
ABSTRACT
This work focused on the amounts and distribution of organic C (OC), organic phosphorus (Po), and acid phosphomonoesterase (AcP) and β-glucosidase (βG) activities in volcanic soils, as well as on the effect of inorganic P (Pi) on potential P-acquiring effort. The OC and Po contents are correlated with the active aluminum (Alox) and iron (Feox) and with the stratified gradients of the enzyme activities. The AcP activity does not seem to be suppressed by soil Pi content, showing a positive linear relationship with Pi. High values for the AcP:βG activity ratio are observed, which suggests a high energy allocation (C consumption) to ensure P demand. With the increase of Alox and Feox, the AcP:βG ratios increase and the potential C:P acquisition (ln(βG):ln(AcP)) values decrease. Nevertheless, the average C:P molar ratio of the six soils (115) are below the critical value, above which soils may present low P bioavailability. The results suggest that the effort for P acquisition (AcP:βG) and the potential C:P acquisition are both affected by the size of the Pi pools and by Alox and Feox contents. If Pi retention increases due to high contents of Alox and Feox, energy allocation shifts toward AcP production, in detriment of microbial population and plant nutrition. Considering the OC accumulation in the studied soils, several hypotheses may be considered such as the energy demand toward P acquisition, and the ability of Alox and Feox metal complexes to protect organic molecules to depress βG activity.
1. Introduction
The recovering process of key nutrients from residue inputs and accumulated organic matter (OM) is an essential microbial function in soils. Generally, these fluxes require the activity of extracellular enzymes to process the more complex compounds of OM into available subunits (Caldwell Citation2005). Since enzyme activities can be reliably determined in laboratory essays, have microbial ecological significance, are sensitive to environmental stress, and respond rapidly to changes in land management, the use of soil extracellular enzymes, as biological indicators is of great interest (Dick Citation1997). Nevertheless, laboratory essays measure the potential activity of the enzymes (Vmax) and not their actual activities, since the conditions in the laboratory (pH, temperature, substrate concentration, and diffusion coefficients) are different from those occurring in situ (Nannipieri, Kandeler, and Ruggiero Citation2002).
Andosols are known to display unique morphological, physical, and chemical properties attributed to the components of their amorphous fraction, like allophane, imogolite, ferrihydrite, and Al/Fe–humus complexes (Shoji, Nanzyo, and Dahlgren Citation1993), combined with a high capacity to adsorb P, and may affect their agronomic use. Volcanic ash soils usually also contain high amounts of OM (Batjes Citation1996). Although comprising only 0.84% of the global land area, these soils store approximately 5.0 % of the global soil C (Eswaran, Berg, and Reich Citation1993; Dahlgren, Saigusa, and Ugolini Citation2004). The hypotheses that can explain OM accumulations in Andosols are: (i) P is a rate-limiting factor for OM mineralization (Buurman, Peterse, and Almendros Citation2007) attributed to the high P-fixing capacity of these soils; (ii) biodegradation of OM is limited by its complexation with Fe, Al, and allophanic constituents (Matus et al. Citation2014); (iii) enzymes and microbial by-products may be deactivated by adsorption on short-range order mineral surfaces of andic soils (Saggar et al. Citation1994); (iv) the microbial decomposition of OM may be limited by low pH values and Al toxicity (Tate and Theng Citation1980) although the formation of metal-humus complexes leaves little or no exchangeable Al available to limit microbes and plants in Andosols (Shoji and Masui Citation1972).
OM fuels the metabolic activity beneath soil surface (Bauer and Black Citation1994) and is an important indicator of soil quality and productivity (Reeves Citation1997). The potential activities of extracellular enzymes are important indicators of nutrient availabilities in soils. Since the synthesis of extracellular glucosidases and phosphatases represents a large metabolic investment, their production is induced when labile C and/or Pi, respectively, are in short supply (Zheng et al. Citation2015). Consequently, enzyme production and release from cells can be repressed or induced by genetic expression according to environmental signals (Sinsabaugh and Shah Citation2012). Soil P concentrations regulate the phosphatase activity through Pi signal transduction pathway (PHO), a mechanism linked to cellular P homeostasis (Dick, Dos-Santos, and Meyer-Fernandes Citation2011). Phosphatase synthesis is regulated throughout the growth phase of microbial cells and population, being influenced by exogenous and endogenous P pools (Persson et al. Citation2003). Phosphatase activity is repressed in non-limiting P medium, but once exogenous P content became deficient, phosphatase mRNA and phosphatases synthesis are induced (Dick, Dos-Santos, and Meyer-Fernandes Citation2011).
The exoenzyme β-glucosidase (βG) plays a major role in degradation of carbohydrates in soils and its synthesis is induced by the products of cellulose breakdown, including cellobiose, glucose, and their metabolites (Stewart and Leatherwood Citation1976). Research has shown that βG is the most abundant and easily detectable of the three enzymes (endo-1,4-glucanase, exo-1,4-glucanase and β-D-glucosidase) involved in cellulose degradation in soil, and is rarely substrate limited, thus making it ideal to examine the importance of physico-chemical controls on the turnover of SOM (Turner et al. Citation2002). Therefore, βG activity has been used to closely monitor rapid changes in soil OC associated with management practices (Bandick and Dick Citation1999). On the other hand, βG contributes to provide energy for soil microorganisms and is directly related to soil OC content (Eivazi and Tabatabai Citation1988; Bandick and Dick Citation1999). This enzyme catalyzes the cleavage of one mole of cellobiose in two moles of glucose, regulating the supply of energy for microorganisms, which are unable to take up cellobiose directly. Alef and Nannipieri (Citation1995) referred that the cellulose breakdown is a slow process, dependent on the concentration, location and mobility of cellulose-degrading enzymes. Cellulases activities, such as βG, are susceptible to the substrate crystallinity, associated substances, and surface area, also to the environmental conditions such as pH, temperature, and water content.
Regarding phosphatases, the name includes a broad group of enzymes that catalyze the hydrolysis of both esters and anhydrides of phosphoric acid (Schmidt and Laskowsky Citation1961). Acid phosphomonoesterase and alkaline phosphomonoesterase are phosphoric monoester hydrolases with optimal activities in acid and alkaline environments, whereas phosphodiesterase is a phosphoric diester hydrolase. All these three enzymes act on ester bonds (Deng and Tabatabai Citation1997). Phosphatases play a key role in soil P cycling by catalyzing mineralization of organic P and contributing to the increasing of soil P bioavailability. Like any exoenzyme molecule, phosphomonoesterases diffuse poorly into the soil matrix (Lefebvre et al. Citation1990) and their activities are dependent on different parameters such as soil characteristics, microorganism, plant cover, inputs, and the presence of inhibitors or activators (Speir and Ross Citation1978). Generally, it is assumed that phosphomonoesterases activity is dependent on the enzymes associated with viable or lysed microbial cells, root-released enzymes into the rhizosphere and detritosphere (Nannipieri Citation1994), although root-released enzymes probably mineralize organic P from sloughed off or damaged cells rather than from native soil organic P (Lefebvre et al. Citation1990). Nevertheless, the major contribution to its activity, in soil, is due to the extracellular enzymes stabilized by colloids (Nannipieri et al. Citation2011). In volcanic soils, minerals such as allophane and ferrihydrite showed evidences of enzymes protection, namely AcP and βG and other enzymes appear to be sorbed in the soil minerals without discrimination of the enzyme type (Allison Citation2006).
Fertilization affects soil biochemical and biological properties (Liang et al. Citation2014), and the influence of external nutrient inputs on soil microbial ecology has been frequently emphasized (Ai et al. Citation2012). As an example, PO43- is a competitive inhibitor of both phosphomonoesterases in soils (Juma and Tabatabai Citation1978), although the absence of response of phosphatase activities to P addition has been also reported (Schneider et al. Citation2001). On the other hand, N fertilization generally increases the activity of phosphatases in soils (Marklein and Houlton Citation2012).
Regarding the equilibrium between C and P Sinsabaugh, Hill, and Shah (Citation2009, Citation2010) found an average value of 1.6 for the ratio of AcP to βG activities in 40 different soils with no andic characteristics. Nevertheless, in a long-term field experiment on an Andosol, Moro, Kunito, and Sato (Citation2015) observed an increase from 3.9 to 9.5 of the same ratio when no P was added or when P was limiting plant growth. Accordingly, the value of this ratio seems to reflect, even for crops, the variable readiness for acquiring phosphorus in soils with different P retention capacity or with different mineral P (or N) fertilizations.
Mineral fertilizers are widely used in the Azores region to enhance soil fertility and improve pasture production, particularly P fertilization due to the high contents of active fractions of Al and Fe and consequent high P retention capacity of the soils. Consequently, it was hypothesized that the abundant presence of Al and Fe active minerals may present a specific role on the relationships between soil OC, soil P enrichment, and the activity of free AcP and βG enzymes in these soils. Therefore, the specific aims of this work were: (i) to evaluate the stratification βG and AcP activities in the 15 cm superficial layer of six Andosols collected in Azores (Pico Island) under cattle grazed pasture and fertilization over the past 20 years, and; (ii) to study the relationships among both enzymes, OM accumulation, and Pi and (iii) assess the effect of Al and Fe active minerals on βG and AcP potential activities and on the indices of P acquisition.
2. Materials and methods
2.1. Soil classification and land management
Six soils, from commercial farms, in Pico island of the Azores archipelago, located between 200 and 700 m of altitude were selected. Soil I is a Haplic Andosol, II and III are Placic Andosol and IV, V and VI are Leptic Vitric Andosol, according to WRB (Citation2006). Brief dicriction: Vitric Andosols are characterized with lower degree of weathering (Alox + 1/2 Feox above 0.4%) and minimum concentrations of glass content (above 10%), bulk density above 0.9 kg dm−3, and PR% above 25%. Haplic and Placic Andosols are soils with higher degree of weathering than Vitric Andosols (Alox + 1/2 Feox above 2.0%) and minimum concentrations of glass content (below 10%), bulk density below 0.9 kg dm−3, and PR% above 70%. Placic Andosols tend to be highly weathered with high metal-humus content.
Soils I and VI were never tilled and the remaining ones have not been tilled for more than 20 years. All soils are under pastures use, being fertilized annually ().
Table 1. Soils management practices
2.2. Soil sampling
Superficial layers of 15 cm were selected in each location and then subdivided in 5 sub-layers (0–1, 1–2, 2–5, 5–10, and 10–15 cm) making a total of 30 soil samples. Samples were air-dried, sieved through a 2-mm screen, and stored until analysis, which were conducted in duplicate.
2.3. Soil analysis
Phosphate retention (PR, %) was assessed according to Blakemore, Searle, and Daly (Citation1987), organic carbon (OC) through near-infrared detection with an elemental analyzer (Primac SC, Skalar; Breda, NL), and pH in a 1:2.5 soil-water suspension.
Active Feox and Alox were extracted according to the 0.2 M (NH4)2(COO2)2.H2O – 0.2 M (COOH2)2.2H2O pH 3.0 method (Schwertmann Citation1964) and quantifications in the extracts were performed by atomic absorption spectrometry (Thermo Scientific ICE 3000 Series AA Spectrometer).
2.4. P characterization
Total P (Pt) was determined after ignition (550°C/2 h), by extracting 2 g of air-dried soil (1:25 soil-solution/16 h) with 0.5 M H2SO4. Inorganic P (Pi) was determined after extracting 2 g of air-dried soil (1:25 soil-solution/16 h) with 0.5 M H2SO4. Organic P (Po) was obtained by the difference between Pt and Pi, according to Saunders and Williams (Citation1955) modified by Walker and Adams (Citation1958).
Extractable inorganic P (Pi-Olsen) was determined by the Olsen method (Olsen et al. Citation1954) by shaking air-dried soil with 0.5 M NaHCO3 pH 8.5 (1:20 soil-solution/30 min). Total P Olsen (Pt-Olsen) was assessed by digesting an aliquot of the previous extract with 11 M H2SO4 and 4% (NH4)2S2O8 (USEPA Citation1982). Organic P Olsen (Po-Olsen) was obtained by the difference between Pt-Olsen and Pi-Olsen.
Soil P intensity (P-CaCl2) was determined according to Houba et al. (Citation1990), by shaking air dried soil with 0.01 M CaCl2 (1:10 soil-solution/30 min).
P concentration in the soil extracts and digestates were determined by the molybdate-ascorbic acid blue method in a segmented flow analyzer (SanPlus, Skalar, Breda, NL) after a dialysis step to prevent interference from color and solids in suspension (Coutinho Citation1996).
2.5. Potential enzymatic activity
The potential enzymatic activities of βG and AcP were assessed according to Tabatabai (Citation1994). Briefly, p-nitrophenyl-β-D-glucopyranoside was used as substrate to determine the βG activity in a modified universal buffer (pH 6.0) and incubated for 1 h at 37°C. AcP activity was determined with p-nitrophenyl-phosphate as substrate in a modified universal buffer (pH 6.5) and incubated for 1 h at 37°C. In both enzymatic activity determinations, the final p-nitrophenol (p-NP) concentration, as the reaction product, was determined by molecular absorption spectrophotometry, at 400 nm, in a segmented flow analyzer equipped with a dialysis step (SanPlus, Skalar, Breda, NL). Activities were expressed in µg p-NP g−1h−1 and converted to nmol p-NP g−1 OC h−1 for stoichiometric assessments.
2.6. Statistical analysis
Weighted means in each profile were calculated taking into consideration the different thickness of each layer. Statistical analyses (linear and logarithmical regressions, arithmetic, and weighed averages, ranges, and Pearson correlation coefficients (r)) were carried out using Analysis ToolPak and Solver supplement of Microsoft Excel®. The symbols *, ** and *** indicate significance at < 0.05, < 0.01, and < 0.001 probability (p) levels, respectively. Prior to analysis of the potentials for nutrient acquisition, exoenzymatic activities were loge transformed to normalize variance, according to the conventions of stoichiometric analyses (Sinsabaugh and Shah Citation2012).
3. Results and discussion
3.1. Soil properties
The weighted averages of some chemical parameters of the soils under study are shown in . Concerning OC, soils I and VI had the highest contents, 156 and 178 g OC kg−1, respectively. Despite having the same classification of soils IV and V (Vitric Andosols), soil VI seems to present a better preservation of its OC fraction, which may be attributed to the fact that it was never tilled, just as soil I. The no-tillage management of the soils is expected to be partially responsible for a general decrease on the OC mineralization fluxes (Reeves Citation1997). Nevertheless, and according to Eswaran, Berg, and Reich (Citation1993), soils derived from volcanic materials tend to accumulate OM, influenced by their andic properties. Inoue and Higashi (Citation1988) observed that the accumulation of OC in the upper 35 cm of andic soils has a strong linear relationship with extractable Alox and Feox. In the present study, a significant trend of OC increase was observed only with Feox (r = 0.54**, n = 30) but not with Alox. Matus et al. (Citation2014) and Kunito et al. (Citation2016) working with Andosols, pointed out that reduced availability of OM, as a result of Al and Fe binding, may protect a substantial pool of OC from microbial degradation, leading to suppress microbial mediated nutrient cycling.
Table 2. Chemical properties of the studied soils (weighed means of the five assessed layers)
In each of the individual soils, the OC contents decrease significantly along the soil profile ()) and this stratification may be due to the management of the soils, since they have been used as permanent grazing grasslands, not being tilled for the past 20 years and thus subjected to topsoil accumulation of crop residues and feces deposition. The average content of the soils was of 120 g OC kg−1 for the whole profile and of 175 g OC kg−1 in the top 0–1 cm layer.
Figure 1. Soil profile distribution of (a) organic carbon (OC) and (b) organic phosphorus (Po) contents
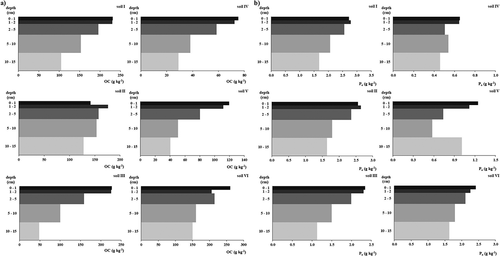
The mean value for Pt contents of the studied soils is 2.7 g P kg−1, where Po represents about 54.3 % of the total, which are in accordance with the range of 30 to 70% for the Po pool on Pt referred by Amano (Citation1981) for Andosols. Nevertheless, the extreme repartition values of 23.0% and 75.3%, observed in soils IV and I respectively, are outside of the range referred by that author. Gudmundsson, Gudmundsson, and Thorvaldsson (Citation2014) also found higher rates of Po than Pi at superficial bulk layer of 15 and 20 cm of andic soils from Iceland, after long-term inorganic fertilization of grasslands. Considering individual profiles, only soils IV and V presented higher Pi content when compared to Po, soils where the lowest contents of OC, Alox and Feox were also observed ().
The overall Po mean of the six soils was 1.5 g P kg−1 in the top 15 cm, decreasing from 2.0 to 1.3 g Po kg−1 in depth. As a component of the SOM, Po is highly correlated with OC (r = 0.91***, n = 6 and r = 0.96***, n = 30), and represents about 0.72% of the soil OM fraction. Soils I and IV had the highest and the lowest contents, 2.1 and 0.52 g Po kg−1, respectively. The lower amounts of Po and their percentages on Pt in Soils IV and V may be due to their vitric character. In Andosols from Japan, Otani and Ae (Citation1999) found that organic forms of P were tightly bound to Al and Fe compounds, thus not easily mineralizable by microorganisms, since active Al and Fe may bind labile Po in a similar way to labile Pi.
Considering that both soils IV and V have lower contents of Alox and Feox, it is expectable that the OC, and subsequently Po, also present lower values, as observed in , probably due to a faster mineralization, or due to the relative youngness of the Vitric Andosols with shorter periods of OC accumulation. Nevertheless, and despite being also a Vitric Andosol, soil VI presents a better preservation of its OC and Po contents, which may be attributed to the fact that it was never tilled, reinforcing the effect of an easier mineralization in soils IV and V. Apart from these same soils (IV and V), Po contents decrease significantly along each soil profile ()). In Otani and Ae (Citation1999, Citation1997) did not observe an accumulation or stratification of Po, even after the addition of OM, but they observed a clear stratification for Pi. Cade-Menun et al. (Citation2010) observed that labile Pi concentration was significantly higher under no-till than in tilled soils at 5–10 cm layer, after a long-term study comparing conservation tillage and conventional tillage, but did not observe significant variation in Pt and Po. Rodrigues et al. (Citation2016), studying tropical soils under conservative tillage and mineral fertilization, reported a Po built up at the surface layers. No-tillage systems led to an accumulation of Po, despite representing less than 20% of soil Pt. An increase of labile and non-labile Pi fractions was also observed, particularly in the 0–5 cm soil layers, when compared to conventional tillage. Alox + Feox contents presented positive correlations with Po (r = 0.51***, n = 30), which may indicate the protective effect of Al and Fe constituents on P mineralization (Otani and Ae Citation1997). As Alox is less correlated with Po (r = 0.48*) than Feox (r = 0.67***), the results may also suggest that Po forms in this type of soils present a higher affinity for Fe than for Al.
Regarding the Po fraction extracted by the Olsen method, soils I and V present once more the highest and the lowest Po-Olsen contents, respectively, 46 and 10 mg P kg−1 (). The top 2 cm presented an average content of 49 mg P kg−1, about 2.1 times higher than the sub-superficial layers (5–15 cm), where the rhizosphere is primarily developed and nutrients are absorbed. As observed for Po, soils IV and V do not show a clear decreasing stratification also for Po-Olsen (data not shown).
The overall mean of Po-Olsen represents nearly 1.9% of the total Po, although with variations among the samples. In fact, Po-Olsen is correlated with both Po (r = 0.95**) and OC (r = 0.91*) when the weighted means of the six soils are considered, since the accumulation of OC in the soil leads to the accumulation of Po and subsequently of potentially labile Po-Olsen. But when the relationships are searched among the individual values of the 30 samples, two linear regressions can be established between Po-Olsen and Po or OC. An expected positive slope and a high level of correlation are observed for soils I, II, III, and VI, but negative slopes are observed for soils IV and V (). In these relationships, Po presents a better correlation (r = 0.90***, n = 20) than OC (r = 0.72***, n = 20) in soils I, II, III, and IV. On the opposite, soils IV and V show a negative slope and weaker correlation for both parameters, significant with OC (r = 0.65*, n = 10) but not significant with Po.
Figure 2. Relationships between (a) organic carbon (OC) versus Olsen extractable organic phosphorus (Po-Olsen), (b) organic phosphorus (Po) versus extractable organic phosphorus (Po-Olsen)
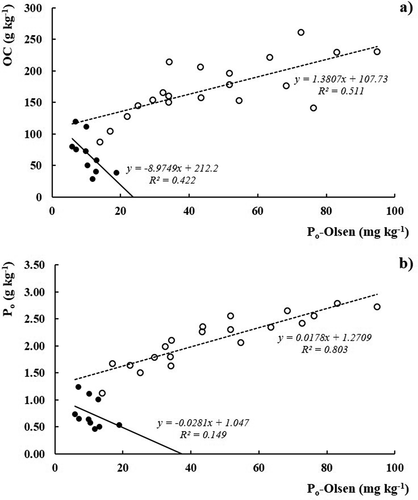
Generally, these results do not fully agree with the data presented by Turner and Haygarth (Citation2005) for temperate pastures in non-volcanic soils, since the authors did not find any correlation between Po-Olsen and soil OM, although Po-Olsen and Po were correlated (r = 0.54**). Soils with different characteristics, after disturbance, undergo changes in the native OC (Cannell and Hawes Citation1994) and the same may happen with native Po, with consequences on the Po-Olsen:Po ratio. In the present study, although this ratio does not seem to depend on the absolute content of soil OC (r = 0.35ns, n = 30), a significant relationship was observed with pH values (r = −0.63***, n = 30), with the ratio Po-Olsen:Po decreasing with soil acidity. On the other hand, whereas Po-Olsen (NaHCO3) is considered as labile form of Po and available P is closely related to these Po fractions (Tiessen, Stewart, and Cole Citation1984), its specific quantification and further study may contribute to advances in soil test procedures and interpretation. In fact, labile Po is undetectable in the molybdate-reactive P in the Olsen extract, but Po-Olsen values, in some of our samples, are higher than Pi-Olsen.
3.2. Soil enzyme activities
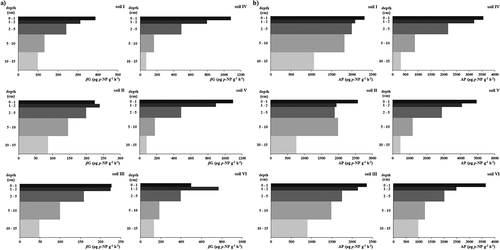
The weighted means of βG activity () vary from 111 (soil III) to 316 µg p-NP g−1 h−1 (soil V), being generally wider than the range of 48 and 169 µg p-NP g−1 h−1 reported by Dick, Breakwell, and Turco (Citation1996) for air-dried soils. The Vitric Andosols under study have a higher average βG activity, presenting approximately twice the activity (296 µg p-NP g−1 h−1) when compared to Haplic and Placic Andosols (143 µg p-NP g−1 h−1).
Table 3. Weighted average ratios of different soil parameter related with enzyme activity and P acquisition effort
Concerning the weighted means of AcP activity in each soil, values ranged from 1278 (soil IV) to 1719 µg p-NP g−1 h−1 for AcP, again in soil V. Between the two soils groups (Vitric Andosols versus Haplic and Placic Andosols), the averages of this enzyme activity are more similar than the averages of βG activity. On the other hand, the observed values tend to be at the upper limit of the range reported for air-dried soils (Dick, Breakwell, and Turco Citation1996) or temperate soils under permanent pasture (Turner and Haygarth Citation2005), 1112 and 1694 µg p-NP g−1 h−1 respectively. Borie and Zunino (Citation1983) have already reported that AcP activities are usually very high in andic soils due to: (i) protective effect of Al and Fe compounds on AcP and (ii) the low P intensity in soil solution. Later, Kunito et al. (Citation2012) also observed, in Japanese Andosols, high AcP activity, referring that microbial communities in these soils produce large amounts of phosphatases to ensure an adequate P supply.
In all the six soils, the 0–2 cm layers presented the higher activity. The gradient of the βG and AcP potential activities for each soil are shown in ) and 3(b). Along the soil profiles, their gradients show similar trends to those of OC and Po previously referred. Eivazi and Tabatabai (Citation1990), when assessing the factors that affect βG of the 100 cm superficial layer of agriculture soils in Iowa, also found a decrease in the potential activity along the soil depth. For Iowa soils, the same observation for AcP activity was reported by Juma and Tabatabai (Citation1978). In a forest andic soil, Redel et al. (Citation2008) observed higher values of AcP activity on the 0–2 cm layer (9700 to 25300 µg p-NP g−1 h−1) than in the remaining layers to a depth of 20 cm (6000 to 18200 µg p-NP g−1 h−1). Šnajdr et al. (Citation2008) also reported a gradient in the AcP and βG activities along the depth in a forest soil.
Figure 3. Soil profile distribution of (a) β-glucosidase (βG) activity and (b) acid phosphomonoesterase (AcP) activity
Although βG and AcP are highly correlated (r = 0.87***, n = 30) with each other ()), there were no correlations with OC or Po when the 30 samples were compared. However, when the linear correlations for the 5 layers of each of the individual soils were considered, relationships were much closer, with most of their r values presenting a high or very high level of significance () and 4(c)). Eivazi and Tabatabai (Citation1990) and Juma and Tabatabai (Citation1978) also found a close relationship between potential soil enzymatic activity and the respective substrate (OM and Po) in each of the studied soils.
Figure 4. Relationships between (a) acid phosphomonoesterases (AcP) versus β-glucosidase (βG); (b) β-glucosidase (βG) activity versus organic C (OC) content; and (c) acid phosphomonoesterases (AcP) versus organic P (Po) on the six studied soils
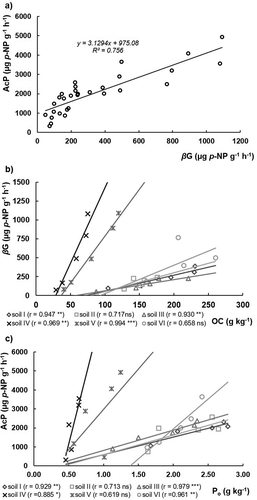
Concerning the contents of Alox and Feox () and 5(b)), inverse correlations with βG or AcP activities were observed, suggesting a repressing effect promoted by these soil minerals, as reported by Allison (Citation2006) on Hawaiian Andosols. Moreover, it seems that these soil characteristics affect more βG activity than AcP ()). In fact, both the relationship of potential enzymatic activities versus the Alox and Feox content were inverse, but the βG versus Alox + Feox was better correlated (r = 0.71***, n = 30) than the AcP versus Alox + Feox (r = 0.46*, n = 30). Yan et al. (Citation2010) in a silty clay Stagnic Anthrosol, reported that the immobilization of βG on inorganic soil colloids may have depressed its free activity until about 73%, suggesting that the active sites of the enzyme were hindered by the soil colloidal surfaces, resulting in its inaccessibility for the substrate or in modifications of the active sites. Therefore, the high contents of organic matter usually reported for Andosols (WRB Citation2006) may be also related with the reduced βG activity due to the high contents of Al and Fe of these soils. In fact, the higher slopes presented by both soils IV and V (Vitric Andosols) for the relationship between βG and OC, about 6 times higher than for other soils ()), suggest a higher substrate availability in the former soils, which also present lower OC contents as a possible consequence of their lower richness in extractable Alox and Feox ().
Figure 5. Logarithmical relationships of the enzymatic activities versus Alox and Feox content: (a) β-glucosidase (βG) and (b) acid phosphomonoesterases (AcP)
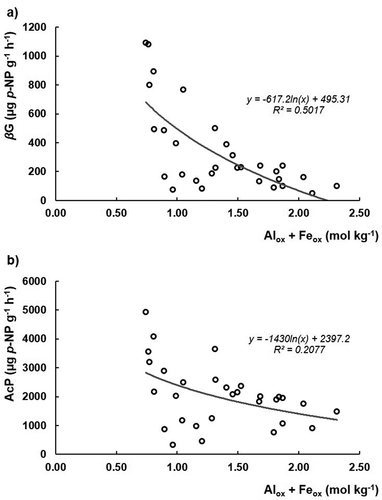
Moreover, both enzymes seem to be more affected by Alox than by Feox. βG activity presented a r = −0.65*** with Alox and r = −0.59** (n = 30) with Feox, while AcP activity presented a r = −0.42* with Alox and no significant correlation with Feox (n = 30). Kunito et al. (Citation2016) reported the repression of the carbon degrading enzymes in soils with high Alox and Feox contents. Rao and Gianfreda (Citation2000) and Rao, Violante, and Gianfreda (Citation2000), studying the interaction between soil AcP enzymatic activity with Al and Fe, observed a 45% reducing effect on its activity when interacting with Al and no effects when interacting with Fe.
The present results do not elucidate if OC accumulation, in soils derived from volcanic material is favored by: (i) the protection of soil OM by the Al and Fe compounds, as pointed out by Matus et al. (Citation2014) and Kunito et al. (Citation2016), lowering its availability for mineralization; (ii) or by the inhibition of βG expression by Al and Fe, lowering the microbial mineralization efficiency. However, it is clear, in both hypotheses, the determinant role of Al and Fe compounds on OC accumulation in the studied volcanic soils.
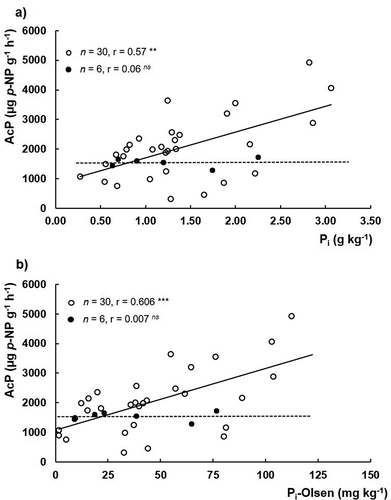
Although soils are subjected to periodic mineral P fertilization, the AcP activity seems not being inhibited by the applied P. As shown in , unexpected positive linear regressions of AcP with the total or available pools of inorganic P (Pi or Pi-Olsen respectively) are observed when all the 30 samples are considered. When the regressions are established with the weighted averages of the 6 soils, no relationship is observed. These results seem to contradict the reported inhibition of AcP expression by the presence of phosphate anion (Dick, Dos-Santos, and Meyer-Fernandes Citation2011) and the observations of Juma and Tabatabai (Citation1978), authors who reported an inhibition effect of soil AcP expression due to the presence of soil applied P. Nevertheless, this apparent contradiction may be explained by the fact that the observations made by Juma and Tabatabai (Citation1978) were done within the same soil, while the present results respect to six different soils and 30 different samples. On the other hand, the soil characteristics of both studies are substantially different, since soils derived from volcanic ashes present a high P retention capacity. Therefore, different P retention capacities of soils may overlap the inhibition of AcP expression promoted by Pi, since applied Pi in andosols can be readily fixed by Al and Fe compounds, thereby reducing its bioavailability. Once more, the lower slopes presented by Haplic and Placic Andosols for the relationship between AcP and Po, of about 7 times less than for Vitric Andosols ()), suggest a lower substrate availability (Po) in the former soils, as a possible consequence of their higher content in extractable Alox and Feox (). Otani and Ae (Citation1997) reported that labile Po in andic soils can be also fixed by Al and Fe in a similar manner to labile Pi.
Figure 6. Relationships between (a) acid phosphomonoesterases (AcP) versus inorganic P (Pi) and (b) acid phosphomonoesterases (AcP) versus extractable inorganic P (Pi-Olsen), for all samples (n = 30) and for the weighted average of each soil (n = 6)
3.3. P acquisition effort
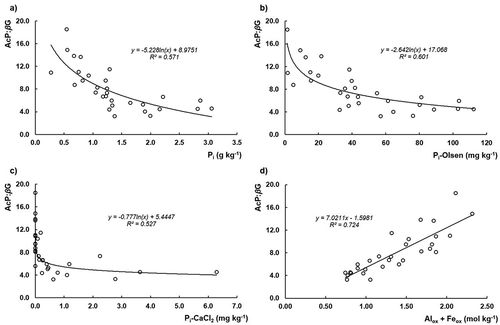
Nutrient availability in soil can be expressed by calculating the acquisition effort by microbial extracellular enzymatic activity. The ratio of P to C enzyme activities (AcP:βG), the so-called P acquisition effort, translates the energy allocation made by microorganisms to acquire P through extracellular enzymes (Moro, Kunito, and Sato Citation2015), providing information about the soil nutrient availability (Sinsabaugh and Shah Citation2012). Haplic and Placic Andosols have a higher ratio than Vitric Andosols, 12 and 5.6, respectively, showing nearly twice of the P acquisition effort (). Moro, Kunito, and Sato (Citation2015) assessed values with the same magnitude for soils derived from volcanic materials, although the values of both studies are much higher than those indicated by Sinsabaugh, Hill, and Shah (Citation2009, Citation2010) in soils that covered 40 ecosystems from the river Mississippi, Missouri, Ohio to the Great Lakes (an average of 1.6). The AcP:βG ratio presents a high inverse correlation with Pi, Pi-Olsen and P-CaCl2 (r = −0.76***, −0.78*** and −0.71***, n = 30; )), meaning that comparing soil P richness among different soils, the increase of their richness leads to the decrease on P acquisition effort, but not necessarily decreases the values of AcP activity (). These observations support the results obtained by Moro, Kunito, and Sato (Citation2015) when relating the AcP:βG ratio versus soil Pi-Olsen and the results recently presented by Fujita et al. (Citation2017) when relating AcP:βG ratio with Pi availability assessed by different methods in forest Andosols and Cambisols. But a high positive correlation ()) is observed with active Alox + Feox contents (r = 0.85***, n = 30), meaning that P acquisition effort increases with the capacity of the soil to retain P.
Figure 7. Logarithmical (ln) regression of acid phosphomonoesterases (AcP) to β-glucosidase (βG) ratio (P acquisition effort) versus: (a) inorganic P (Pi); (b) extractable inorganic P (Pi-Olsen); c) soil solution inorganic P (P-CaCl2); and (d) linear relationship of acid phosphomonoesterases (AcP) to β-glucosidase (βG) ratio versus Alox + Feox content
These results suggest that the effort for the acquisition of P decreases, through the allocation of energy to produce AcP enzymes, when available P exists at levels able to supply microorganism and plants homeostasis and full development needs. So, when Al and Fe are present at higher levels (Haplic and Placic Andosols), high values of AcP:βG ratio are expected, since P retention is higher, resulting in low Pi availability, leading to a more intense production and exudation of AcP to mineralize soil Po. In acidic Andosols from Japan, Kunito et al. (Citation2012) have already reached equivalent conclusions, observing that the high AcP activity is due to low P availability. In these soils, microbial communities produce large amounts of AcP to ensure an adequate P supply, resulting in an increase of activity in P-limited soils. Relating soil AcP:βG ratio with plant P availability and uptake, Moro, Kunito, and Sato (Citation2015) reported values between 3.9 and 9.5 for cabbage field plots subjected to P fertilization. Their results indicate the potential usefulness of this index also for agronomic purposes, since the higher values of AcP:βG were observed on plots where no P was applied, where the crop also presented the lower yield and the lower P contents in the aerial biomass.
Based on the same principles, another approach has been proposed by Sinsabaugh and Moorhead (Citation1994) and Sinsabaugh and Shah (Citation2012), using the concepts of potential activity for C acquisition and for P acquisition, ln(βG) and ln(AcP), respectively, and defining the potential of C:P acquisition as the ratio ln(βG):ln(AcP) ratio. This potential of C:P acquisition measures the different energy resource partition between Po acquisition and microbial growth (Sinsabaugh et al. Citation2008). As expected from AcP and βG relationship ()), ln(βG) versus ln(AcP) are highly correlated (r = 0.90***, n = 30), with a slope of 1.40. Studying nutrient acquisition in soils and Sinsabaugh, Hill, and Shah (Citation2009, Citation2010) indicated a slope of 1.16. Waring, Weintraub, and Sinsabaugh (Citation2014) performed a meta-analysis to explore energetic and stoichiometric constraints on enzyme production in tropical soils with low P bioavailability. These authors also found a high linear correlation (r = 0.80***, n = 57) between potential C acquisition and potential P acquisition, with a slope of 1.18. The soil type may also play a key role on the resource allocation for P-acquiring enzymes synthesis, as recently also referred by Fujita et al. (Citation2017). Therefore, the higher value achieved in our study is consistent with the low bioavailability P in soils derived from volcanic materials.
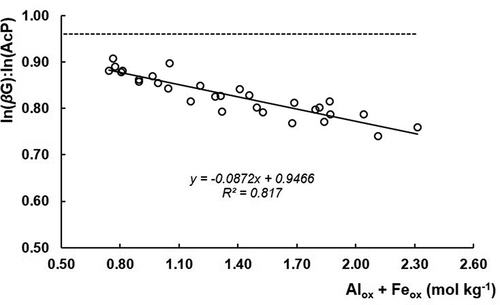
Concerning the potential of C:P acquisition (ln(βG):ln(AcP)), its value is expected to be higher when lesser energy effort is directed to P acquisition (AcP:βG), being both parameters highly correlated (r = −0.93***, n = 30) in our soils. Therefore, the potential of C:P acquisition is positively affected by the Pi pools and negiatively affected by Alox and Feox contents, in accordance with the results of the present study referred previously () and with Allison (Citation2006) and Sinsabaugh and Shah (Citation2012) observations. In the case of the regression of ln(βG):ln(AcP) with Alox + Feox, the relationship is much closer (, r = 0.90***, n = 30) than with AcP: βG ()). Considering that active Al and Fe bond to soil OM (Dahlgren, Saigusa, and Ugolini Citation2004), repressing the activity of AcP (Kunito et al. Citation2016), the results suggest that active Al and Fe are involved in the decreasing of the potential C:P acquisition activity (r = −0.86***, n = 30) on these soils. Sinsabaugh, Hill, and Shah (Citation2009, Citation2010) also indicated the average of 0.95 for soils (n = 929) as the threshold value below which the energy resource allocation shifts toward Po mineralization on microbial population growth. In the present study, all the observed values are much lower (), either for the weighed mean of the Haplic and Placic Andosols or Vitric Andosols, with values of 0.66 and 0.75, respectively (), as for each individual layer (between 0.74 and 0.91). Therefore, these lower ratios indicate, once more, the low Pi bioavailability of these soils, which leads to an intense AcP production and tends to shift the energy partition from growth toward P acquisition and homeostasis maintenance, especially on the Haplic and Placic Andosols, with higher levels of extractable Alox and Feox. Thus, the limitation on the size of the soil microbial biomass through the energy restriction for microbial growth may also contribute for the OC accumulation in these soils.
Figure 8. Linear relationship of potential of C:P acquisition (ln(βG):ln(AcP)) versus Alox + Feox extractable content, n = 30; the threshold value for energy shift for P acquisition is represented by the dashed line
Cleveland and Liptzin (Citation2007) pointed out an average value of 186 (166 for pasture soils) for the molar C:P ratio, assessed on molar basis of total content of each Sinsabaugh, Hill, and Shah (Citation2009, Citation2010) consider this value as the threshold value above which a P limitation may occur. In our work, it was observed that Vitric Andosols show a value of 84 for the molar C:P ratio, lower than the average value of 145 observed for the Haplic and Placic Andosols (). Vitric Andosols show lower P acquisition effort, when compared to Haplic and Placic Andosols that have higher Al and Fe contents, being related to the observed lower C:P ratio in Vitric Andosols (). Therefore, it seems that the average C:P ratio of 186 as a general threshold value, pointed by the former authors, needs to be adjusted for soils derived from volcanic materials. In these conditions, the abundance of Alox and Feox compounds are responsible for an intense P retention and a lower effective availability of mineral P, their high contents on Pt and Pi.
4. Conclusions
Based on the available results, the following conclusions may be presented:
The management of the soils for the past 20 years (no-till and grazing) led to a strong increment of all the analyzed parameters (OC, Po, Po-Olsen and βG and AcP expression) on the 2 cm top layers, when compared with the 10–15 cm layers;
Activity of soil βG is repressed by both Alox and Feox, being the effect of Alox much closer related to enzyme expression than that of Feox;
The expression of soil AcP seems to be affected by Alox but not by Feox.
The presence of abundant and growing Pi in itself is not a condition for the decrease of soil AcP, since its expression in different soils also depends on their capacity for P retention;
The effort for P acquisition (AcP:βG activity ratio) and the potential of C:P acquisition (ln(βG):ln(AcP)) are both affected by the Pi pools and by Al and Fe contents. Therefore, to maintain the homeostasis condition, the allocation of energy to produce and exudate AcP enzymes decreases, when available Pi exist at levels able to supply microbial growth requirements. On the contrary, when Al and Fe minerals are abundant and Pi retention is high, the energy allocation shifts toward AcP synthesis enzymes, which may affect the microbial growth rate;
The general C:P molar ratio threshold value for soil biomass resource energy partition between growth and P acquisition probably needs to be adjusted for soils derived from volcanic materials.
Acknowledgments
Rui Bajouco was funded by a fellowship granted by the Regional Directorate for Agriculture. The authors would like to thank UTAD Soil and Plants Laboratory staff for technical assistance, and Dr David Walker and Dr Cristina Bajouco for reviewing the English version of this manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Ai, C., G. Liang, J. Sun, X. Wang, and W. Zhou. 2012. “Responses of Extracellular Enzyme Activities and Microbial Community in Both the Rhizosphere and Bulk Soil to Long-term Fertilization Practices in a Fluvo-aquic Soil.” Geoderma 173-174: 330–338. doi:doi:https://doi.org/10.1016/j.geoderma.2011.07.020.
- Alef, K., and P. Nannipieri. 1995. “Enzyme Activities.” In Methods in Applied Soil Microbiology and Biochemistry, edited by K. Alef and P. Nannipieri, 311–373. London: Academic press.
- Allison, S. D. 2006. “Soil Minerals and Humic Acids Alter Enzyme Stability: Implications for Ecosystem Processes.” Biogeochemistry 81: 361–373. doi:doi:https://doi.org/10.1007/s10533-006-9046-2.
- Amano, Y. 1981. “Phosphorus Status of Some Andosols in Japan.” Japan Agricultural Research Quarterly 15: 14–21.
- Bandick, A. K., and R. P. Dick. 1999. “Field Management Effects on Soil Enzyme Activities.” Soil Biology & Biochemistry 31: 1471–1479. doi:doi:https://doi.org/10.1016/S0038-0717(99)00051-6.
- Batjes, N. H. 1996. “Total Carbon and Nitrogen in the Soil of World.” European Journal of Soil Science 65: 4–21. doi:doi:https://doi.org/10.1111/ejss.12114_2.
- Bauer, A., and A. L. Black. 1994. “Quantification of the Effect of Soil Organic Matter Content on Soil Productivity.” Soil Science Society of America Journal 58: 185–193. doi:doi:https://doi.org/10.2136/sssaj1994.03615995005800010027x.
- Blakemore, L. C., P. L. Searle, and B. K. Daly 1987. “Methods for Chemical Analysis of Soils”. NZ Soil Bureau Scientific Reprt 80. Lower Hutt, New Zealand: Department of Scientific and Industrial Research.
- Borie, F., and H. Zunino. 1983. “Organic Matter-phosphorus Associations as a Sink in P-fixation Processes in Allophanic Soils of Chile.” Soil Biology Biochemistry 15: 599–603. doi:doi:https://doi.org/10.1016/0038-0717(83)90056-1.
- Buurman, P., F. Peterse, and M. G. Almendros. 2007. “Soil Organic Matter Chemistry in Allophanic Soils: A pyrolysis-GC/MS Study of A Costa Rican Andosol Catena.” European Journal of Soil Science 58: 1330–1347. doi:doi.https://doi.org/10.1111/j.1365-2389.2007.00925.x.
- Cade-Menun, B. J., M. R. Carter, D. C. James, and C. W. Liu. 2010. “Phosphorus Forms and Chemistry in the Soil Profile under Long-term Conservation Tillage: A Phosphorus-31 Nuclear Magnetic Resonance Study.” Journal of Environmental Quality 39: 1647–1656. doi:doi:https://doi.org/10.2134/jeq2009.0491.
- Caldwell, B. A. 2005. “Enzyme Activities as A Component of Soil Biodiversity: A Review.” Pedobiologia 49: 637–644. doi:doi:https://doi.org/10.1016/j.pedobi.2005.06.003.
- Cannell, R. Q., and J. D. Hawes. 1994. “Trends in Tillage Practices in Relation to Sustainable Crop Production with Special Reference to Temperate Climates.” Soil & Tillage Research 30: 245–282. doi:doi:https://doi.org/10.1016/0167-1987(94)90007-8.
- Cleveland, C. C., and D. Liptzin. 2007. “C:N:P Stoichiometry in Soil: Is There a “Redfield Ratio” for the Microbial Biomass?” Biogeochemistry 85: 235–252. doi:doi:https://doi.org/10.1007/s10533-007-9132-0.
- Coutinho, J. 1996. “The Molybdate/ascorbic Acid Blue Method for the Phosphorus Determination in Very Dilute and Colored Extracts by Segmented Flow Analysis.” Communications in Soil Science and Plant Analisys 27: 1363–1375. doi:doi:https://doi.org/10.1080/00103629609369638.
- Dahlgren, R. A., M. Saigusa, and F. C. Ugolini. 2004. “The Nature, Properties and Management of Volcanic Soils.” Advances in Agronomy 82: 113–182. doi:doi:https://doi.org/10.1016/S0065-2113(03)82003-5.
- Deng, S. P., and M. A. Tabatabai. 1997. “Effect of Tillage and Residue Management on Enzyme Activities in Soils: III. Phosphatases and Arylsulfatase.” Biology and Fertility of Soils 24: 141–146. doi:doi:https://doi.org/10.1007/s003740050222.
- Dick, C. F., A. L. A. Dos-Santos, and J. R. Meyer-Fernandes. 2011. “Inorganic Phosphate as an Important Regulator of Phosphatases.” Enzyme Research 2011: 1–7. doi:doi.https://doi.org/10.4061/2011/103980.
- Dick, R. P. 1997. “Soil Enzyme Activities as Integrative Indicators of Soil Health.” In Biological Indicators of Soil Health, edited by C. E. Pankhurst, B. M. Doube, and V. V. S. R. Cupta, 121–156. Wallingford, England: CABI Publisching.
- Dick, R. P., D. P. Breakwell, and R. F. Turco. 1996. “Soil Enzyme Activities and Biodiversity Measurements as Integrative Microbial Indicators.” In Methods of Assessing Soil Quality, edited by J. W. Doran and A. Jones, 247–271. Madison, Wisconsin, USA: Soil Science Society of America Special Publication.
- Eivazi, F., and M. A. Tabatabai. 1988. “Glucosidases and Galactosidases in Soils.” Soil Biology & Biochemistry 20: 601–606. doi:doi:https://doi.org/10.1016/0038-0717(88)90141-1.
- Eivazi, F., and M. A. Tabatabai. 1990. “Factors Affecting Glucosidase and Galactosidase Activities in Soils.” Soil Biology & Biochemistry 22: 891–897. doi:doi:https://doi.org/10.1016/0038-0717(90)90126-K.
- Eswaran, H., E. V. Berg, and P. Reich. 1993. “Organic Carbon in Soils of the World.” Soil Science Society of America Journal 57: 192–194. doi:doi:https://doi.org/10.2136/sssaj1993.03615995005700010034x.
- Fujita, K., T. Kunito, H. Moro, H. Toda, S. Otsuka, and K. Nagaoka. 2017. “Microbial Resource Allocation for Phosphatase Synthesis Reflects the Availability of Inorganic Phosphorus across Various Soils.” Biogeochemistry 136: 325–339. doi:https://doi.org/10.1007/s10533-017-0398-6.
- Gudmundsson, T., S. T. Gudmundsson, and G. Thorvaldsson. 2014. “Soil Phosphorus Fractionation in Icelandic Long-term Grassland Field Experiments.” Icelandic Agricultural Sciences 27: 81–94.
- Houba, V. J. G., I. Novozamsky, T. M. Lexmond, and J. J. van der Lee. 1990. “As a Single Extraction Solution for the Assessment of the Nutrient Status of Soils and Other Diagnostic Purposes.” Communications in Soil Science and Plant Analisys 21: 2281–2290. doi:doi:https://doi.org/10.1080/00103629009368380.
- Inoue, K., and T. Higashi 1988. “Al– And Fe–humus Complexes in Andisols.” Proceedings 9th International Soil Classification Workshop and Soil Management Support Services. Edited by D. I. Kinloch, S. Shoji, F. M. Beinroth, and H. Eswaran, 535–546. Washington DC.
- Juma, N. G., and M. A. Tabatabai. 1978. “Distribution of Phosphomoesterases in Soils.” Soil Science 126: 101–108.
- Kunito, T., I. Isomura, H. Sumi, H. D. Park, H. Toda, S. Otsuka, K. Nagaoka, K. Saeki, and K. Senoo. 2016. “Aluminum and Acidity Suppress Microbial Activity and Biomass in Acidic Forest Soils.” Soil Biology & Biochemistry 97: 23–30. doi:doi:https://doi.org/10.1016/j.soilbio.2016.02.019.
- Kunito, T., T. Tobitani, H. Moro, and H. Toda. 2012. “Phosphorus Limitation in Microorganisms Leads to High Phosphomonoesterase Activity in Acid Forest Soils.” Pedobiologia 55: 263–270. doi:doi:https://doi.org/10.1016/j.pedobi.2012.05.002.
- Lefebvre, D. D., S. M. G. Duff, C. A. Fife, C. Julien-lnalsingh, and W. C. Plaxton. 1990. “Response to Phosphate Deprivation in Brassica Nigra Suspension Cells.” Plant Physiology 93: 504–511. doi:doi.https://doi.org/10.1104/pp.93.2.504.
- Liang, Q., H. Chen, Y. Gong, H. Yang, M. Fan, and Y. Kuzyakov. 2014. “Effects of 15 Years of Manure and Mineral Fertilizers on Enzyme Activities in Particle-size Fractions in a North China Plain Soil.” European Journal of Soil Biology 60: 112–119. doi:doi:https://doi.org/10.1016/j.ejsobi.2013.11.009.
- Marklein, A. R., and B. Z. Houlton. 2012. “Nitrogen Inputs Accelerate Phosphorus Cycling Rates across a Wide Variety of Terrestrial Ecosystems.” New Phytologist 193: 696–704. doi:doi:https://doi.org/10.1111/j.1469-8137.2011.03967.x.
- Matus, F., C. Rumpel, R. Neculman, M. Panichini, and M. L. Mora. 2014. “Soil Carbon Storage and Stabilisation in Andic Soils: A Review.” Catena 120: 102–110. doi:doi:https://doi.org/10.1016/j.catena.2014.04.008.
- Moro, H., T. Kunito, and T. Sato. 2015. “Assessment of Phosphorus Bioavailability in Cultivated Andisols from a Long-term Fertilization Field Experiment Using Chemical Extractions and Soil Enzyme Activities.” Archives of Agronomy and Soil Science 61: 1107–1123. doi:https://doi.org/10.1080/036503402014984697.
- Nannipieri, P. 1994. “The Potential Use of Soil Enzymes as Indicators of Productivity, Sustainability and Pollution.” In Soil Biota: Management in Sustainable Farming Systems, edited by C. E. Pankhurst, B. Doube, V. Gupta, and P. R. Grace, 238–244. East Melbourne, AU: Commonwealth Scientific and Industrial Research Organization.
- Nannipieri, P., L. Giagnoni, L. Landi, and G. Renella. 2011. “Role of Phosphatase Enzymes in Soil.” In Phosphorus in Action: Biological Processes in Soil Phosphorus Cycling, edited by E. Bunemann, A. Oberson, and E. Frossard, 215–244. Berlin, Germany: Springer-Verlag, Heidelberg.
- Nannipieri, P., E. Kandeler, and P. Ruggiero. 2002. “Enzyme Activities and Microbiological and Biochemical Processes in Soil.” In Enzymes in the Environment: Activity, Ecology, and Applications, edited by R. G. Burns and R. P. Dick, 1–34. Basel, Switzerland: Marcel Dekker AG.
- Olsen, S. R., C. V. Cole, F. Watanabe, and L. Dean 1954. “Estimation of Available Phosphorus in Soil by Extraction with Sodium Bicarbonate”. In Circular (United States. Department of Agriculture) no 939. U.S. Dept. of Agriculture: Washington DC.
- Otani, T., and N. Ae. 1997. “The Status of Inorganic and Organic Phosphorus in Some Soils in Relation to Plant Availability.” Soil Science and Plant Nutrtion 43: 419–429. doi:doi:https://doi.org/10.1080/00380768.1997.10414765.
- Otani, T., and N. Ae. 1999. “Extraction of Organic Phosphorus in Andosols by Various Methods.” Soil Science and Plant Nutrtion 45: 151–161. doi:doi:https://doi.org/10.1080/00380768.1999.10409331.
- Persson, B. L., J. O. Lagerstedt, J. R. Pratt, J. Pattison-Granberg, K. Lundh, S. Shokrollahzadeh, and F. Lundh. 2003. “Regulation of Phosphate Acquisition in Saccharomyces Cerevisiae.” Current Genetics 43: 225–244. doi:doi:https://doi.org/10.1007/s00294-003-0400-9.
- Rao, M. A., and L. Gianfreda. 2000. “Properties of Acid Phosphatase-tannic Acid Complexes Formed in the Presence of Fe and Mn.” Soil Biololgy and Biochemistry 32: 1921–1926. doi:doi:https://doi.org/10.1016/S0038-0717(00)00167-X.
- Rao, M. A., A. Violante, and L. Gianfreda. 2000. “Interaction of Acid Phosphatase with Clays, Organic Molecules and Organo-mineral Complexes: Kinetics and Stability.” Soil Biololgy and Biochemistry 32: 1007–1014. doi:doi:https://doi.org/10.1016/S0038-0717(00)00010-9.
- Redel, Y., R. Rubio, R. Godoy, and F. Borie. 2008. “Phosphorus Fractions and Phosphatase Activity in an Andisol under Different Forest Ecosystems.” Geoderma 145: 216–221. doi:doi:https://doi.org/10.1016/j.geoderma.2008.03.007.
- Reeves, D. W. 1997. “The Role of Soil Organic Matter in Maintaining Soil Quality in Continuous Cropping Systems.” Soil & Tillage Research 43: 131–167. doi:doi:https://doi.org/10.1016/s0167-1987(97)00038-x.
- Rodrigues, M., P. S. Pavinato, P. J. A. Withers, A. P. B. Teles, and W. F. B. Herrera. 2016. “Legacy Phosphorus and No Tillage Agriculture in Tropical Oxisols of the Brazilian Savanna.” Science of the Total Environment 542: 1050–1061. doi:doi:https://doi.org/10.1016/j.scitotenv.2015.08.118.
- Saggar, S., K. R. Tate, C. W. Feltham, C. W. Childs, and A. Parshotam. 1994. “Carbon Turnover in a Range of Allophanic Soils Amended with C-14-Labeled Glucose.” Soil Biololgy and Biochemistry 26: 1263–1271. doi:doi:https://doi.org/10.1016/0038-0717(94)90152-x.
- Saunders, W. M. H., and E. G. Williams. 1955. “Observations on the Determination of Total Organic Phosphorous in Soils.” European Journal of Soil Science 6: 254–267. doi:doi:https://doi.org/10.1111/j.1365-2389.1955.tb00849.x.
- Schmidt, G., and M. Laskowsky Sr. 1961. “Phosphate Ester Cleavage (Survey).” In The Enzymes, edited by P. D. Boyer, H. Lardy, and K. Myrbach, 3–35. 2nd ed. New York: Academic Press.
- Schneider, K., M. B. Turrion, P. F. Grierson, and J. F. Gallardo. 2001. “Phosphatase Activity, Microbial Phosphorus, and Fine Root Growth in Forest Soils in the Sierra De Gata, Western Central Spain.” Biololy and Fertility of Soils 34: 151–155. doi:doi:https://doi.org/10.1007/s003740100387.
- Schwertmann, U. 1964. “Differentiation of Soil Iron Oxides by Extraction with Ammonium Oxalate Solution.” [In German] Zeitschrift Für Pflanzenernährung, Düngung, Bodenkunde 105 (3): 194–202. doi:doi:https://doi.org/10.1002/jpln.3591050303.
- Shoji, S., and J. Masui. 1972. “Amorphous Clay Minerals of Recent Volcanic Ash Soil: III.” Journal of Soil Science and Plant Nutrition 15: 191–201. doi:https://doi.org/10.1080/00380768.1969.10432801.
- Shoji, S., M. Nanzyo, and R. Dahlgren 1993. “Productivity and Utilization of Volcanic Ash Soils.” In Volcanic Ash Soils Genesis, Properties and Utilization. Vol. 21, edited by S. Shoji, M. Nanzyo, and R. Dahlgren, 209–251. Amsterdam: Elsevier Science Publishers b.v. doi:doi:https://doi.org/10.1016/S0166-2481(08)70269-1
- Sinsabaugh, R. L., B. H. Hill, and J. J. F. Shah. 2009. “Ecoenzymatic Stoichiometry of Microbial Organic Nutrient Acquisition in Soil and Sediment.” Nature 462: 795–798. doi:doi:https://doi.org/10.1038/nature08632.
- Sinsabaugh, R. L., B. H. Hill, and J. J. F. Shah. 2010. “Ecoenzymatic Stoichiometry of Microbial Organic Nutrient Acquisition in Soil and Sediment.” Nature 468: 122. doi:doi:https://doi.org/10.1038/nature09548.
- Sinsabaugh, R. L., C. L. Lauber, M. N. Weintraub, B. Ahmed, S. D. Allison, C. Crenshaw, A. R. Contosta, et al. 2008. “Stoichiometry of Soil Enzyme Activity at Global Scale.” Ecology Letters 11:1252–1264. doi:doi:https://doi.org/10.1111/j.1461-0248.2008.01245.x.
- Sinsabaugh, R. L., and D. L. Moorhead. 1994. “Resource Allocation to Extracellular Enzyme Production: A Model for Nitrogen and Phosphorus Control of Litter Decomposition.” Soil Biology & Biochemistry 26: 1305–1311. doi:doi:https://doi.org/10.1016/0038-0717(94)90211-9.
- Sinsabaugh, R. L., and J. J. F. Shah. 2012. “Ecoenzymatic Stoichiometry and Ecological Theory.” Annual Review of Ecology, Evolution, and Systematics 43: 313–343. doi:doi:https://doi.org/10.1146/annurev-ecolsys-110411-160245.
- Šnajdr, J., V. Valášková, V. Merhautová, J. Herinková, T. Cajthaml, and P. Baldrian. 2008. “Spatial Variability of Enzyme Activities and Microbial Biomass in the Upper Layers of Quercus Petraea Forest Soil.” Soil Biology & Biochemistry 40: 2068–2075. doi:doi:https://doi.org/10.1016/j.soilbio.2008.01.015.
- Speir, T. W., and D. J. Ross. 1978. “Soil Phosphatase and Sulphatase.” In Soil Enzymes, edited by R. G. Burns, 197–250. New York: Academic Press.
- Stewart, B. J., and J. M. Leatherwood. 1976. “Derepressed Synthesis of Cellulase by Cellulomonas.” Journal of Bacteriology 128: 609–615.
- Tabatabai, M. A. 1994. “Soil Enzymes.” In Methods of Soil Analysis Part 2: Microbiologial and Biochemical Properties, edited by R. W. Weaver, J. R. Angle, and P. S. Bottomley, 775–833. Wisconsin, USA: Soil Science Society of America: Madison.
- Tate, K. R., and B. K. G. Theng. 1980. “Organic Matter and Its Interactions with Inorganic Soil Constituents.” In Soils with Variable Charge, edited by B. K. G. Theng, 225–249. Lower Hutt, New Zealand: Soil Bureau.
- Tiessen, H., J. W. B. Stewart, and C. V. Cole. 1984. “Pathways of Phosphorus Transformations in Soils of Differing Pedogenesis.” Soil Science Society of America Journal 48: 853–858. doi:doi:https://doi.org/10.2136/sssaj1984.03615995004800040031x.
- Turner, B. L., and P. M. Haygarth. 2005. “Phosphatase Activity in Temperate Pasture Soils: Potential Regulation of Labile Organic Phosphorus Turnover by Phosphodiesterase Activity.” Science of the Total Environment 344: 27–36. doi:doi:https://doi.org/10.1016/j.scitotenv.2005.02.003.
- Turner, B. L., D. W. Hopkins, P. M. Haygarth, and N. Ostle. 2002. ““β-Glucosidase Activity in Pasture Soils.”.” Applied Soil Ecology 20: 157–162. doi:doi:https://doi.org/10.1016/S0929-1393(02)00020-3.
- USEPA.1982. “Methods for Chemical Analysis of Water and Wastes”. EPA 600/4-79-020. US Government Print Office: Washington DC, USA
- Walker, T., and A. Adams. 1958. “Studies on Soil Organic matter:I. Influence of Phosphorus Content of Parent Materials on Accumulations of Carbon, Nitrogen, Sulfur, and Organic Phosphorus in Grassland Soils.” Soil Science 85: 307–318.
- Waring, B. G., S. R. Weintraub, and R. L. Sinsabaugh. 2014. “Ecoenzymatic Stoichiometry of Microbial Nutrient Acquisition in Tropical Soils.” Biogeochemistry 117: 101–113. doi:doi:https://doi.org/10.1007/s10533-013-9849-x.
- WRB, IWG 2006. “World Reference Base for Soil Resources. World Soil Resources Report 103. Rome: Food and Agriculture Organization of the United Nations. ISBN 92-5-10511-4.” Experimental Agriculture 43: 264. doi:doi:https://doi.org/10.1017/S0014479706394902.
- Yan, J., G. Pan, L. Li, G. Quan, C. Ding, and A. Luo. 2010. “Adsorption, Immobilization, and Activity of B-glucosidase on Different Soil Colloids.” Journal of Colloid and Interface Science 348: 565–570. doi:doi:https://doi.org/10.1016/j.jcis.2010.04.044.
- Zheng, M., J. Huang, H. Chen, H. Wang, and J. Mo. 2015. “Responses of Soil Acid Phosphatase and Beta-glucosidase to Nitrogen and Phosphorus Addition in Two Subtropical Forests in Southern China..” European Journal of Soil Biology 68: 77–84. doi:doi:https://doi.org/10.1016/j.ejsobi.2015.03.010.