2013: Mean age of carbon in fine roots from temperate forests and grasslands with different management. Biogeosciences, 10, 4833–4843. doi:10.5194/bg-10-4833-2013
- Soussana JF, Loiseau P, Vuichard N, Ceschia E, Balesdent J, Chevallier T, Arrouays D 2004: Carbon cycling and sequestration opportunities in temperate grasslands. Soil Use Manage., 20, 219–230. doi:10.1079/SUM2003234
- Soussana JF, Tallec T, Blanfort V 2010: Mitigating the greenhouse gas balance of ruminant production systems through carbon sequestration in grasslands. Animal, 4, 334–350.
- Tochigi Prefectural Government 2002: Fertilizer Recommendations for Agricultural Crops, pp. 57–58. Tochigi Prefectural Government, Utsunomiya. (in Japanese)
- Whitehead DC, Bristow AW, Lockyer DR 1990: Organic matter and nitrogen in the unharvested fractions of grass swards in relation to the potential for nitrate leaching after ploughing. Plant Soil, 123, 39–49.
- Zhou T, Shi P, Hui D, Luo Y 2009: Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implications for carbon-climate feedback. J Geophys. Res., 114, G02016.