
ABSTRACT
This study aims to characterize soil chemical properties and microbial biomass, greenhouse gas production, and organic matter dynamics in upland rice field as affected by the long-term fertilizer managements in Uganda. Soil total C (TC) and N (TN) contents were in the relatively smaller range under different fertilizer treatments, even after 20 crop seasons. However, available phosphate contents showed positive correlation with average yield of upland rice. Incubation experiments were conducted under aerobic or under flooding conditions to measure CO2, methane, and nitrous oxide productions. After the incubation, soil samples were extracted to quantify nitrification rate for aerobic condition and ammonification rate for flooding condition. Soil microbial biomass carbon (MBC) and nitrogen were measured. Stable isotope ratio of 13C and 15N were also determined for the soil samples. CO2 production potential under aerobic condition was higher than the flooding condition. The qCO2 (CO2/MBC) in the treatment applied with compost tended to be higher than the other treatments. Positive correlation between nitrous oxide production and nitrification was found. The delta 13C values of the soil samples indicated that the effect of C4 plants before rice cultivation still remained, while the contribution of biological N2 fixation was little according to delta 15N values. These results indicate that soil microbial biomass in upland rice field of the long-term fertilizer experiment in Uganda was characterized with higher qCO2. Greenhouse gas production was affected by fertilizer management, while soil organic C before the long-term experiment still remained in the experiment.
1. Introduction
Rice is an important staple crop, not only in monsoon Asia, but also in sub-Saharan Africa. In Africa where the increasing risk of food shortage is one of the urgent issues to be solved, the cultivation of ‘NERICA’ (New Rice for Africa) is increasing recently (Sekiya et al. Citation2013), which has benefits for high yield, pest and drought resistance, shorter growth period than conventional rice varieties, and suitable for both upland and lowland conditions (Tsuboi et al. Citation2018). However, there are few studies on soil fertility for upland rice cultivation, especially by long-term fertilizer experiment in Africa, and it is still not clear how the soil fertility can be maintained, how the sustainable rice production can be achieved, and how the environmental impact such as greenhouse gas produced under upland conditions influenced by soil managements, especially fertilization, affect the microbial properties and activities in the soil under such upland rice cultivation. In this study, we investigated how the chemical, biochemical properties, and greenhouse gas production of the upland rice soils are affected by the long-term fertilizer experiment field in Uganda cropped with NERICA rice with incubation experiment to consider sustainable soil managements and effect on nutrient dynamics. Stable isotope ratio of 13C and 15N of the soil organic matter were also determined to elucidate the long-term dynamics of soil organic matter and nutrients. Natural abundance of 13C in soils usually reflect that of the source of C in soils. It is widely known that C4 plant species have heavier C as a result of C4 photosynthesis as compared with C3 plants (Yoneyama et al. Citation2001; Yoneyama, Okada, and Ando Citation2010). In this study, we aimed to collect information that tell us how the soils in the NERICA rice experiment were formed (affected more by C3 or by C4 plants). As well as 13C, natural abundance of 15N in soils give us information on how N was added in the soils. For example, N that are fixed from the air through biological N fixing activities usually show around 0‰ of δ15N. On the other hand, N that are processed relatively well show a higher δ15N. Well decomposed manures and soils that are affected by these materials usually have high δ15N reflecting microbial N processes. Our hypothesis was that soils in the NERICA experiment show around 0‰ of δ15N because N fixers living in NERICA roots affect the N isotopic ratio in soils.
2. Materials and methods
2.1. Site description
Upland rice soils (Acric Ferralsols, with sandy clay loam texture) were sampled from long-term fertilizer experiment field in National Crop Research Institute (NaCRRI), Namulonge, Uganda (00º30ʹ46.4N, 32º38ʹ03.6E, 1120 m above sea level, air temperature as 23°C, and precipitation as 1225 mm). The fertilizer treatments were maintained as a combination of N, P, and K and compost as Control, N, NP, PK, NK, NPK, NPK Compost, and Compost, each having duplicated plots, cultivated for double rice (NERICA-4, from Asian type (Oryza sativa L) x African type (Oryza glaberrima Steud)) cropping in each year since 2004. Before the experiment, the site area was covered mainly by wild grasses (mainly Poaceae). For plots with N, 60 kg N ha−1 in urea was applied in three equal splits, as basal fertilizer, at 30 days after sowing, and at the panicle initiation stage. P (triple super phosphate) and K (muriate of potash) were applied as basal fertilizer at a rate of 30 kg ha−1 each. Compost, which was made of rice straw composted for one season (composed of organic matters 23.2%, N 0.96%, P 1.14%, K 0.05%, Ca 0.58%, and Mg 0.17%), was applied as basal at a rate of 5 t ha−1. Treatments were block randomized with duplicate plot and their yield data were described by Tsuboi et al. (Citation2018).
2.2. Soil sampling and analysis
Surface soil (0–10 cm) was sampled from three portions between rice plant rows from each replicate and mixed together for each replicate before harvest in December 2013 after 17 crop seasons excluding three seasons without harvest due to severe drought, then partially dried to pass through a 2-mm sieve and their chemical properties were measured in the composite soil samples with each replicate, for soil moisture, pH (H2O), electric conductivity (EC; soil:water ratio 1:5), total carbon (TC), total nitrogen (TN) using a C/N analyzer (MT 700 with an auto-sampler MTA 600, Yanaco, Japan), permanganate oxidizable carbon (POXC) by digestion-titration method (Tirol-Padre et al. Citation2007), available P by Truog and Bray II methods (Dojo Yobun Bunseki hou). Soil microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) were measured by chloroform-fumigation-extraction method (Inubushi, Brookes, and Jenkinson Citation1991), 1 week for pre-incubation after sieving. Soluble organic C in 0.5 M K2SO4 extraction from unfumigated soil was determined by total organic carbon analyzer (TOC 5000, Shimadzu, Kyoto, Japan). All analyses were carried out in duplicate and indicated as average and difference between duplicated plot samples’ data.
2.3. Incubation
The incubation experiments were conducted with the soil under aerobic condition or under flooding condition, both for 28 days, starting 1 week for pre-incubation after sieving. Ten gram of moist soil was put into 100 mL glass bottle and headspace was replaced with N2 for flooded incubation, while no gas exchange but sealed with rubber stopper and placed in the incubators either at 25°C for aerobic at 60% water-holding capacity, or 30°C for flooded incubation. Gas (CO2, CH4, and N2O) productions in the headspace were measured weekly by gas chromatography (Shimadzu, GC 14B, Kyoto, Japan) equipped with a thermal conductivity detector (TCD), a flame iodized detector (FID), and an electron capture detector (ECD) for CO2, CH4, and N2O, respectively. After gas measurement, headspace gas was exchanged to fresh air or N2, for aerobic and anaerobic incubation, respectively. Respiration quotient (qCO2) and ratio of biomass C to N were calculated as CO2/MBC and MBC/MBN, respectively. After the incubation, soil samples were extracted with 10% KCl solution to determine nitrate (NO3−) and ammonium (NH4+) contents by the hydrazine reduction (Hayashi, Sakamoto, and Yoshida Citation1997) and nitroprusside (Anderson and Ingram Citation1989) methods, respectively, and nitrification rate for aerobic condition and ammonification rate for flooding condition were calculated as increase of NO3– and NH4+ contents during incubation period.
2.4. Stable isotopic analysis
The stable isotopic composition of carbon (δ13C) in ‰ = [(13C/12C of soil)/(13C/12C of standard)–1] × 1000 and nitrogen (δ15N) in ‰ = [(15N/14N of soil)/(15N/14N of standard)–1] × 1000 was determined using an isotope-ratio mass spectrometer (IRMS) connected to an elemental analyzer (Integra-CN, PDZ Europa, UK). The reproducibility of the measurements of δ13C and δ15N checked using a working reference material was >0.1‰ and >0.2‰, respectively. The working reference material, glycine (δ13C: −31.9‰ ± 0.1‰; δ15N: 2.0‰ ± 0.1‰), was calibrated against NBS19 for δ13C and against IAEA-N1 for δ15N. We analyzed stable isotopic composition of soils after air-drying process and enclosing sample into a tin capsule.
3. Results
3.1. Soil chemical properties
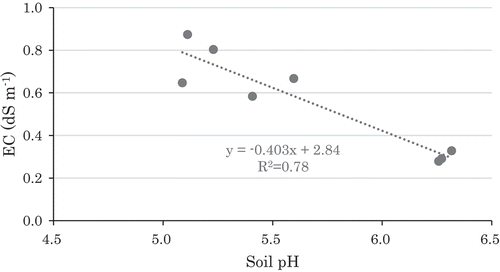
Soil pH was in the range of 5.1–6.3, higher in control, NK, and compost-amended soil (). Soil pH in NPK Compost was higher than NPK treatment. Soil EC was in the range of 0.3–0.9 dS m−1, higher in NPK, NP, PK, and NPK Compost-amended soil. Soil total C (TC) and N (TN) contents were in the range of 1.32–1.53% and 0.12–0.16%, respectively, and soil C/N of 9.7–11.0 (). Contents of NO3– were in the range of 0.62–11.2, which was much larger range than that of NH4+ as 3.32–4.03 mg N kg−1. NO3– in N, NP, PK, NPK, and NPK Compost were higher than the other treatments. Soluble organic C was in the range of 70–143 mg C kg−1, while POXC was in the range of 338–434 mg C kg−1 (). Soluble organic C in N, PK, and NPK were higher than the other treatments, while POXC in compost-applied treatments was higher than the other treatments, even though TC in all treatments was almost the same. There was a negative relationship between soil pH and EC in each treatment (), and a positive relationship between EC and NO3– contents. Available P was higher in NP, NPK, and NPK Compost, especially by Bray II and Truog methods, while the correlation to yield data will be described later.
Table 1. Chemical properties of examined soils.
Figure 1. Soil pH and EC.
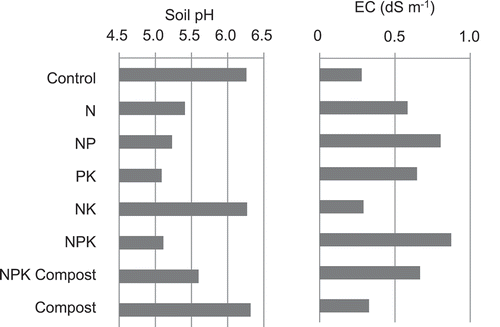
Figure 2. Correlation between soil pH and EC.
3.2. Soil microbial properties, N transformation, and greenhouse gas production
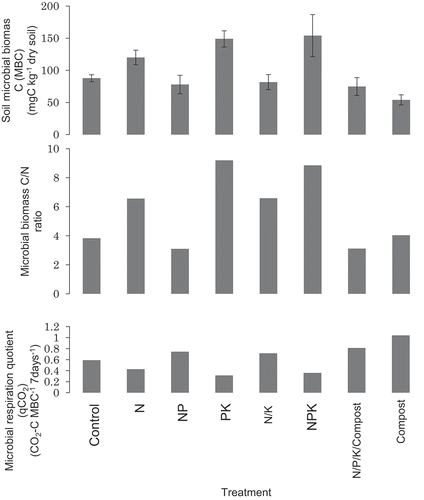
Microbial biomass C was in the range of 54–153 mgC kg−1, higher in NPK, PK, and N amended soil () and biomass N in 12–25 mgN kg−1 (data not shown). The biomass C/N ratio in those soils was lower than 10, indicating that the biomass consisted of bacteria mainly in the examined upland rice soil.
Figure 3. Microbial properties of examined soils.
As a result of incubation experiment, CO2 production increased almost lineally and reached higher after 4 weeks incubation (180–250 mgC kg−1) under aerobic condition than flooding condition (62–82 mgC kg−1) (), so that qCO2 was higher under aerobic condition. There was a tendency that qCO2 in compost-applied treatment was higher than the other treatments. Methane (CH4) production increased almost linearly and reached after 4 weeks in the range of 120–240 ug kg−1 and tended to be higher under flooded condition than aerobic condition. It seemed that CH4 production under flooded condition was similar as that under aerobic condition among the treatments, except that CH4 production under flooded condition in control treatment was lower than other treatments.
Figure 4. Greenhouse gas production during aerobic (upper column) or flooded (lower column) incubation.
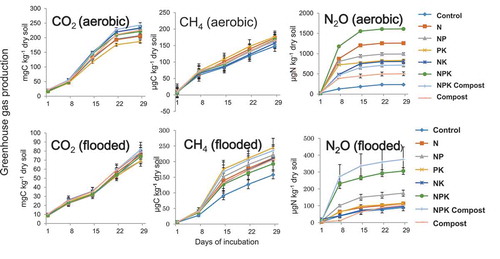
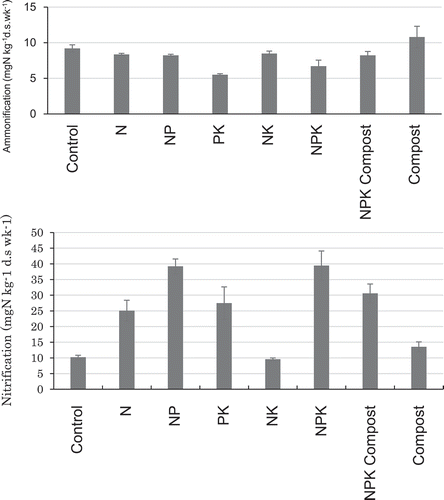
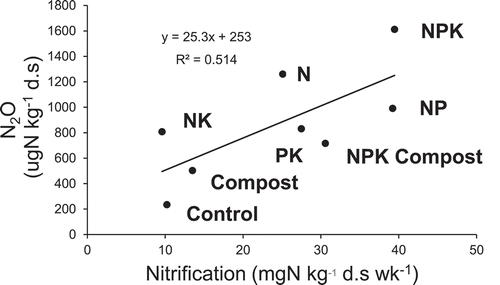
In case of N2O production, increase in the first 1 week was faster than in the following weeks (). The difference among treatment in N2O production was wider than in CO2 and CH4 productions. Particularly, the production under aerobic condition in NPK treatment was the highest, while control treatment was the lowest. Similarly, the production under flooded condition in NPK Compost treatment was the highest, while PK, NK, and control treatments were the lowest. Ammonification was in the narrower range of 5.5–10.8 mg kg−1 (). On the other hand, nitrification was in the wider range of 10–39 mgN kg−1.
Figure 5. Ammonification under flooded condition and nitrification under aerobic condition.
3.3. Stable isotope ratio of soil organic matter
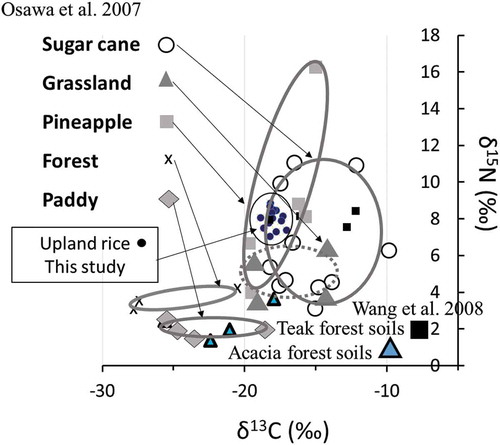
Stable isotope ratio of 13C and 15N were similar to each other for the soil samples examined, and they did not show significant differences depending on the treatments. We made a diagram to compare our results with a wider range of the data (), by citing study of Osawa et al. (Citation2007) with data of Wang et al. (Citation2008) (a study for paddy soils, grassland soils, pineapple soils, and sugar cane soils in Ishigaki island, Okinawa, subtropical Japan, and also acacia and teak forest soils in Botswana and Zambia). The δ13C values of the soil samples in NERICA study were from −17.7‰ to −19.0‰. These values were close to those soils from sugar cane and pineapple fields in Okinawa area. On the other hand, the δ15N values of the soil samples in NERICA study were from 6.67‰ to 7.79 ‰. Both δ13C and δ15N values in soils were not significantly affected by fertilization treatments.
Figure 6. Stable isotope ratio of soil organic matter in Uganda upland rice soil to compare with other ecosystems.
4. Discussion
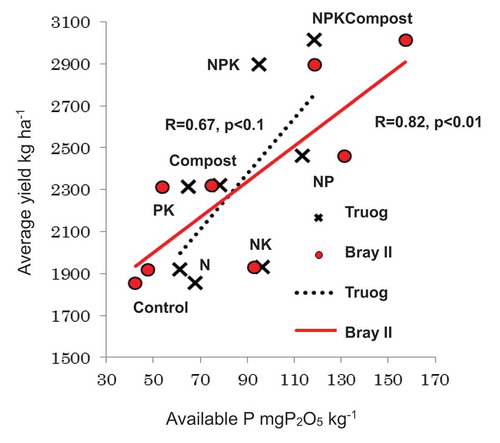
Fertilization would generally decrease soil pH as described by Bloom (Citation2000), also in this study in N, NP, PK, and NPK-treated soils (), but these values were not as low as critical level of 4.2 (Sanchez Citation1976). Higher soil pH in compost-amended soil indicated that compost might be a countermeasure to avoid lowing soil pH, but not clear why pH in NK did not decrease. Fertilization would generally increase soil EC, but these values were not as high as critical level of 1.5 dS m−1 (Keren Citation2000). Higher soil EC in NPK, NP, PK, and NPK Compost-amended soil was also related to nitrification (), as described later. On the other hand, higher pH and lower EC in NK-treated soil were also related to lower nitrification. From present results, soil total C and N contents were not much influenced by long-term fertilizer treatment (), which is the same finding as Tsuboi et al. (Citation2018), indicating that soil organic matter would not be a critical factor for soil fertility, while soil available P and precipitation could be more critical factors for rainfed upland rice cultivation. Available P was significantly positively correlated with upland rice yield (Tsuboi et al. Citation2018) ().
Figure 7. Correlation between available P and average yield of upland rice.
It was also indicated that the microbial biomass C and biomass C/N ratio in upland rice soil in the Uganda soil were lower than the upland soils, except sandy soil, in Japan (Shibahara and Inubushi Citation1995), and that microbial biomass C in upland rice soil in the Uganda soil were lower than upland soils, except sandy soil, in Japan (Sakamoto and Hodono Citation2000). Higher qCO2 under aerobic incubation condition were reflected to aerobic upland condition, similar to actual field condition for the upland rice (). It was also affected much more by compost than by other fertilizer managements, as indicated especially by nitrogen fertilization, which was lowering qCO2 probably due to lowering stress on microbial biomass, similar as in long-term rice-wheat experiments in Kyushu, southern Japan (Tirol-Padre et al. Citation2005).
Higher biomass C was expected in compost-amended soil, but it was similar or even lower than control soil in this investigation. Microbial biomass and soil quality were examined in long-term rice-wheat experiments in Kyushu, southern Japan, to compare with Indian rice-wheat experiments (Tirol-Padre et al. Citation2005, Citation2007). Based on these examinations, organic manure is recognized as being essential for maintaining sustainable soil quality in those humid agroecosystems, while in water-limited upland condition like this investigation, NPK, especially P, would be more important to increase microbial biomass and decrease qCO2. Furthermore, balance of organic and inorganic matter is important under specific climate condition such as temperature and moisture as investigated by Azmal et al. (Citation1996), so the amount of organic manures needed to supplement inorganic fertilizers must be optimized to increase C and N accumulation in the soil without negative effects on crop yield. These differences to Uganda upland soil should be investigated further.
From our investigation, it was also indicated that greenhouse gas production in Uganda upland rice soil, especially CH4 production, was quite different, several order magnitudes lower than Japanese paddy soils (Inubushi et al. Citation1997). Dubey et al. (Citation2014) investigated CH4 production potential in tropical irrigated Indian paddy soils taken from five different soil orders and range from 178.4 to 431.2 mg CH4 kg−1 dry soil, which were again three order magnitudes higher than Uganda upland rice soil, which are reflected to anaerobic irrigated lowland rice condition, to compare with upland condition, or simply measured background CH4 concentration of headspace in incubation tubes.
The N2O production was more active under aerobic condition than flooded condition, which was opposite from Andosol in Japan (Inubushi, Naganuma, and Kitahara Citation1996), probably due to the difference of soil structure influencing nitrification and denitrification processes. From our study, nitrification was also positively correlated with average yield of upland rice in the long-term experiment (Tsuboi et al. Citation2018) (), indicating that soil microbial activities response to rice growth and yield more critically than soil total C and N. Nitrification was in positive correlation also with N2O production (). Significant positive correlation between N2O production and nitrification indicates that the process of nitrification is predominant in N2O production, rather than denitrification, presumably due to low available carbon contents in these upland soils, influencing nitrogen availability and upland rice yield.
Figure 8. Correlation between nitrification and average yield of upland rice in the experiment.
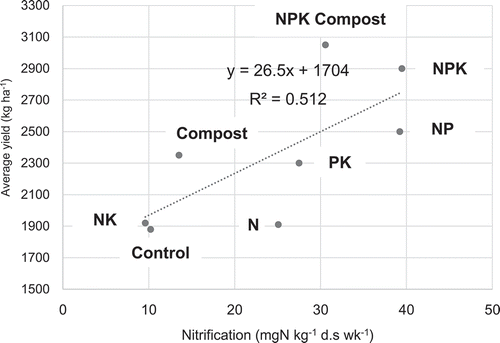
Figure 9. Relation between nitrification and N2O production after 4 weeks incubation.
The application of N-fertilizer mitigated the adverse effect of continuous cropping for panicles and spikelets but aggravated it for the rate of grain filling (Gf) and 1000 grain weight (Gwt) (Tsuboi et al. Citation2018). However, to consider that N2O has 300 times higher global warming potential than CO2 (IPCC Citation2013), it would be important to manage these upland soil environmental-friendly for the upland rice. Tsuboi et al. (Citation2018) also mentioned that among the soil properties analyzed by Uganda side, the most peculiar was N-content; after 17 seasons of continuous cropping, N content differed very little among the treatments, which is similar to this study. They considered that one possible explanation is that some nitrogen-fixing mechanisms were present. If the main source of N in the NERICA soils is N fixed through biological fixation, soils may have shown around 0‰ of δ15N. On the contrary to our hypothesis, our data of natural abundance of 15N in soils showed insignificant contribution of biological N2 fixation. The treatment also had no effect on 15N in soils. The values of δ13C in the soils indicated that the soil organic C are mainly from C4 plants, probably before the NERICA planting started. So, both of C and N in the soils may be still largely affected by the previous management (not the long-term experiment). Long-term C and N dynamics in soil organic matter still remain unclear to understand the mechanisms to support sustainable soil fertilities.
Acknowledgments
This is a part of NARO/JICA (National Agricultural Research Organization of Uganda and Japan International Cooperation Agency) project and kind support of coordinator, JICA expert, Dr. Masao Kikuchi, Prof. Emer. Chiba University, is appreciated.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Anderson, J. M., and J. S. I. Ingram. 1989. “Colometric Determination of Ammonium.” In Tropical Soil Biology and Fertility. ISSS (Ed.), 42–43. Wallingford, CT: CAB International.
- Azmal, A. K. M., T. Marumoto, H. Shindo, and M. Nishiyama. 1996. “Mineralization and Microbial Biomass Formation in Upland Soil Amended with Some Tropical Plant Residues at Different Temperature.” Soil Science and Plant Nutrition 42 (3): 463–473. doi:10.1080/00380768.1996.10416315.
- Bloom, P. R. 2000. “Soil pH and pH Buffering.” In Handbook of Soil Science, edited by M. E. Summer, B-333–350. Boca Raton: CRC Press.
- Dubey, S. K., A. Singh, T. Watanabe, S. Asakawa, A. Singla, H. Arai, and K. Inubushi. 2014. “Methane Production Potential and Methanogenic Archaeal Community Structure in Tropical Dryland Irrigated Indian Paddy Soils.” Biology and Fertility of Soils 50: 369–379. doi:10.1007/s00374-013-0858-7.
- Hayashi, A., K. Sakamoto, and T. Yoshida. 1997. “A Rapid Method for Determination of Nitrate in Soil by Hydrazine Reduction Procedure.” Japanese Journal of Soil Science and Plant Nutrition 68: 322–326. (in Japanese with English summary).
- Inubushi, K., P. C. Brookes, and D. S. Jenkinson. 1991. “Soil Microbial Biomass C, N and ninhydrin-N in Aerobic and Anaerobic Soils Measured by the Fumigation-extraction Method.” Soil Biology and Biochemistry 23: 737–741. doi:10.1016/0038-0717(91)90143-8.
- Inubushi, K., K. Hori, S. Matsumoto, and H. Wada. 1997. “Anaerobic Decomposition of Organic Carbon in Paddy Soil in Relation to Methane Emission to the Atmosphere.” Water Science and Technology 36: 523–530. doi:10.2166/wst.1997.0632.
- Inubushi, K., H. Naganuma, and S. Kitahara. 1996. “Contribution of Denitrification, Autotrophic and Heterotrophic Nitrification in Nitrous Oxide Production in Andosols.” Biology and Fertility of Soils 23: 292-298. doi:10.1007/BF00335957.
- IPCC. 2013. “The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change.” The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press.
- Keren, R. 2000. “Handbook of Soil Science.” In Salinity, edited by M. E. Summer, G-3–25. Boca Raton: CRC Press.
- Osawa, K., T. Kanda, S. Ikeda, and K. Noda. 2007. “Sediment and Nutrients Movements in Watershed Using Stable Isotopes.” Proceedings of Agricultural and Rural Engineering 55: 700–701. doi:10.2183/pjab.85.374.
- Sakamoto, K., and N. Hodono. 2000. “Turnover Time of Microbial Biomass Carbon in Japanese Upland Soils with Different Textures.” Soil Science and Plant Nutrition 46 (2): 483–490. doi:10.1080/00380768.2000.10408801.
- Sanchez, P. A. 1976. Properties and Management of Soils in the Tropics, 618. New York: Wiley.
- Sekiya, N., K. J. Khatib, S. M. Makame, M. Tomitaka, N. Oizumi, and H. Araki. 2013. “Performance of a Number of NERICA Cultivars in Zanzibar, Tanzania: Yield, Yield Components and Grain Quality.” Plant Production Science 16: 141–153. doi:10.1626/pps.16.141.
- Shibahara, F., and K. Inubushi. 1995. “Measurements of Microbial Biomass C and N in Paddy Soils by the Fumigation-extraction Method.” Soil Science and Plant Nutrition 41: 681–689. doi:10.1080/00380768.1995.10417018.
- Tirol-Padre, A., J. K. Ladha, A. P. Regmi, A. L. Bhandari, and K. Inubushi. 2007. “Organic Amendments Affect Soil Parameters in Two Long-term Rice-wheat Experiments.” Soil Science Society of America Journal 71: 442–452. doi:10.2136/sssaj2006.0141.
- Tirol-Padre, A., K. Tsuchiya, K. Inubushi, and J. K. Ladha. 2005. “Enhancing Soil Quality through Residue Management in a Rice-wheat System in Fukuoka, Japan.” Soil Science and Plant Nutrition 51: 849–860. doi:10.1111/j.1747-0765.2005.tb00120.x.
- Tsuboi, T., G. Asea, A. Maruyama, S. Matsumoto, K. Miyamoto, A. Goto, E. Justin, et al. 2018. “Effects of Rainfall, Continuous Cropping, and Fertilizer on Yield and Yield Components of an Upland NERICA Variety: A Statistical Analysis of A Long-term Field Experiment in Uganda.” Tropical Agriculture and Development 62: 78–93. doi:10.11248/jsta.62.78.
- Wang, L., P. D’Odorico, L. Ries, and S. A. Macko. 2008. “Patterns and Implications of Plant-soil δ13C and δ15N Values in African Savanna Ecosystems.” Quaternary Research 73: 77–83. doi:10.1016/j.yqres.2008.11.004.
- Yoneyama, T., Y. Nakanishi, A. Morita, and B. C. Liyanage. 2001. “δ13C Values of Organic Carbon in Cropland and Forest Soils in Japan.” Soil Science and Plant Nutrition 47: 17–26. doi:10.1080/00380768.2001.10408364.
- Yoneyama, T., H. Okada, and S. Ando. 2010. “Seasonal Variations in Natural 13C Abundances in C3 and C4 Plants Collected in Thailand and the Philippines.” Soil Science and Plant Nutrition 56: 422–426. doi:10.1111/j.1747-0765.2010.00477.x.