
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
This study investigated the effect of land-use and management change (LUMC) on carbon (C) and nitrogen (N) dynamics after 15–40 years. LUMC constituted change from rice paddies to chestnut orchard, wetland, and buckwheat upland fields in Shonai region, Yamagata Prefecture, Northeast Japan. Soil samples were collected from the top – (0–15 cm) and sub-layers (15–30 cm) for analysis of soil organic C (SOC) and its δ13C value, total N (TN), and their stocks. C decomposition (Dec-C) and net N mineralization (Net Min-N) were determined according to the production of CO2 and NH4+ + NO3– by aerobic incubation and CO2 + CH4 and NH4+ by anaerobic incubation, respectively. The results reveal that 40 years after change from rice paddy to orchard and wetland, the SOC and TN contents in the top-layer were not significantly altered. However, in buckwheat upland fields and in the parking area where vegetation was absent, the SOC content decreased significantly. Conversion of rice paddies to amur silver grass wetland altered the soil δ13C the most, leading to an increase of 5.1‰ and 2.9‰ for the top – and sub-layers, respectively. In general, the incubation experiment results revealed that a change to orchard and wetland did not significantly decrease the Dec-C and Net Min-N. Whereas, the change from rice paddies to parking area significantly decreased the Dec-C and Net Min-N in both aerobic and anaerobic conditions. We conclude that LUMC over decades had various effects on the SOC and TN contents and stocks as well as their mineralization potentials.
1. Introduction
Rice (Oryza sativa L.) is a staple food in Japan, and rice production is crucial to the nation’s food supply. However, Japanese diets are changing, and the consumption of milk, meat, and bread has overtaken that of rice (Smil and Kobayashi Citation2012). Consequently, the area for rice cultivation has decreased from 3.3 million ha in the 1960s to 1.6 million ha in 2015, leading to numerous types of land-use and management change (LUMC). For example, rice paddies have been turned into orchards and upland fields, built over, or have simply been abandoned (Cheng et al. Citation2018; Shimoda and Wagai Citation2020). LUMC plays a critical role in affecting global carbon (C) and nitrogen (N) cycles (Cheng Citation2020). Crucially, soils constitute the largest C reservoir in terrestrial ecosystems and contain more C than the atmosphere and terrestrial vegetation combined (FAO Citation2017).
C cycling in soils under anaerobic conditions differs in several aspects from that under aerobic conditions. Under anaerobic conditions, O2 supply to the soil is greatly reduced, and alternate electron acceptors such as sulfate (SO42-), nitrate (NO3−), and ferric iron (Fe3+) are utilized by the microbial population during decomposition of organic matter (Mclatchey and Reddy Citation1998). The decomposition of soil organic C (SOC) under anaerobic conditions not only produces CO2 but also CH4, which has a 34 times greater global warming potential than CO2 (IPCC Citation2013). Numerous studies have investigated LUMC and its effects on SOC and total N (TN). Mitsuchi (Citation1974) reported that a LUMC from flooded rice paddy to upland crop cultivation caused substantial losses of soil C, consistent with enhancement of aerobic soil C decomposition. In a more recent case from Japan, the net loss of ecosystem C in the first 15 years was followed by a rapid accumulation that reached an apparent saturation more than 30 years after rice paddies had been abandoned (Shimoda and Wagai Citation2020). Wu et al. (Citation2020) reported that within 12 years the change from paddy to wetland soil led to a 9–25% decrease in dissolved organic C (C stock change was not observed). Nishimura et al. (Citation2008) observed a decrease in SOC content of the surface soil (0–5 cm) from 20.2 to 18.5 g C kg–1 soil within 2.5 years of change from a rice paddy to an upland crop cultivation field. Koga et al. (Citation2020) reported slightly negative CH4 fluxes throughout the year after land-use conversion to upland crop cultivation.
N cycling in submerged rice paddy soils differs from that in upland soils. Because of the anaerobic conditions, the redox potential is maintained at a lower level, and the NH4+ cannot transform into NO3− because it is the dominant form of inorganic N (Cheng et al. Citation2001). A lysimeter study by Takakai et al. (Citation2017), reported that soil N stocks decreased more in upland crop cultivation than those in submerged rice paddy cultivation. In addition, less N leached out during rice paddy cultivation periods, and this was attributable to lower N mineralization and nitrification caused by a limited supply of O2 under flooded conditions.
The focus of these studies was mainly on changes in SOC and TN contents and their labile forms, especially concerning the change from flooded rice paddies to upland farming. The objective of this research was to understand the dynamics of SOC and TN and their mineralization potentials within 15–40 years after the change from submerged rice paddy soils to aerobic agricultural soils. To achieve a comprehensive overview of the effects of LUMC, we also evaluated the impact of the conversion of rice paddies into a parking area and the effects of the establishment of a natural wetland.
2. Material and methods
2.1. Study site
Soil sampling was conducted in a mountain village near the Kumagai Shrine, Shonai town (Shonai region), Yamagata, northeast Japan (38°43ʹN; 140°01ʹE, Figure S1), an area with a typical humid temperate climate. In winter, the site is generally covered by snow for approximately 3 months from the end of December to early March. From 1981 to 2010 (30 years), the mean annual temperature, annual precipitation, and annual cumulative depth of snowfall in the region were 11.7°C, 2,009.9 mm, and 573 cm, respectively (Karikawa Meteorological Station; http://www.data.jma.go.jp/; located 8 km from the sampling site (Figure S2). The area near the sampling site used to be well known for the famous rice variety Kamenoo, which became one of the preeminent varieties in Japan during the Meiji period (1868–1912). The paddy fields are typically cropped once a year, from the end of May to early October, and have been in this use for over 100 years. A part of the area was converted to a chestnut (Castanea crenata) orchard in 1975. A part of this orchard is regularly flooded by spring water and was later abandoned. It evolved into a wetland area in the same year with vegetation characterized overwhelmingly by amur silver grass (Miscanthus sacchariflorus B.). In 1987, one of the rice paddies closed to Kumagai Shrine was converted into a parking area, and the formerly cultivated soil was covered by a layer of small construction stones. In 2011, a 2-cm layer of asphalt was added on top. Since 2000, most rice paddies have been converted into upland fields for cultivating buckwheat (Fagopyrum esculentum). Both rice and buckwheat are cultivated according to conventional methods. In this study, five LUMC types were determined as treatments: 1) rice paddies (continuing rice production for more than a hundred years); 2) conversion to an orchard (chestnut cultivation for 40 years); 3) change to wetland (vegetation dominated by amur silver grass for 40 years); 4) upland field, (buckwheat cultivation for 15 years); 5) parking area, (29 years). The land-use history is summarized in . The map of investigation site and the distance between fields is shown in Figure S1.
Table 1. Cultivation history and management practices of each land-use type and LUMC until 2015 (parking area until 2016)
2.2. Soil sampling and analysis
Soil samples were collected on 23 October 2015 from the top- (0–15 cm) and sub-layers (15–30 cm) on each field using an auger sampler. For the orchard and wetland, nine core soil samples were collected; three cores of each field were mixed, and each mix constituted one replication. The areas of the rice paddies and upland field were approximately two and three times larger, respectively, than those of the other sites, and more samples were collected here. For the rice paddies and upland field, 18 and 27 cores samples were collected, and were mix to six and nine cores, respectively, each mix constituted become one replication. Soil samples from the parking area were collected on 3 December 2016 after digging a soil pit for soil profile description. The 0-cm mark was below the asphalt and stones at the original soil surface. Top- and sub-layer soil samples were collected from the three sides of the profile. All soil samples were air-dried in a greenhouse and sieved to 2 mm to enable measuring SOC and TN. These samples were used for subsequent incubation experiments. In addition, fine soil samples were used to analyze the soil δ13C value via isotope ratio mass spectrometer (IR-MS; Flash 2000, Delta V Plus; Thermo Scientific, Germany; Nguyen-Sy et al. Citation2020). The soil bulk density (ρ) was measured in undisturbed soil at both soil layers, and determined using cylinders of 100 cm−3. The soil C stock (Mg ha−1) in a given horizon was calculated as:
where SOC is the SOC concentration (g kg−1), ρ is the bulk density (g cm−3), and D is the depth (cm). Soil N stocks were calculated using the same formula (Kautsar et al. Citation2020).
For measuring the δ13C value of the main vegetation in the five LUMC fields, leaves of chestnut, amur silver grass and buckwheat were sampled on the same day as soils sampling. The leaves of rice were sampled on 18 September 2016 before harvest. δ13C value was analyzed as soil samples.
2.3. Aerobic incubation
Five grams of the soil samples (oven-dried basis) were placed into 68-mL serum bottles, and distilled water was added with a mini pipette to 50% of the water filled pore space (WFPS). We chose 50% WFPS condition for this aerobic incubation was bases on our preliminary test and referred to Tirol-Padre et al. (Citation2005). To adjust soil moisture to 50% WFPS, the amount of water added to the air-dried soil samples was based on the following equation:
where θv is the volumetric soil moisture, ρ is the soil bulk density, and PD is the soil particle density (2.65 g cm−3 as a fixed constant; Tang et al. Citation2016).
Bottles were capped with a butyl rubber stopper and an aluminum seal. The headspace was flushed with CO2-free air (80% N2 + 20% O2). The samples were then incubated at 25°C. To measure CO2 production, the gas was collected after 2, 4, 6, and 8 weeks in the headspace of each serum bottle. The butyl rubber stopper, aluminum seal, and CO2-free air were replaced after each gas sampling (Nakajima et al. Citation2016; Tang et al. Citation2016). To ensure aerobic conditions during the entire incubation period, the weights of the bottles were measured and the 50% WFPS condition adjusted if applicable. The cumulative CO2 production during 8 weeks was used to calculate C mineralization (Dec-C) under aerobic conditions (no CH4 production). The inorganic N (NH4+-N and NO3–_N) concentrations were measured after 8 weeks of incubation using 10% KCl extractions (JSSSPN Citation1986). To determine Dec-C in the aerobic incubation, we used only CO2, whereas for N mineralization (Net Min-N), we used NH4+-N and NO3–_N after aerobic incubation, minus the initial NH4+-N and NO3–_N content. CO2 and CH4 (for anaerobic incubation only) were analyzed by gas chromatography. NH4+-N and NO3–_N were quantified by colorimetric techniques at 655 and 540 nm, respectively, with a Hitachi U-2900 Spectrophotometer (Hitachi High-Tech Science Corporation, Tokyo, Japan).
2.4. Anaerobic incubation
Five grams of soil samples (oven-dried basis) were placed into 68-mL serum bottles, and 10 mL of distilled water were added (the depth of water from the soil surface is about 1 cm). Bottles were capped with a butyl rubber stopper and an aluminum seal, and the headspace was flushed with N2. The samples were then incubated at 30°C under submerged conditions. After 4 weeks of incubation, soil samples were retrieved from the incubator. The CO2 and CH4 production were measured by collecting gas from the headspace of each bottle. After the gas measurements, soil samples were immediately extracted using 20 mL KCl (final concentration was 10%) to measure the amount of NH4+-N in the soil after anaerobic incubation (Cheng et al. Citation2007). Dec-C for the anaerobic incubation was calculated by adding the amount of CO2 to that of the CH4 emission, whereas Net Min-N was calculated by subtracting the initial amount NH4+-N from the final amount.
2.5. Statistical analysis
Analysis of variance (ANOVA) was used to assess the effect of land use on the measured parameters. If the results from ANOVA showed significant differences (P < 0.05) among land-use types, Tukey’s honest significant difference (HSD) test was used for pairwise comparisons as a post-hoc test. Data evaluation was performed using JMP® Pro 15 Statistics software (SAS Institute Inc., Cary, NC 1989–2021).
3. Results
3.1. Effect of LUMC on SOC, TN, and C/N ratio
At the top layer, the C content significantly decreased from 28.9 g kg−1 to 24.1 g kg−1 and 12.5 g kg−1, respectively, after conversion of rice paddies to upland field and parking area, respectively. By contrast, the C contents in the wetland field and orchard exhibited no significant different from those in the rice paddies. The sub-layer of the upland field showed a significantly decreased C content after the change from rice paddies to upland field (16.6 g kg−1 to 12.8 g kg−1). The TN disclosed a similar pattern to that of SOC in both soil layers (). The TN content of the parking area (1.1 g kg−1) was significantly lower than that of the rice paddies, with a relative difference of approximately 58%. However, the TN contents in the wetland and orchard indicated no significant differences to those of the rice paddies (approximately 2.6 g kg−1 in the top layer). The TN content in the upland field showed a slight decline both in the top- and sub-layers, but the difference from that of the paddy soil was not statistically significant ().
Figure 1. Changes in SOC (a), TN contents (b) and SOC/TN (c) in top- (□0-15 cm) and sub-layers (■15-30 cm) for the five LUMC types. Bars indicate standard deviation. The upper- and lower-case letters indicate significant differences among land-use types in the top- and sub-layers, respectively.
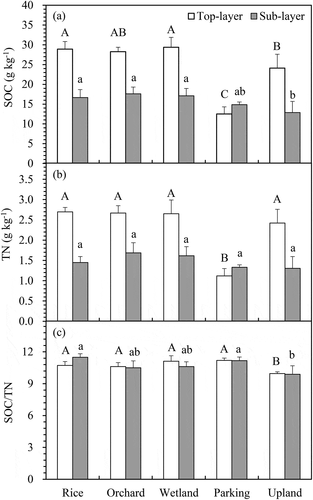
The range of C/N ratio for all land-use types was between 9.9 and 11.5. Excepted upland, there were no differences in soil C/N ratio for both soil layers among rice paddy, orchard, wetland and parking area. However, conversion to upland field led to a significant change in C/N ratio. The C/N ratios of rice paddies was 10.7 and 11.5 in the top and sub-layers, respectively, and decreased to 10.0 and 9.9, respectively, for the upland field ().
3.2. Effect of LUMC on soil δ13C value
The δ13C values of plants for five LUMC types were showed in . δ13C values of rice, chestnut and buckwheat as C3 plants were −28.5, −30.0 and −26.1‰, respectively. The δ13C value of amur silver grass was −12.1‰ ().
Figure 2. The δ13C values of the vegetations (a) and the changes in δ13C values of the soils (b) in the top- (□0-15 cm) and sub-layers (■15-30 cm) among the five LUMC types. Bars indicate standard deviation. The upper- and lower-case letters indicate significant differences among land-use types in the top- and sub-layers, respectively.
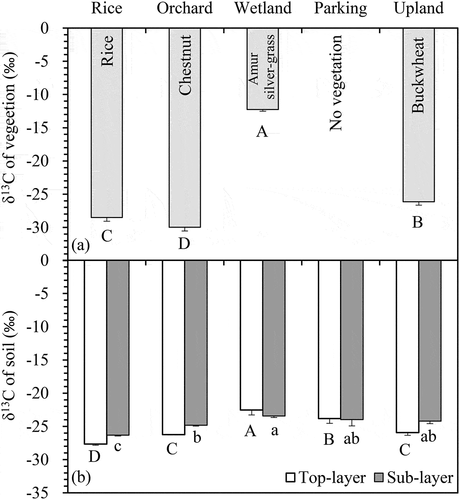
The soil δ13C values differed significantly after land-use change in all treatments (). The δ13C values in the wetland (−22.5‰), orchard (−26.2‰), upland field (−25.9‰), and parking area (−23.8‰) were higher than those in the rice paddies (−27.7‰) in the top-layers. The δ13C values were higher in the sub – than in the top-layer in rice paddies (−26.3‰), orchard (−24.8‰), and upland field (−24.2‰). By contrast, in the wetland (−23.4‰) in the sub-layer values were lower than those in the top layer. The LUMC from rice paddies to wetland field revealed the most significant changes in soil δ13C, with increases of 5.1‰ and 2.9‰ in the top- and sub-layers, respectively, compared to the values of rice paddies ().
3.3. Effect of LUMC on ρ, SOC, and TN stocks
Soil bulk densities (ρ) differed significantly among the five treatments. In the top layer, ρ was the highest in the parking area (1.032 g cm−3) and the lowest in the wetland (0.653 g cm−1). In the sub-layer, bulk density difference among rice paddies, orchard, parking area, and upland field was not significant. However, bulk density in wetland was lower than in the other treatments ().
Figure 3. Changes in soil bulk density (a), SOC (b) and TN stocks (c) in the top- (□0-15 cm) and sub-layers (■15-30 cm) among the five LUMC types. Bars indicate standard deviation. The upper- and lower-case letters indicate significant differences among the five land-uses types in the top- and sub-layers, respectively.
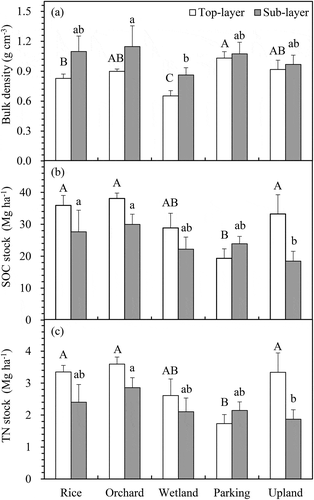
A significant effect of LUMC on SOC stocks was observed only in the parking area in the top layer and in the upland field in the sub-layer. In the top layer (0–15 cm), SOC stocks decreased from 35.9 Mg ha−1 in rice paddies to 19.4 Mg ha−1 in the parking area, while in the sub-layer (15–30 cm), SOC stocks decreased from 27.6 Mg ha−1 to 23.9 Mg ha−1, respectively, for these fields (). Change from rice paddies to parking area led to a significant decreased of TN stocks only in the top layer by 1.6 Mg ha−1 ().
3.4. Effect of LUMC on C and N mineralization under aerobic conditions
In the top-layer, after 8 weeks of aerobic incubation, Dec-C significantly decreased from 837.4 mg kg−1 (rice paddies) to 451.1 mg kg−1 (upland field), 609.8 mg kg−1 (orchard), and 93.9 mg kg−1 (parking area). A significant decrease of Dec-C in the sub-layer was observed only for the change from rice paddies (242.8 mg kg−1) to parking area (123.6 mg kg−1). In the top-layer, the change from rice paddies to all other land-use types led to decrease in the percentage of Dec-C/SOC ratio. In the parking area only 0.75% of SOC in the top layer and 0.83% of SOC in the sub-layer were completely mineralized ().
Table 2. Mineralization indicators of SOC and N at the end of the 8-week aerobic incubation at 25°C for the five LUMC types in the top- and sub-layers
The conversion of rice paddies to upland and parking area significantly reduced the Net Min-N in the top layer from 72.7 mg kg−1 to 30.4 mg kg−1 and 11.1 mg kg−1, respectively. No significant change in Net Min-N was observed in the sub-layer compared to rice paddy. The percentage of Net Min-N/TN ratio decreased largely after the change to upland field and parking area only in the top layer, where 1.26% and 0.99% of TN were mineralized. The average Dec-C/Net Min-N ratios at the end of the aerobic incubation were 11.6 and 17.7 in the top- and sub-layers, respectively ().
3.5. Effect of LUMC on C and N mineralization under anaerobic conditions
After 4 weeks of anaerobic incubation of top-layer soil, Dec-C at 807.5 mg kg−1 (paddy fields) significantly decreased to 187.3 mg kg−1 (parking area) (). The sub-layer exhibited no significant changes of Dec-C. The percentage of Dec-C to SOC ranged from 0.79% to 3.18%, with the lowest ratio observed in the parking area at both top and sub-layers with 1.12% and 0.79% of Dec-C/SOC, respectively (). At the top layer, the highest CH4/Dec-C ratio was observed in rice paddies with 10%. In the sub-layer, the CH4/Dec-C ratios were lower than 1%. There was no CH4 production from parking area in both top- and sub-layers (Table S1).
Table 3. Mineralization indicators of SOC and N at the end of the 4-week anaerobic incubation at 30°C for the five LUMC types in the top- and sub-layers
Except for the parking area, no significant effect of LUMC on N mineralization was observed in both soil layers. The Net Min-N of rice paddies was 299.8 mg kg−1 and 68.1 mg kg−1 in the top- and sub-layers, respectively, and decreased to 21.6 and 21.1 mg kg−1 in the parking area in the top- and sub-layers, respectively. The change to parking area led to decrease in the Net Min-N/TN ratio in the top layer, changing from 11.1% to 1.93%. In the sub-layer, the value decreased from 4.71% (rice paddies), to 2.54% (orchard), 2.39% (wetland), 1.59% (parking area) and 4.07% (upland). The average Dec-C/Net Min-N ratios at the end of the anaerobic incubation were 4.66 and 5.09 in the top- and sub-layers, respectively (), which were lower than those in anaerobic incubation ().
4. Discussion
4.1. Effects of LUMC on SOC, TN, and Soil δ 13C
4.1.1. Change to chestnut orchard
The change to an orchard did not lead to significant changes in SOC and TN contents. In addition, the literature exhibited a gap concerning research related to LUMC from rice paddies to orchards. We assume that the absence of significant changes is probably a result of the chestnut trees biomass including the dry leaves and the grasses around trees in the field at similar levels to those observed in rice paddies. The soil δ13C values were significantly higher in the both soil layers of orchard than the rice paddy, though the δ13C values of chestnut leaves was lower than rice plant (). Two reasons can explain above result. Firstly, the C4 weeds grew in orchard land around chestnut tree should contribute the higher value of δ13C to soil combined with lower value of δ13C from chestnut. Second, the C turnover in aerobic orchard is faster than anaerobic rice paddy (Tete, Viaud, and Walter Citation2015; Chen et al. Citation2021), which led to δ13C enrichment (Nguyen-Sy et al. Citation2020).
4.1.2. Change to wetland
LUMC that involves a change of vegetation from C3 to C4 plants logically enriches soil δ13C because C4 plants are isotopically heavier than C3 plants due to differences in their biosynthetic pathways (West et al. Citation2006). Our results showed that the highest soil δ13C was observed in the wetland for both top- and sub-layers (). This change was caused by the change of vegetation from rice to amur silver grass. Studies have also shown that land-use change from a forest dominated by C3 plants to a pasture dominated by Brachiaria, a C4 plant, increased the soil δ13C value (Tarre et al. Citation2001). Similarly, Durigan et al. (Citation2017) reported an increase of soil δ13C after the conversion of a forest dominated by C3 plants to pasture dominated by C4 plants; the δ13C values were −28.1‰ for forest soil and -24.4‰ for pasture. The change in soil δ13C was not related to a change in the SOC and TN contents in both soil layers. This is possibly due to the similar conditions between rice paddies and wetlands, both of which exhibit soil that is submerged for long periods of time and thus, inhibited decomposition of organic matter (Middleton Citation2020). Wu et al. (Citation2020) reported that conversion from a rice paddy to wetland did not alter SOC stocks but led to a decrease in the labile fraction.
4.1.3. Change to parking area
The C and N contents and stocks observed in the parking area were the lowest of all treatments in both layers. This is logical, because any input of organic matter to the soil was absent. Our results are consistent with those of Zhao et al. (Citation2012) who reported that land-use change by soil sealing significantly decreased soil organic matter (SOM) when compared to the original forest soils. Sealing the soil prevents plant growth and results in the suppression of organic matter input to the soil (Chen et al. Citation2013). Our results indicate that the soil δ13C of the parking area increased although no external input of SOM in the soil surface occurred (). The reason for this increase can be explained by two reasons. Firstly, the easily decomposable organic matter with lower δ13C value from rice cultivation before it was changed to parking area was exhausted. Secondly, the original SOM had higher δ13C value before rice cultivation more than 100 years ago. Thus, further study of soil δ13C alteration under sealed soil is necessary.
4.1.4. Change to upland field
Conversions from rice paddies to other land-use types led to significant changes for SOC, TN, and δ13C in the soil. The change in SOC and TN in the upland was possibly due to the change in the soil’s redox potential. The alteration of soils from rice paddies with poor drainage for long periods to uplands with easy drainage conditions may affect the C and N dynamics (Tete et al. Citation2015). Takakai et al. (Citation2017) reported that the decrease in soil C storage was larger in soybean upland than in paddy rice cultivation, which might be mainly caused by the difference in redox conditions between upland fields (with crop rotation) and continuous paddy fields. The conversion of rice paddies that have anaerobic conditions most of the time to aerobic upland buckwheat plots enhances the degradation of SOM. Anaerobic conditions in paddies weaken the microbial degradation of plant residues, and thus paddy soils tend to accumulate more C in the form of plant-derived C compared with upland soils (Chen et al. Citation2021). In this context, organic matter accumulates more in paddy soils than it does in upland soils, even in the same climate (Kyuma Citation2004). The high accumulation of SOC observed in paddy soils, results from the high input of plant residues combined with retarded decomposition under anaerobic conditions. The δ13C is consistent with the preservation of fresh organic matter input, whereas this material tends to decompose more readily in upland soils due to their aerobic condition. Upland soils are thus characterized by enriched δ13C. This is consistent with the findings of Chen et al. (Citation2021), who state that upland soils are more replenished by microbial-derived C, whereas paddy soils are enriched with plant-derived C. They attributed this effect to the retarded microbial decomposition under anaerobic conditions induced by the flooding of paddies.
4.2. Effects of LUMC on soil C and N dynamics under aerobic and anaerobic incubation
4.2.1. Aerobic incubation
Under aerobic conditions, generally, Dec-C exhibits a similar trend to that of Net Min-N after change to any of the investigated land-use types. Under aerobic conditions, the C mineralization occurring in rice paddies and wetland soil was significantly higher than that of the other treatments. The Dec-C/SOC ratio revealed that 2.89% of SOC was mineralized under aerobic conditions in paddy soil. This is possibly due to the disturbance that the change to an aerobic condition causes in paddy and wetland soil. Thus, the high oxygen availability increased the C mineralization of the plant-derived organic matter that was preserved under anaerobic conditions. Tete, Viaud, and Walter (Citation2015) demonstrated in an incubation experiment that soils at field capacity and in a transient waterlogged condition showed a higher C mineralization than fields that were permanently submerged.
The Dec-C and Net Min-N in orchard, parking area and upland exhibited lower values compared to rice paddy in top-layer. The conversion of rice paddy to chestnut orchard decreased the Dec-C in the top-layer. The Dec-C/SOC ratio showed that 2.16% of SOC was mineralized. This could be related to the stability of carbon fraction in the soil under chestnut trees. Following De Feudis et al. (Citation2020) during the chestnut growth there is a decrease of the labile SOC pool contents and an increase of the stable SOC pool concentrations.
The change to a parking area resulted in a significant decrease in the Dec-C, Net Min-N, percentages of Dec-C/SOC, and Net Min-N/TN ratio under aerobic conditions. A lack of vegetation on the parking area resulted in zero organic matter input from plant residue and root exudates in the soil. This is consistent with the findings of Zhao et al. (Citation2012) who reported that after 10 years of land-use change to impervious land, the net potential N mineralization (2.48 mg kg−1) was significantly decreased compared to that of the original forest land (7.78 mg kg−1). Lack of nutrients and alteration of the environment reduced the microbial community, resulting in the reduction of SOM mineralization.
A change to upland field decreased the Dec-C, Net Min-N, percentage of Dec-C/SOC, and Net Min-N/TN, especially in the top layer. This could be attributable to the different crop biomass inputs. In our previous study, rice paddies had significantly higher aboveground biomass than upland buckwheat fields: The average rice biomass was 910 g m−2 and only 311 g m−2 for upland buckwheat (unpublished data).
4.2.2. Anaerobic incubation
Under anaerobic incubation, only the change to parking area led to a significant decrease in the Dec-C and Net Min-N in both soil layers. Soil sealing followed by no organic matter input to the soil was the main reason for this result. Our results are consistent with those reported by Goyal et al. (Citation1999), who indicated that the decrease of soil organic matter is usually followed by a decrease of microbial biomass and nutrient turn over. Meanwhile, soils from the orchard and wetland areas exhibited a higher Dec-C and Net Min-N in orchard compared to that from rice paddies, although the difference was not statistically significant. These results have to do with the absence of tillage in both the orchard and wetland. Smith et al. (Citation2016) reported that no-till fields tended to have higher nutrient levels and distinct microbial communities related to nutrient cycling compared with fields that were tilled.
Rice paddies exhibited a high CH4/Dec-C ratio, approximately 10.0 in the top layer and 0.4 in the sub-layer. This could be caused by the long continuous rice cultivation under submerged conditions. This finding is consistent with that of Yuan et al. (Citation2016), who investigated the LUMC from flooded rice paddy to upland vegetable fields and observed decreasing CH4 fluxes in two rice growing seasons. In our study, the sub-layer of soil was generally less affected by LUMC than the top layer, except in the parking area.
The Net Min-N/TN ratios under anaerobic incubation after 4 weeks (around 1.59–11.1%) were higher than those under aerobic incubation after 8 weeks (around 0.61–2.70%) (). The large difference between those two incubation conditions was due to the different oxygen availability. Under anaerobic conditions, the mineralized NH4+ could be not nitrified to NO3−. However, mineralized N could be lost by nitrification and sequential denitrification processes under aerobic conditions. Aerobic denitrification is a process reducing the NO3−, into gaseous nitrogen forms in the presence of oxygen gas, which makes the nitrification and denitrification performed simultaneously (Lv et al. Citation2017). As the similar mechanism, Dec-C/Net Min-N ratios (around 3.66–5.09) under anaerobic incubation were lower than those under aerobic incubation (around 11.5–17.7) (), which also suggest that the mineralized N was lost by nitrification and sequential denitrification processes under aerobic conditions.
5. Conclusion
In this study, LUMC affected the dynamics of C and N and their mineralization. The significant decrease of both Dec-C and Net Min-N during the aerobic incubation was observed in upland soil compared to rice paddy soil. LUMC to orchard and wetland did not alter the Dec-C, Net Min-N, or SOC and TN contents. However, the conversion of rice to amur silver grass in the wetland changed the soil δ13C. LUMC to parking area affected C and N dynamics the most by significantly decreasing the SOC and TN contents, Dec-C and Net Min-N under both aerobic and anaerobic incubations. This indicates that soil sealing caused by the construction of the parking area resulted in the decline of SOC and TN stocks. Because the carbon and nitrogen mineralization experiments were conducted in the laboratory, in situ studies are needed to confirm the effect of LUMC on the change of carbon and nitrogen dynamics under real field conditions.
Supplemental Material
Download PDF (495.8 KB)Acknowledgments
First author, Patria Novita Kusumawardani, is thankful to the Japanese Student Services Organization (JASSO) for supporting her studies in Japan from September 2015 to August 2016. We are grateful to the students in the Plant Nutrition and Soil Science Lab of the Faculty of Agriculture, Yamagata University and Mr. Takayuki Chonan of Kumagai Shrine for helping with the soil sampling. This research was funded partly by the Sumitomo Foundation with ID 173275 and the Heiwa Nakajima Foundation.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed here.
References
- Chen, X., Y. Hu, Y. Xia, S. Zheng, C. Ma, Y. Rui, H. He, et al. 2021. “Contrasting Pathways of Carbon Sequestration in Paddy and Upland Soils.” Global Change Biology 27 (11): 2478–2490. doi:https://doi.org/10.1111/gcb.15595.
- Chen, Y., S. D. Day, A. F. Wick, B. D. Strahm, E. Wiseman, and W. L. Daniels. 2013. “Changes in Soil Carbon Pools and Microbial Biomass from Urban Land Development and Subsequent Post-development Soil Rehabilitation.” Soil Biology & Biochemistry 66: 38–44. doi:https://doi.org/10.1016/j.soilbio.2013.06.022.
- Cheng, W., K. Inubushi, K. Yagi, H. Sakai, and K. Kobayashi. 2001. “Effects of Elevated Carbon Dioxide Concentration on Biological Nitrogen Fixation, Nitrogen Mineralization and Carbon Decomposition in Submerged Rice Soil.” Biology and Fertility of Soils 34 (1): 7–13. doi:https://doi.org/10.1007/s003740100345.
- Cheng, W., K. Yagi, H. Akiyama, S. Nishimura, S. Sudo, T. Fumoto, T. Hasegawa, A. E. Hartley, and J. P. Megonigal. 2007. “An Empirical Model of Soil Chemical Properties that Regulate Methane Production in Japanese Rice Paddy Soils.” Journal of Environmental Quality 36 (6): 1920–1925. doi:https://doi.org/10.2134/jeq2007.0201.
- Cheng, W., S. M. Kimani, T. Kanno, S. Tang, A. Z. Oo, K. Tawaraya, S. Sudo, Y. Sasaki, and N. Yoshida. 2018. “Forage Rice Varieties Fukuhibiki and Tachisuzuka Emit Larger CH4 than Edible Rice Haenuki.” Soil Science and Plant Nutrition 64 (1): 77–83. doi:https://doi.org/10.1080/00380768.2017.1378569.
- Cheng, W. 2020. “Soil Carbon and Nitrogen Dynamics by Land Use and Management Changes in East and Southeast Asian Countries (Soil C and N by LUMC).” Soil Science and Plant Nutrition 66 (1): 34–36. doi:https://doi.org/10.1080/00380768.2020.1718923.
- De Feudis, M., G. Falsone, G. Vianello, and L. V. Antisari. 2020. “Stable Organic Carbon Pool Rises in Soil under Chestnut (Castanea Sativa Mill.) Forest for Timber Production after 15 Years since Grafting onto Satin-Cut Stumps.” EQA (International Journal of Environmental Quality) 40 (1): 1–10. doi:https://doi.org/10.6092/.2281-4485/10731.
- Durigan, M. R., M. R. Cherubin, P. B. De Camargo, J. N. Ferreira, E. Berenguer, T. A. Gardner, J. Barlow, et al. 2017. “Soil Organic Matter Responses to Anthropogenic Forest Disturbance and Land Use Change in the Eastern Brazilian Amazon.” Sustainability 9 (3): 379–395. doi:https://doi.org/10.3390/su9030379.
- FAO. 2017. Soil Organic Carbon: The Hidden Potential. Rome, Italy: Food and Agriculture Organization of the United Nations.
- Goyal, S., K. Chander, M. C. Mundra, and K. K. Kapoor. 1999. “Influence of Inorganic Fertilizers and Organic Amendments on Soil Organic Matter and Soil Microbial Properties under Tropical Conditions.” Biology and Fertility of Soils 29 (2): 196–200. doi:https://doi.org/10.1007/s003740050544.
- IPCC. 2013. “Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change.” Cambridge University Press.
- JSSSPN. 1986. Soil Normal Analysis Methods. Tokyo (in Japanese): Japanese Society of Soil Science and Plant Nutrition. Hakuyusha Press.
- Kautsar, V., W. Cheng, K. Tawaraya, S. Yamada, K. Toriyama, and K. Kobayashi. 2020. “Carbon and Nitrogen Stocks and Their Mineralization Potentials are Higher under Organic than Conventional Farming Practices in Japanese Andosols.” Soil Science and Plant Nutrition 66 (1): 144–151. doi:https://doi.org/10.1080/00380768.2019.1705739.
- Koga, N., S. Shimoda, Y. Shirato, T. Kusaba, T. Shima, H. Niimi, T. Yamane, et al. 2020. “Assessing Changes in Soil Carbon Stocks after Land Use Conversion from Forest Land to Agricultural Land in Japan.” Geoderma 377: 114487. doi:https://doi.org/10.1016/j.geoderma.2020.114487.
- Kyuma, K. 2004. Paddy Soil Science. Kyoto: Kyoto Universiy Press.
- Lv, P., J. Luo, X. Zhuang, D. Zhang, Z. Huang, and Z. Bai. 2017. “Diversity of Culturable Aerobic Denitrifying Bacteria in the Sediment, Water and Biofilms in Liangshui River of Beijing, China.” Scientific Reports 7 (1): 10032. doi:https://doi.org/10.1038/s41598-017-09556-9.
- Mclatchey, G. P., and K. R. Reddy. 1998. “Regulation of Organic Matter Decomposition and Nutrient Release in a Wetland Soil.” Journal of Environmental Quality 27 (5): 1268–1274. doi:https://doi.org/10.2134/jeq1998.00472425002700050036x.
- Middleton, B. A. 2020. “Trends of Litter Decomposition and Soil Organic Matter Stocks across Forested Swamp Environments of the Southeastern US.” Plos One 15 (1): 1–23. doi:https://doi.org/10.1371/journal.pone.0226998.
- Mitsuchi, M. 1974. “Characters of Humus Formed under Rice Cultivation.” Soil Science and Plant Nutrition 20 (3): 249–259. doi:https://doi.org/10.1080/00380768.1974.10433247.
- Nakajima, M., W. Cheng, S. Tang, Y. Hori, E. Yaginuma, S. Hattori, S. Hanayama, K. Tawaraya, and X. Xu. 2016. “Modeling Aerobic Decomposition of Rice Straw during the Off-rice Season in an Andisol Paddy Soil in a Cold Temperate Region of Japan: Effects of Soil Temperature and Moisture.” Soil Science and Plant Nutrition 62 (1): 90–98. doi:https://doi.org/10.1080/00380768.2015.1121116.
- Nguyen-Sy, T., W. Cheng, S. M. Kimani, H. Shiono, R. Sugawara, K. Tawaraya, T. Watanabe, and K. Kumagai. 2020. “Stable Carbon Isotope Ratios of Water-extractable Organic Carbon Affected by Application of Rice Straw and Rice Straw Compost during a Long-term Rice Experiment in Yamagata, Japan.” Soil Science and Plant Nutrition 66 (1): 125–132. doi:https://doi.org/10.1080/00380768.2019.1708209.
- Nishimura, S., S. Yonemura, T. Sawamoto, Y. Shirato, H. Akiyama, S. Sudo, and K. Yagi. 2008. “Effect of Land Use Change from Paddy Rice Cultivation to Upland Crop Cultivation on Soil Carbon Budget of a Cropland in Japan.” Agriculture, Ecosystems & Environment 125 (1–4): 9–20. doi:https://doi.org/10.1016/j.agee.2007.11.003.
- Shimoda, S., and R. Wagai. 2020. “Ecosystem Dynamics after Abandonment of Rice Paddy Fields: Does Alien Plant Invasion Enhance Carbon Storage?” Ecosystems 23 (3): 617–629. doi:https://doi.org/10.1007/s10021-019-00426-1.
- Smil, V., and K. Kobayashi. 2012. Japan’s Dietary Transition and Its Impacts. Cambridge: MIT Press.
- Smith, C. R., P. L. Blair, C. Boyd, B. Cody, A. Hazel, A. Hedrick, H. Kathuria, et al. 2016. “Microbial Community Responses to Soil Tillage and CropRotation in a Corn/Soybean Agroecosystem.” Ecology and Evolution 6 (22): 8075–8084. doi:https://doi.org/10.1002/ece3.2553.
- Takakai, F., T. Kikuchi, T. Sato, M. Takeda, K. Sato, S. Nakagawa, K. Kon, T. Sato, and Y. Kaneta. 2017. “Changes in the Nitrogen Budget and Soil Nitrogen in a Field with Paddy–Upland Rotation with Different Histories of Manure Application.” Agriculture 7 (5): 39–52. doi:https://doi.org/10.3390/agriculture7050039.
- Tang, S., W. Cheng, R. Hu, J. Guigue, S. M. Kimani, K. Tawaraya, and X. Xu. 2016. “Simulating the Effects of Soil Temperature and Moisture in the Off-rice Season on Rice Straw Decomposition and Subsequent CH4 Production during the Growth Season in a Paddy Soil.” Biology and Fertility of Soils 52 (5): 739–748. doi:https://doi.org/10.1007/s00374-016-1114-8.
- Tarre, R., R. Macedo, R. B. Cantarutti, C. D. P. Rezende, J. M. Pereira, F. E. Ferreira, B. J. R. Alves, S. Urquiaga, and R. M. Boddey. 2001. “The Effect of the Presence of a Forage Legume on Nitrogen and Carbon Levels in Soils under Brachiaria Pastures in the Atlantic Forest Region of the South of Bahia, Brazil.” Plant and Soil 234 (1): 15–26. doi:https://doi.org/10.1023/A:1010533721740.
- Tete, E., V. Viaud, and C. Walter. 2015. “Organic Carbon and Nitrogen Mineralization in a Poorly-Drained Mineral Soil under Transient Waterlogged Conditions: An Incubation Experiment.” European Journal of Soil Science 66 (3): 427–437. doi:https://doi.org/10.1111/ejss.12234.
- Tirol-Padre, A., K. Tsuchiya, K. Inubushi, and J. K. Ladha. 2005. “Enhancing Soil Quality through Residue Management in a Rice-Wheat System in Fukuoka, Japan.” Soil Science and Plant Nutrition 51 (6): 849–860. doi:https://doi.org/10.1111/j.1747-0765.2005.tb00120.x.
- West, J. B., G. J. Bowen, T. E. Cerling, and J. R. Ehleringer. 2006. “Stable Isotopes as One of Nature’s Ecological Recorders.” Trends in Ecology & Evolution 21 (7): 408–414. doi:https://doi.org/10.1016/j.tree.2006.04.002.
- Wu, X., T. Nguyen-Sy, Z. Sun, T. Watanabe, K. Tawaraya, R. Hu, and W. Cheng. 2020. “Soil Organic Matter Dynamics as Affected by Land Use Change from Rice Paddy to Wetland.” Wetlands 40 (6): 2199–2207. doi:https://doi.org/10.1007/s13157-020-01321-5.
- Yuan, Y., X. Dai, H. Wang, M. Xu, X. Fu, and F. Yang. 2016. “Effects of Land-use Conversion from Double Rice Cropping to Vegetables on Methane and Nitrous Oxide Fluxes in Southern China.” Plos One 11 (5): 1–14. doi:https://doi.org/10.1371/journal.pone.0155926.
- Zhao, D., F. Li, R. Wang, Q. Yang, and H. Ni. 2012. “Effect of Soil Sealing on the Microbial Biomass, N Transformation and Related Enzyme Activities at Various Depths of Soils in Urban Area of Beijing, China.” Journal of Soils and Sediments 12 (4): 519–530. doi:https://doi.org/10.1007/s11368-012-0472-6.