
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
This paper reviews the available vertebrate record from the Lesser Sunda Islands to explore the effect the Last Glacial Maximum had on human subsistence strategies. By focusing on vertebrate assemblages from Laili and Matja Kuru 2 in Timor Leste, Tron Bon Lei in Alor Island, and Here Sorot Entapa in Kisar, this paper investigates biodiversity and resource availability in these nearby islands through the application of standardising indices and statistical testing. Results indicate that vertebrate biodiversity remained fairly stable through and after the Last Glacial Maximum, suggesting that in terms of available mammals, birds and reptiles, this period did not led to severe resource depletion. Hence, potential variations in human subsistence practices or occupation dynamics might not be due to changes in vertebrate diversity. As such, this analysis contributes to investigating anatomically modern humans’ subsistence adaptation in the Lesser Sunda Islands pre- and post-Last Glacial Maximum.
1. Introduction
Archaeological research in Wallacea, the biogeographic region separating the Asian and Australian continental shelves, is igniting exciting research questions. These include the antiquity of human occupations and artistic manifestations (e.g. Brumm et al. Citation2017, Citation2021) migratory routes and the development of navigation techniques (O’Connor and Veth Citation2000; O’Connor Citation2007; Kealy, Louys, and O’Connor Citation2016, Citation2018; O’Connor et al. Citation2017; Norman et al. Citation2018), insights into genetic diversity from the mid-Holocene onwards (McColl et al. Citation2018; Purnomo et al. Citation2021), diversity of cultural practices (Langley, Clarkson, and Ulm Citation2019; Samper-Carro et al. Citation2022), as well as the effect of human interactions with endemic biota (e.g. Louys et al. Citation2021).
Research in Wallacea has traditionally focused on the large islands of Sulawesi, Flores and Timor. Since the beginning of new excavations in Timor-Leste in 2000, a number of sites dating back from the Pleistocene, before and through the Last Glacial Maximu (LGM) have been documented (O’Connor et al., Citation2002; O’Connor and Veth Citation2005; O’Connor Citation2007; O’Connor and Aplin Citation2007; O’Connor et al. Citation2010; O’Connor, Ono, and Clarkson Citation2011; Hawkins et al. Citation2017). To date, the archaeological deposits from Timor-Leste yielded some of the earliest dates for anatomically modern human (AMH) occupations in Wallacea, with Laili (44.6 ka cal BP), Asitau Kuru (former Jerimalai; ca.42 ka cal BP) and Lene Hara (ca.42 ka cal BP) providing the oldest dates (). Sites excavated in Timor-Leste have evidence of episodic occupation, although when the occupations documented in every site are combined, they suggest a continuous occupation of the coastal regions from ca. 44 ka, throughout the LGM, across the Pleistocene-Holocene boundary and throughout the Holocene (O’Connor and Aplin Citation2007).
Figure 1. Map showing the location of the archaeological sites mentioned in the text.
The modern native terrestrial fauna in Timor is poor, limited to a small murid and a tiny shrew, and introduced species such as the New Guinean cuscus (Phalanger orientalis), and Asian domesticates and commensals, translocated in the Late Holocene (Glover Citation1986; O’Connor Citation2015). Fossils of two pygmy stegodons, a giant land tortoise, a large lizard and four to seven genera of giant rat have been reported in Timor (O’Connor and Aplin Citation2007; Aplin and Helgen Citation2010; Louys, Price, and O’Connor Citation2016; Louys et al. Citation2018).
However, not only these larger Wallacean islands have yielded evidence of human occupations. Smaller island are also suitable to sustain human populations, especially when considering how factors such as distance to another landmasses, or the degree of maritime skills, can dramatically affect insular habitability (Keegan et al. Citation2008; Fitzpatrick et al. Citation2016; O’Connor et al. Citation2018). In recent years, a series of archaeological excavations and surveys in the Lesser Sunda islands (i.e. the archipelago from Bali to the West to Wetar to the East; ) are yielding interesting data from these previously poorly explored insular environments (e.g. Louys et al. Citation2017; O’Connor et al. Citation2017). Since 2014, excavations in the islands of Alor, Kisar, Lembata and Pantar are demonstrating how these small insular environments were suitable for human survival from, at least, the late Pleistocene onwards (Samper Carro et al. Citation2016, Citation2018, Citation2019, Citation2021, Citation2022; O’Connor et al. Citation2018; Hawkins et al. Citation2020).
The land-based fauna of the Lesser Sunda Islands is impoverished, consisting only of reptiles (lizards and snakes), several rodent species, shrews, a few species of bats and several birds species (Trainor Citation2005, Citation2007; Trainor et al. Citation2006, Citation2008). However, the diversity of marine ichthyofauna is immense, with thousands of species identified. The introduction of domesticates and agriculture has transformed human subsistence practices in most of these islands.
The Last Glacial Maximum (LGM) – dated from 26,500 to 19,000 years ago based on relative sea level data – entailed the most recent interval in Earth history when ice sheets reached their maximum volume (Mix, Bard, and Schneider Citation2001; Clark et al. Citation2009). In the Northern Hemisphere, especially in northern Europe and America, the limit of the ice sheets during the LGM and their subsequent retreat are well documented, with plentiful records for local mean sea levels (e.g. Yokoyama et al. Citation2000; Lambeck and Chappell Citation2001; Lambeck et al. Citation2014). In the Southern Hemisphere, changes in rainfall and monsoon regimes led to a southward shift of the Intertropical Convergence Zone (ITCZ) (Denniston et al. Citation2013, Citation2017). These shifts affected the Indo-Pacific Warm Pool (IPWP), the ‘global heat engine’, affected its size and other processes, such as deep atmospheric convection and sea surface temperatures (SST) (DiNezio and Tierney Citation2013; De Deckker Citation2016). Additionally, marine sediment samples from offshore Borneo yielded no evidence of a pronounced dry season. For the Lesser Sunda Islands, palynological analysis indicate a reduced overall precipitation, with great interannual variability in some areas (Hope Citation2001, Citation2005; O’Connor and Aplin Citation2007). Sediment samples from cores extracted offshore Sumba, dated to the LGM, indicate enhanced aridity and water stress during the dry season, with dominance of grassland -C4 vegetation- (Dubois et al. Citation2014).
The large number of vertebrate and invertebrate remains recovered in recent excavations in the Lesser Sunda Islands are suitable proxies to investigate the effect that the LGM had on vertebrate diversity. Through the standardisation and statistical testing of the vertebrate assemblages from two coastal sites – Tron Bon Lei (Alor) and Heret Sorot Entapa (Kisar)- and two inland sites – Laili and Matja Kuru 2 (Timor-Leste) – this paper explores the current evidence for vertebrate fauna accumulations dating prior, during and post LGM in these islands. By combining palaeoecological observations based on taxonomic composition with statistical testing of raw and standardised values, new data on human settlement dynamics and fauna diversity in these insular environments from the late Pleistocene to the mid-Holocene are presented.
2. Material and methods
2.1. Archaeological context
2.1.1. Laili (Laleia, Timor-Leste)
Laili is a partially collapsed cave located ca. 86 m asl, about 350 m east to the closest water source, the Laleia River, and 4.3 km from the coast. Three occupational phases are identified in Laili: an earliest period, dated to the Late Pleistocene (44.6–27 ka cal BP); a second period coinciding with the LGM (22.5–18.5 ka cal BP) and a younger period dated to the terminal Pleistocene (17.5–11.2 ka cal BP) (Hawkins et al. Citation2017). A large number of stone artefacts were recovered, with the highest concentration documented in stratigraphic units postdating the LGM (Hawkins et al. Citation2017; ).
Table 1. Total number of remains (NR) for each of the sites analysed in this paper.
Vertebrate remains were recorded throughout the archaeological sequence, being most abundant in the earlier periods of occupation and declining over time (Hawkins et al. Citation2017). The faunal assemblage is dominated by mammals, with sixteen mammal taxa documented, including five small and four extinct giant rodents, bats, birds and translocated domesticates (Hawkins et al. Citation2017). The zooarchaeological material contains evidence of the exploitation of aquatic resources, with abundant molluscs (NISP = 6,686) and few remains of freshwater (i.e. eel) and marine bony fishes (parrotfish), mostly found in the earlier and terminal Pleistocene period, with an abrupt decline during the LGM (Hawkins et al. Citation2017).
2.1.2. Matja Kuru 2 (Poros, Timor-Leste)
Matja Kuru 2 (MK2) is a large cave in an uplifted limestone (ca. 370 m asl), located ca. 8 km inland, north of the Lake Ira Lalaro, the largest freshwater source in Timor. Three occupational phases, based on radiocarbon dates on charcoal and shell, were recorded at MK2: a Late Pleistocene phase (40–31 ka cal BP), an early Holocene phase (11–9.5 ka cal BP) and a late Holocene phase (3.5–2 ka cal BP) (Veth, Spriggs, and O’Connor Citation2005; O’Connor, Robertson, and Aplin Citation2014; Roberts et al. Citation2020).
The faunal assemblage from Matja Kuru 2 is dominated by terrestrial resources, with abundant remains of murids (including giant murids), reptiles, birds and bats, including marine shellfish and fish, as well as a freshwater chelid turtle (O’Connor and Aplin Citation2007; O’Connor, Robertson, and Aplin Citation2014; ). Matja Kuru 2 also produced a complex bone artefact from the Late Pleistocene units, marine shell beads, and a dog burial dated to the Holocene (O’Connor Citation2006; González et al. Citation2013; O’Connor, Robertson, and Aplin Citation2014). The faunal assemblages from these inland sites contrast with those recovered in the Pleistocene coastal sites, which show a focus on aquatic resources.
2.1.3. Tron Bon Lei (Alor, Indonesia)
Tron Bon Lei are two adjacent rock shelters located ca. 33 m asl and 160 m inland in a volcanic ridge on the southwest coast of Alor Island, near Lerabain village. In 2014, three test pits were excavated in Tron Bon Lei, squares A and C in the west-facing shelter and square B in the southerly facing shelter, with two more squares (D and E) opened adjacent to square B in 2018 (Samper Carro et al. Citation2016, Citation2017, Citation2019, Citation2022). The squares in the southern shelter (B, D and E) were excavated down to bedrock at a depth of 3.2 metres, yielding 24 radiocarbon dates from charcoal and marine shell. Modelled dates suggest four occupational phases: a Late Pleistocene phase dated to 21–18.9 ka cal BP, a terminal Pleistocene-early Holocene phase (12.5–9.5 ka cal BP), a middle Holocene phase (8.3–7.2 ka cal BP) and a late Holocene phase (5.4–3 ka cal BP) (Samper Carro et al. Citation2016, Citation2019).
The faunal assemblage from square B in Tron Bon Lei produced a total of 49,984 remains (NR), largely dominated by marine fish, and distributed unevenly throughout the sequence (Samper Carro, Louys, and O’Connor Citation2017; ; S1). Phase I (Late Holocene) yielded few remains (NR = 1,321) compared to Phase II (NR = 19,843), date to the middle Holocene. Phase III, dated to the terminal Pleistocene-early Holocene, produced the largest number of remains overall (NR = 21,994). Phase IV (Late Pleistocene) yielded a total of 6,740 remains.
2.1.4. Here Sorot Entapa (Kisar, Indonesia)
The island of Kisar, 25 km northeast of Timor-Leste, is the smallest island of Wallacea known to have been occupied in the Pleistocene (O’Connor et al. Citation2018). There, Here Sorot Entapa shelter, located about 24 m asl and approximately 80 m from the coast, has evidence of human occupation in two chronological phases: Phase 1, ranging from the terminal Pleistocene to the early Holocene (15.5–9.5 ka cal BP); and Phase 2 dated to the Middle-Late Holocene (4.9–1.8 ka cal BP). The archaeological deposits contained stone artefacts, animal bone and a large quantity of molluscs, urchin and crab, as well as, fishhooks and beads (O’Connor et al. Citation2018).
Fish bones dominate the faunal assemblage, with 26 families identified (; S1). Phase 1 contains a larger number of remains, as well as a higher taxonomic diversity than Phase 2 (O’Connor et al. Citation2018). The rest of the faunal assemblage includes small numbers of reptiles, large and small murid rodents, birds, fruit bats and some marine turtles, with the introduction of Pacific rat, black rat and domestic dogs during the Late Holocene.
2.2. Methods
2.2.1. Analysis of vertebrate remains
The complete archaeological assemblages from one of the excavated 1x1m squares from Laili, Tron Bon Lei and Here Sorot Entapa have been published elsewhere (Hawkins et al. Citation2017; Samper Carro, Louys, and O’Connor Citation2017; O’Connor et al. Citation2018). For the purpose of this study, only basic quantitative units for vertebrate remains (i.e. Number of identifiable specimens; NISP) were considered.
For Matja Kuru 2, the results from the initial zooarchaeological analysis of four of the six 1 × 1 squares excavated are presented here for the first time. Bones were sorted into major categories (Class) and identified to the lowest taxonomic level by reference to the ANU Archaeology and Natural History osteological collection. Vertebrate remains were quantified by recording the total number of remains, including identifiable and unidentifiable identifiable specimens (NR). For Muridae, specimens were classified into three size categories: giant, large and small murid.
To reassure that differences in the relative abundance of taxa among sites are not related to inter-observer errors, the categories and counts published for the assemblages from Laili and Here Sorot Entapa (originally not sorted and classified by the same researcher) were revisited by the author.
2.2.2. Density index calculation
In order to be able to compare the data obtained from the four sites object of study, a standardisation procedure was applied to control whether peaks on faunal remains were due to differences in the excavated volume per spit or otherwise, were the effect of significant variations in the density of material documented. Following methods previously published (e.g. Geiling et al. Citation2018), a density index (DI) was calculated by dividing the number of artefacts (NISP or NR) in each spit by the spit depth in cm (Zspit), which was calculated as the difference between the top (Ztop) and bottom (Zbottom) depth measured in cm, following the formula:
Values for the indexes were later correlated with the chronostratigraphic sequence of the site, in order to provide an overview of occupation dynamics in each of the sites during the different occupational phases. Finally, the indexes were compared between sites to provide a general view of occupation dynamics in the Lesser Sunda Islands during the Pleistocene/early Holocene.
2.2.3. Statistical testing
Statistical testing was conducted to examine (a) samples biodiversity in each site across different chronological periods, (b) distribution of taxa between sites and chronological periods, and (c) effect of climatic events and inter-site variation on the calculated density indexes for fauna. These tests were run separately for tetrapods and ichthys (i.e. cartilaginous and bony fishes), unless otherwise indicated.
Samples biodiversity (including tetrapods and ichthys) was calculated through a dominance index (Simpson index, D) and an information statistic index (Shannon index, H). Unbiased options were selected for both indexes as they are less sensitive to sample size (Hurlbert Citation1971; Chao and Shen Citation2003), and computed as:
Where n is the total number of individuals and S the number of taxa. For each index, 95% centred confidence intervals were computed with a bootstrap procedure. In this procedure, a given number of random samples (9,999) are produced, each with the same total number of individuals as in the original sample. For each individual in the random sample, the taxon is chosen with probabilities proportional to the original abundances (Hammer, Harper, and Ryan Citation2001).
Two nonparametric Kruskal-Wallis H test (i.e. one-way ANOVA on ranks) were conducted to test whether there was a similar distribution of taxa between (a) sites and (b) chronological periods. Correspondence analysis (CA) was chosen as an appropriate algorithm to compare associations between NISP and difference occupational phases (Legendre and Legendre Citation1998). The CA finds the eigenvalues and eigenvectors of a matrix containing the Chi-squared distances between samples. CA also gives the percentages of similarity accounted for by these components (Hammer, Harper, and Ryan Citation2001).
A two–way analysis of variance (ANOVA) was performed to examine the effect of climatic events in different chronological periods (i.e pre-LGM, LGM, and post-LGM) and inter-site variations on density indexes of faunal remains (including tetrapods and ichthys) as an effect of occupation densities. Residual analysis was performed to test for two-way ANOVA’s assumptions. Outliers were assessed by inspection of a boxplot, normality was assessed using Shapiro-Wilk’s normality test and homogeneity of variances was assessed by Levene’s test. Statistical tests were performed on PAST v.4.09 (Hammer, Harper, and Ryan Citation2001) and IBM SPSS Statistics v.28 (IBM SPSS Statistics for Windows Citation2021).
3. Results
3.1. Taxonomic composition
3.1.1. Pre-LGM (45,000-27,000 cal BP)
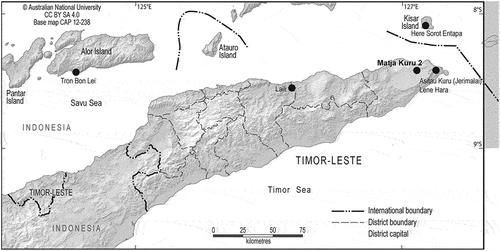
Occupations dating prior to the peak of the Last Glacial Maximum are documented in phase III of Laili (27,000–44,600 cal BP) and phase III of Matja Kuru 2 (31,000–42,000 cal BP). The tetrapod composition in both sites is dominated by, at least, two genera of small Muridae (i.e. Komodomys sp., Melomys sp.) with different size individuals for both genus (Hawkins et al. Citation2017). Giant Muridae, consisting of at least two different genera, were identified in both sites, while the presence of large Muridae in Laili has not been confirmed, although this could be an effect of analysis bias (i.e. skeletal elements misclassified as small or giant murids). The terrestrial or arboreal habitat of the giant murids could not be defined.
The rest of tetrapods identified consist of Chiroptera, including mega- and microbats, Lacertilia (likely skink or gecko species), Serpentes, Anura, and Aves (). The avifauna in Laili included two species of quail, characteristic of open grasslands or wetland, as well as three species of songbirds (Passeriformes), which are likely to be forest or woodland species (Hawkins et al. Citation2017). Specimens from the order Testudines were identified in both sites, although there are a notably larger number of remains in the Matja Kuru 2 (). Some of the skeletal elements identified in Matja Kuru 2 might be classified as Chelodina mccordi timorensis (Kuchling et al. Citation2007; Eisemberg et al. Citation2016), an endemic freshwater species, while no evidence of marine turtles was recorded in Laili and Matja Kuru 2. The ichthyofauna identified in both sites consist of a small number of mostly unidentified bony fish remains (vertebrae, spines, ribs and cranial fragments). Identified taxa include freshwater eel (Anguillidae), sea breams (Sparidae) and wrasses (Labridae).
Figure 2. Distribution of number of tetrapod remains per site in occupational phases dated to before the Last Glacial Maximum. Abbreviations: MK2 = Matja Kuru 2, followed by square (B, C, D or DD).
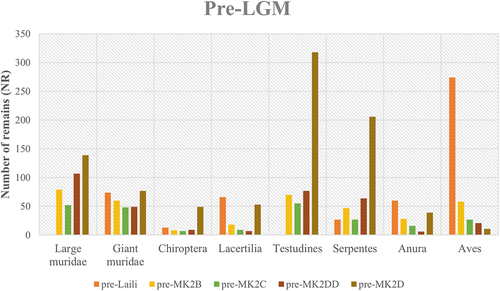
Diversity indices indicate that the taxonomic composition in these occupations is relatively uneven. Laili yielded a dominance score 0.68 (95% CI, 0.65–0.70), with similar scores for squares B and C in Matja Kuru 2, while square DD in this site scored 0.52 (95% CI, 0.49–0.56). Shannon indices range from 0.74 in Laili (95% CI, 0.69–0.78) to 1.10 (95% CI, 1.02–1.17) in square DD from Matja Kuru 2 (; ).
Table 2. Diversity indices calculated for the sites analysed. 95% Confidence interval in brackets.
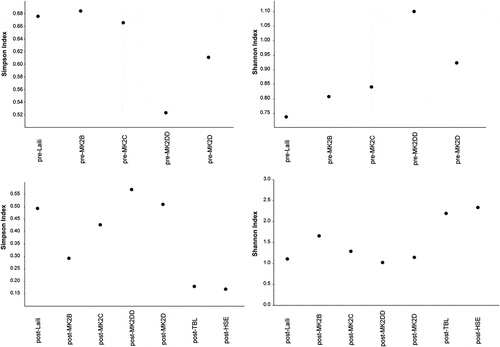
Figure 3. Diversity Indices calculated per site in occupations dated to before (pre-) and after (post-) the Last Glacial Maximum. Abbreviations: MK2 = Matja Kuru 2, followed by square (B, C, D, DD); TBL = Tron Bon Lei; HSE = Here Sorot Entapa.
3.1.2. LGM (22,000-18,500 cal BP)
Occupations coinciding with the Last Glacial Maximum are documented in phase II from Laili (18,500–22,000 cal BP) and phase IV from Tron Bon Lei (18,900–21,000 cal BP). In Laili, taxonomic composition is dominated by small murids (genus Komodomys and Melomys), followed by Aves (songbirds and quails), and two genera of giant murids. In Tron Bon Lei, there is a large amount of snake remains, followed by small murids and small numbers of other taxa (). However, tetrapod fauna represents 2% of the total NR, with abundant remains of cartilaginous and bony fish specimens. Identified fish families include groupers (Serranidae), triggerfish (Balistidae), wrasses (Labridae) and snappers (Lutjanidae) (; S1).
Figure 4. Distribution of number of tetrapod remains per site in occupational phases dated to the Last Glacial Maximum. Abbreviations: TBL = Tron Bon Lei.
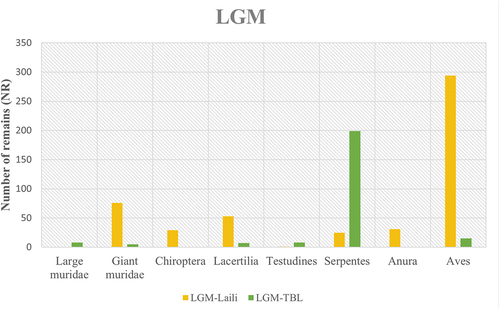
Simpson index produced a score 0.57 (95%CI, 0.55–0.60) for Laili, while the dominance score in Tron Bon Lei is 0.22 (95%CI, 0.20–0.25), indicating a more diverse taxonomic composition (). This result is supported by lower values of the Shannon index for Laili (H = 0.91 95%CI, 0.86–0.97) compared to Tron Bon Lei (H = 1.96 95%CI, 1.86–2.07).
3.1.3. Post-LGM (17,500–9,500 cal BP)
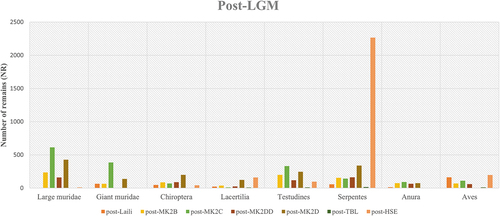
Every site included in this analysis preserved occupational phases dated to after the Last Glacial Maximum, with chronologically overlapping occupations. The tetrapod fauna is dominated by small murids – two genera identified in Laili (Hawkins et al. Citation2017) – with a peak on snake remains in Here Sorot Entapa (). The number of Chiropteras remains in every site increase from previous occupational phases, including the presence of large fruit bats (Pteropodidae) and several genera of microbats (Yangochiroptera). Regarding Testudines, remains of marine turtles (Chelonioidea) are documented in Tron Bon Lei and Here Sorot Entapa, with the presence of freshwater turtles (Chelodina) only in Matja Kuru 2.
Figure 5. Distribution of number of tetrapod remains per site in occupational phases dated to before the Last Glacial Maximum. Abbreviations: MK2 = Matja Kuru 2, followed by the square label (B, C, D or DD); TBL = Tron Bon Lei; HSE = Here Sorot Entapa.
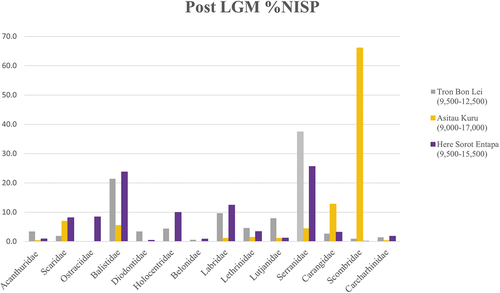
There is a large amount of ichthyofauna documented in Tron Bon Lei and Here Sorot Entapa, representing a total of 26 fish families (; S1). Identified families include a majority of species from reef/near shore habitats (neritic zone), although some families identified in both assemblages, such as Carangidae (jacks and trevallies) and Scombridae (tuna and mackerels), contain open ocean/pelagic species.
Diversity indices show differences between the sites analysed, with inland sites scoring larger D values (0.29 in square B to 0.57 in square DD of Matja Kuru 2) than Tron Bon Lei (D = 0.18, 95%CI, 0.16–0.19) and Here Sorot Entapa (D = 0.17, 95% CI, 0.16–0.18) (Fig. X). Shannon indices indicate diverse taxonomic composition in every site, with Here Sorot Entapa (H = 2.33) and Tron Bon Lei (H = 2.19) yielding the higher scores (; ).
3.2. Statistical testing taxonomic distribution
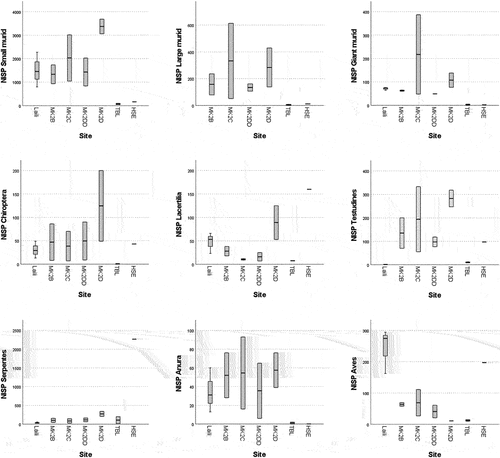
A Kruskal-Wallis H test was run to determine if there were differences in the taxonomic distribution (i.e. species representation) between the different sites analysed. Distribution of species representation scores for tetrapods was similar for all sites, as assessed by visual inspection of the boxplots generated (). Medians and distribution of tetrapods representation scores were not statistically significantly different between sites (). Conversely, medians and distribution of ichthys representation scores were statistically significantly different for several fish families, which representation is restricted to the coastal sites of Tron Bon Lei and Here Sorot Entapa (). Distribution was also dissimilar in these sites.
Table 3. Results of the Kruskal-Wallis test (X2) for the median values per species across the sites analysed.
Figure 6. Boxplots showing the distribution representation scores for tetrapods across sites. For site.
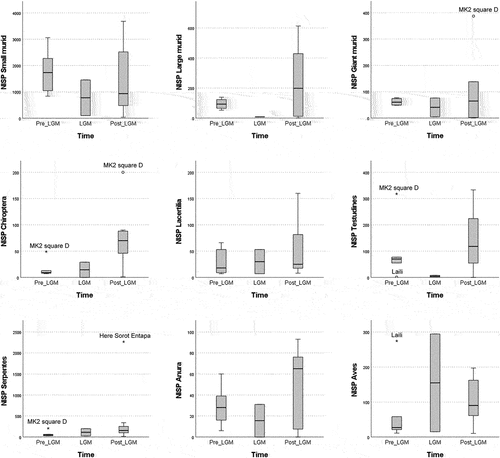
The second Kruskal-Wallis H test, aimed at investigating differences in species representation between the different chronological events analysed, yielded similar results. Taxonomic distribution of scores in tetrapods and ichthys was similar when comparing before, during and after Last Glacial Maximum occupations (). Medians and mean ranks of species distribution scores were not statistically significantly different between chronological periods ().
Table 4. Results of the Kruskal-Wallis test (X2) for the median values per species across the chronological periods analysed.
Figure 7. Boxplot showing the distribution representation scores for tetrapods across chronological periods, indicating the sites that produced extreme outliers (stars) and regular outliers (dots). For site abbreviations, see previous figures.
Axis 1 and 2 of the correspondence analysis (CA) explain 90.55% and 7.43% of the variance in the abundances of taxa, respectively (). The different occupational phases documented in each site group together, indicating inter-site instead of intra-site changes in taxonomic compositions. There is a clear division in axis 1 between inland and coastal sites, with negative values for Tron Bon Lei and Here Sorot Entapa due to the dominance of fish bones (98% of NR in Tron Bon Lei, 96% in Here Sorot Entapa) versus tetrapods in these sites. Laili and Matja Kuru 2 appear clustered together, with similar positive values in axis 1 due to the large number of murids in these sites. Along axis 2, Laili show more positive scores than Matja Kuru 2. Differences in the values between these two sites resides in the differences in numbers between bird remains (6%-10% of total NR in Laili) and turtle remains (2%-3% of total NR in Matja Kuru 2). Overall, the CA demonstrates a clear division between inland and coastal sites based on taxonomic composition, with site-specific variations related to frequency of species but regardless of timing of the occupational phases.
Figure 8. Plot of the correspondence analysis comparing abundance of taxa per site across chronological periods. Yellow dots = pre-LGM; Black dots = LGM; Purple dots = post-LGM. For site abbreviations, see previous figures.
3.3. Statistical testing of density indexes
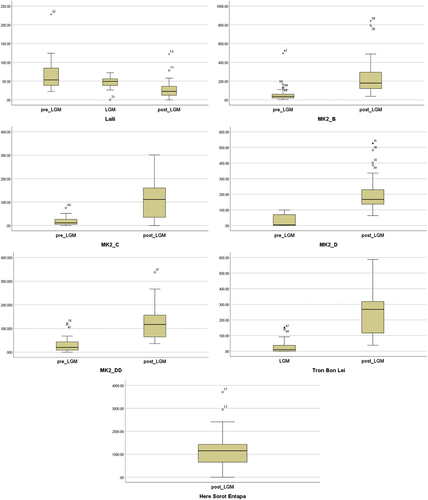
Outliers were identified in most of the sites for the different chronological periods of occupation (). These outliers could be an effect of differences in occupation densities (i.e. larger indexes for more intense occupations and vice versa). Nevertheless, to evaluate whether these outliers had an appreciable effect in the analysis, the same tests were conducted for datasets including and excluding the outliers. The results for the two-way ANOVA conducted excluding outliers did not yield statistically significant results. As such, the analysis continued including outliers. The density indexes for most of the sites considered violated the assumption of normality, as assessed by Shapiro-Wilk’s test (p < .05) ().
Table 5. Results of the Shapiro-Wilk’s normality test (W) for the fauna density indices.
Figure 9. Boxplot showing outliers in the distribution of density indices for each site (numbers correspond to spit numbers). For site abbreviations, see previous figures.
As ANOVAs are considered fairly ‘robust’ to deviations from normality (Maxwell and Delaney Citation2004), the analysis proceeded to evaluating the homogeneity of variances. The assumption of homogeneity of variances was violated, as assessed by the Levene’s test for equality of variances (p < .001). Nevertheless, as data transformation through variance stabilizing transformations (ln, log10 and square root) did not sorted the heterogeneity of variances (scatter plots residuals vs predicted, studentized vs predicted), the analysis continued by the determining the interaction effect between the estimated marginal means and the focal (chronological period) and moderator (site) variables with profile plots () and the test of between-subjects effects.
Figure 10. Profile plots showing the estimated marginal means of the density indices calculated for each site per chronological period.
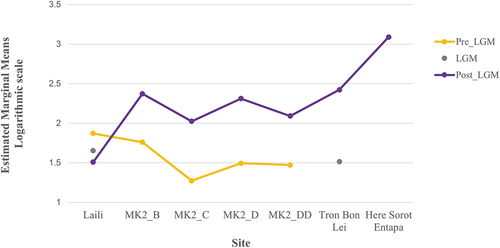
Results indicate that there is no statistically significant interaction effect between the site and chronological period for the density indexes score (F (5,329) = 1.46, p = .204, partial η2 = .022). Therefore, an analysis of the main effect for chronological period and site was performed. There is a statistically significant different in mean density indexes score (i.e. main effect) for chronological period (Fv(2,329) = 7.90, p < .001, partial η2 = .046) and site (F (6,329) = 56.79, p < .001, partial η2 = .509). All pairwise comparisons were run where reported 95% confidence intervals and p-values are Bonferroni adjusted. The unweighted marginal means for pre-LGM, LGM and post-LGM periods were 42.31 (SE = 22.49), 38.96 (SE = 46.45), and 131.46 (SE = 19.01), respectively. Regarding site unweighted marginal means, scores were: Tron Bon Lei 148.9 (SE = 41.18); Laili 50.57 (SE = 38.3); Matja Kuru 2, square B 146.58 (SE = 29.06); Matja Kuru 2, square C 62.2 (SE = 29.61); Matja Kuru 2, square D 118.01 (SE = 37.76); Matja Kuru 2, square DD 76.6 (SE = 29.05); Here Sorot Entapa 1227.4 (SE = 55.08).
No statistically significant differences were observed between pre-LGM occupations, associated with a mean density index score 3.35 (95% CI, −120.84, 127.54) points higher than during the LGM (p > .05). LGM occupation yielded a mean density score 274.94 (95% CI, 395.27, 153.72) points lower than post-LGM occupations, p < .001. Post-LGM occupations are associated with a score 271.15 (95%CI, 200.29, 342) points higher than pre-LGM occupations, statistically significantly different p < .001. Geographically, Here Sorot Entapa is the only site that scores statistically significant differences (p < .001) with every other site tested.
4. Discussion
The analysis of the vertebrate assemblages documented in Laili, Matja Kuru 2, Tron Bon Lei and Here Sorot Entapa provides valuable data to investigate environmental and biodiversity changes related to the Last Glacial Maximum. Additionally, it provides insights into human occupation intensity during the Late Pleistocene-early Holocene in the Lesser Sunda Islands.
Taxonomic composition of tetrapods in the four sites analysed, during every chronological period, follows a similar pattern without statistically significant differences: large number of small murids and varied representation of a diversity of other taxa, including large and giant murids, bats, reptiles, turtles and birds. Laili and Matja Kuru 2 sites produce remains of giant and large murids, with abundant bird remains in Laili and turtle specimens in Matja Kuru 2. As shown by correspondence analyses (), there are no clear differences regarding changes in foraging strategies between diachronic occupational phases. As such, site-specific ‘specialisation’ on distinct taxa could instead be the result of ease of access to specific niches, as lake Ira Lalaro would have been accessible for turtle hunting, and it is likely that arboreal habitats were accessible from Laili. Interestingly, no evidence of LGM occupations has been recorded in MK2; therein lies the question whether this occupational hiatus could be related to changes in water levels in the lake and consequently loss of this resource.
Previous taphonomical analysis on natural (i.e. owl roost) and archaeological assemblages from Tron Bon Lei and Laili documented direct evidence of owl predation in small murids and small birds (Hawkins et al. Citation2018; Hawkins, O’Connor, and Louys Citation2019). However, in these sites, larger animals such as giant murids, freshwater and marine turtles, large fruit bats and medium-large birds are likely to comprise the result of human foraging activities (Hawkins et al. Citation2018; Hawkins, O’Connor, and Louys Citation2019). In Matja Kuru 2, a taphonomic analysis of larger murids, freshwater turtle, birds and fish is ongoing, although preliminary observations suggest an anthropogenic accumulation, with sporadic contributions from avian predators (Samper Carro et al. Citation2022).
A remarkable difference in terms of taxonomic composition is observed between inland and coastal sites. The abundance of fish remains in Tron Bon Lei and Here Sorot Entapa far exceeds the number of any other taxa identified, including small murids. Predictably, a similar pattern is observed when comparing coastal with inland sites within Timor, with fish remains in Lene Hara and Asitau Kuru (formerly Jerimalai) outnumbering terrestrial vertebrates (O’Connor and Aplin Citation2007; O’Connor, Ono, and Clarkson Citation2011).
Similar percentage of NISP values are observed for the different fish families in Here Sorot Entapa and Tron Bon Lei. On the contrary, the percentage of offshore fish, especially Scombridae (tuna and mackerel) in Asitau Kuru, is remarkably higher that in any of the other two sites (). These statistically significant differences (p > 0.05) could be explained in terms of coastal topography, sea levels and structure of fish communities in marine environments. Although the coastal profile in the three islands shows a similar profile, with a nearshore abrupt drop, the distance between the neritic and pelagic zones in each of the coasts varies. The east coast of Timor, in the vicinity of Asitau Kuru, has greater potential for offshore fishing, with a deep channel immediately offshore and opening to the Timor Sea. Hence, preferences in fishing strategies related to coastal and marine current dynamics might have explained the differences observed between these assemblages.
Figure 11. Percentage of number of specimens (NISP) per fish family in Tron Bon Lei, Here Sorot Entapa and Asitau Kuru (former Jerimalai).
In terms of palaeoecological reconstructions, the results from the earliest occupations documented in Timor Leste (Laili and Matja Kuru 2) confirmed previous observations (Hawkins et al. Citation2017). Simpson and Shannon indices indicate a diverse fauna, without single taxa dominating any of the assemblages. There are no significant differences between the different sites and occupational phases, although, as expected, Here Sorot Entapa and Tron Bon Lei yielded higher diversity scores, as a result of the large number of different fish families identified in these assemblages.
Paleoecological inferences from the fauna recovered in Laili support the hypothesis that a mosaic of heterogeneous habitats was present in Timor prior and after the LGM event (Hawkins et al. Citation2017). In relation to the ‘savanna corridor’ hypothesis and the potential absence of forest in the Sunda Islands (Heaney Citation1991; Bird, Taylor, and Hunt Citation2005), the results from Laili and Matja Kuru 2 suggest the presence of forested habitats. Future analysis of the possible habitat of the different genera of giant murids identified in Matja Kuru 2 will increase the available data to assess local forest cover before and after the LGM.
The statistical testing of standardised fauna values based on excavated volumes yielded useful data to assess occupation dynamics. The presence of outliers indicates accumulation peaks at different depth of the deposit. Whether these peaks correspond to the presence of more intense human occupations or otherwise, periods of cave use by avian predators need to be tested in the future. As such, comparing density indexes from vertebrate remains with those obtained from invertebrate remains (i.e. shellfish), stone artefacts and other human by-products (i.e. ochre, shell beads) will provide data to evaluate the degree of anthropogenic contribution to the accumulation of the vertebrate assemblages.
Although no statistically significant differences were observed between before and during LGM occupational phases, differences in the mean density score indicate a larger number of remains in the pre-LGM occupations. Moreover, post-LGM occupations yielded the highest mean density scores among occupational phases. Consequently, occupation dynamics in the Lesser Sunda Islands during these periods show a decrease on intensity during the LGM, bouncing back after this event, as sea levels rapidly rise. Geographically, these changes in density of vertebrate remains accumulation are observed in Here Sorot Entapa, which assemblage yielded an enormous quantity of vertebrate remains compared to the other sites, supporting the hypothesis that small islands in Wallacea were suitable for human inhabitation, at least, from the LGM onwards (O’Connor et al. Citation2018).
The results presented in this paper, even though significant, entail a substantial caveat: the fauna analysis does not include invertebrate remains. Shellfish and other marine invertebrates were a major resource in Wallacean coastal sites during the Pleistocene (e.g. Ono, Soegondho, and Yoneda Citation2009; Ono et al. Citation2020; Szabo and Amesbury Citation2011). In the Lesser Sunda Islands, molluscs and other invertebrates (i.e. barnacles, crabs and sea urchin) usually comprise the larger number of remains documented in coastal sites, such as Here Sorot Entapa and Tron Bon Lei. However, none of these sites provides quantitative data comparable to that obtained from the vertebrate remains, as NISP has not been reported for any of these sites. The invertebrate assemblage from Tron Bon Lei is unpublished, while only a total weight of the invertebrate remains have been reported in Here Sorot Entapa (O’Connor et al. Citation2018). For the Timor sites analysed in this paper, the invertebrate assemblage from Matja Kuru 2 has not been published, with available quantitative data (i.e.NISP) from Laili (Hawkins et al. Citation2017).
Molluscs data from Laili indicate the exploitation of marine, estuarine and freshwater zones during the Pleistocene, with an intensification on harvesting of mangrove molluscs post-LGM (Hawkins et al. Citation2017). During the peak of the LGM, there is a decline in freshwater species, likely reflecting a drop in precipitation during this event, consequently affecting the water flow in the Laleia River and loss or diminishment of freshwater habitats (Hawkins et al. Citation2017). The intensification on the exploitation of marine/brackish environments after the LGM observed in Laili might also be observed in Here Sorot Entapa, where a peak on the presence of marine molluscs is documented in units dated post-LGM (O’Connor et al. Citation2018). Nevertheless, the lack of more specific invertebrate data for these sites hampers correlating these results with those obtained from the vertebrate assemblages in the four sites of study.
5. Conclusions
The analysis of vertebrate remains from sites in Timor, Alor and Kisar provides remarkable data to investigate human occupation dynamics and living conditions in these insular environments before, during and after the Last Glacial Maximum.
Results from the comparison of taxonomic composition in the four sites analysed suggest that this climatic event did not dramatically affect vertebrate diversity, which shows the presence of persistent heterogeneous habitats throughout the Late Pleistocene-early Holocene period. Additionally, fauna diversity suggest the adaptability of human populations in these environments, implying their ability to hunt a myriad of vertebrates, from terrestrial, arboreal and avian species to freshwater and marine taxa. Site-specific changes in species representation can be explained as a result of resource availability and accessibility, instead of defined by climatic constraints.
Although occupation intensity based on fauna density indices shows slight differences among chronological periods and sites, these variations do not seem to indicate periods of extreme resource depletion. Consequently, fauna density indices suggest that the Lesser Sunda Islands were not dramatically affected by the environmental shifts brought by the Last Glacial Maximum and the subsequent rapid sea level rise. Future analyses, aimed at comparing the data obtained from the vertebrate assemblage with that of standardised values from stone artefacts, shellfish and other material culture (i.e. ochre, beads), as well as vertebrate taphonomy, will allow us to investigate the correlation between faunal assemblages and other material providing direct evidence of human involvement in the formation of these archaeological assemblages.
Acknowledgments
Fieldwork and analysis in Laili, Matja Kuru 2, Tron Bon Lei and Here Sorot Entapa was funded by an ARC Discovery Grant (DP0878543), an ARC Laureate Fellowship awarded to Prof. Sue O’Connor (FL120100156), and the ARC Centre of Excellence for Australian Biodiversity and Heritage (CE170100015). Fieldwork in Timor-Leste was undertaken with the assistance of Cecilia Assis and the Ministério do Turismo, Artes e Cultura from Timor-Leste. We thank the representative of the Department of Culture (Mr. Gil), the Department of Forestry (Mr. Marvaos), the Chef de Suco of Mehara (Mr. Antonio) and the Ratu Heads from Poros and Mkury villages and the local villagers for their assistance during fieldwork in Matja Kuru 2. Fieldwork in Indonesia was conducted with the assistance of Dr. Mahirta and staff and students from Universitas Gadja Mada and the Balai Arkeologi Organisation. Fieldwork in Tron Bon Lei was carried out on a FRP visa granted by RISTEK (O’Connor 1304/FRP/SM/V/2014). Fieldwork in Here Sorot Entapa was carried out on a FRP visa granted by RISTEK (O’Connor 1456/FRP/SM/VII/2015).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
Sofia C. Samper Carro
Sofia C. Samper Carro is currently a Research Fellow at the School of Culture, History and Language (CHL) at the Australian National University and a Legacy project manager at the ARC Centre of Excellence for Australian Biodiversity and Heritage (CABAH). Dr. Samper Carro a is zooarchaeologist and taphonomist with a background on Pleistocene and Holocene faunal assemblages. She has a wide experience researching Neanderthal and AMH subsistence strategies, completing taphonomical analysis of human and non-human accumulations, studying human occupation dynamics through density analysis, geometric morphometrics and photogrammetry and 3D modelling. She has also experience on bioanthropological and archaeothanatological analyses of human remains from Island Southeast Asia. Her current projects aim at investigating the possibilities of palaeoproteomic studies to improve our knowledge about Neanderthal and AMH diet and behaviour.
References
- Aplin, K., and K.M. Helgen. 2010. “Quaternary Murid of Timor Part I: New Material of Coryphomis Buehleri Schaub, 1937, and Description of a Second Species of the Genus.” Bulletin of the American Museum of Natural History 341: 1–80.
- Bird, M.I., D. Taylor, and C. Hunt. 2005. “Palaeoenvironments of Insular Southeast Asia during the Last Glacial Period: A Savanna Corridor in Sundaland?” Quaternary Science Reviews 24 (20–21): 2228–2242. doi:10.1016/j.quascirev.2005.04.004.
- Brumm, A., M.C. Langley, M.W. Moore, B. Hakim, M. Ramli, I. Sumantri, B. Burhan, et al. 2017. “Early Human Symbolic Behavior in the Late Pleistocene of Wallacea.” PNAS 114 (16): 4105–4110. doi:10.1073/pnas.1619013114.
- Brumm, A., A.A. Oktaviana, B. Burhan, B. Hakim, R. Lebe, J-X. Zhao, P.H. Sulistyarto, et al. 2021. “Oldest Cave Art Found in Sulawesi.” Science Advances 7 (3): eabd4648. doi:10.1126/sciadv.abd4648.
- Chao, A., and T.J. Shen. 2003. “Nonparametric Estimation of Shannon-s Diversity Index When There are Unseen Species in Sample.” Environmental and Ecological Statistics 10 (4): 429–443. doi:10.1023/A:1026096204727.
- Clark, P.U., A.S. Dyke, J.D. Shakun, A.E. Carlson, J. Clark, B. Wohlfarth, J.X. Mitrovica, et al. 2009. “The Last Glacial Maximum.” Science 325 (5941): 710–714. doi:10.1126/science.1172873.
- De Deckker, P. 2016. “The Indo-Pacific Warm Pool: Critical to World Oceanography and World Climate.” Geoscience Letters 3 (1): 20. doi:10.1186/s40562-016-0054-3.
- Denniston, R.F., Y. Asmerom, V.J. Polyak, A.D. Wanamaker, C.C. Ummenhofer, W.F. Humphreys, J. Cugley, et al. 2017. “Decoupling of Monsoon Activity across the Northern and Southern Indo-Pacific during the Late Glacial.” Quaternary Science Reviews 176 (November): 101–105. doi:10.1016/j.quascirev.2017.09.014.
- Denniston, R.F., K-H. Wyrwoll, Y. Asmerom, V.J. Polyak, W.F. Humphreys, J. Cugley, D. Woods, et al. 2013. “North Atlantic Forcing of Millennial-Scale Indo-Australian Monsoon Dynamics during the Last Glacial Period.” Quaternary Science Reviews 72 (July): 159–168. doi:10.1016/j.quascirev.2013.04.012.
- DiNezio, P.N., and J.E. Tierney. 2013. “The Effect of Sea Level on Glacial Indo-Pacific Climate.” Nature Geoscience 6 (6): 485–491. doi:10.1038/ngeo1823.
- Dubois, N., D.W. Oppo, V.V. Galy, M. Mohtadi, S. van der Kaars, J.E. Tierney, Y. Rosenthal, et al. 2014. “Indonesian Vegetation Response to Changes in Rainfall Seasonality over the past 25,000 Years.” Nature Geoscience 7 (7): 513–517. doi:10.1038/ngeo2182.
- Eisemberg, C.C., B.G. Costa, E.C. Guterres, S.J. Reynolds, and K.A. Christian. 2016. “Notes on Chelodina Mccordi Timorensis Biology, Harvest, Current Threats, and Community Perceptions in the Lake Iralalaro Region, Timor-Leste.” Chelonian Conservation and Biology 15 (1): 69–78. doi:10.2744/CCB-1138.1.
- Fitzpatrick, S.M., V.D. Thompson, A.S. Poteate, M.F. Napolitano, and J.M. Erlandson. 2016. “Marginalization of the Margins: The Importance of Smaller Islands in Human Prehistory.” Journal of Island and Coastal Archaeology 11 (2): 155–170. doi:10.1080/15564894.2016.1192568.
- Geiling, J.M., A.B. Marín-Arroyo, L.G. Straus, and M.R. González Morales. 2018. “Deciphering Archaeological Palimpsests with Bone Micro-Fragments from the Lower Magdalenian of El Mirón Cave (Cantabria, Spain).” Historical Biology 30 (6): 730–742. doi:10.1080/08912963.2017.1385611.
- Glover, I. 1986. Archaeology in East Timor, 1966–67. Vol. 11. Terra Australis. Canberra: Department of Prehistory, Research School of Pacific Studies, Australian National University.
- González, A., G. Clark, S. O’Connor, and L. Matisoo-Smith. 2013. “A 3000 Years Old Dog Burial in Timor-Leste.” Australian Archaeology 76: 13–20. doi:10.1080/03122417.2013.11681961.
- Hammer, O, D.A.T. Harper, and P.D. Ryan. 2001. “PAST: Paleontological Statistics Software Package for Education Adn Data Analysis.” Palaeontologia Electronica 4 (1): 1–9.
- Hawkins, S., F.S. Arumdhati, M. Litster, T.S. Lim, G. Basile, M. Leclerc, C. Reepmeyer, et al. 2020. “Metal-Age Maritime Culture at Jareng Bori Rockshelter, Pantar Island, Eastern Indonesia.” Records of the Australian Museum 72: 237–262. doi: 10.3853/j.2201-4349.72.2020.1726.
- Hawkins, S., S. O’Connor, and J. Louys. 2019. “Taphonomy of Bird (Aves) Remains at Laili Cave, Timor-Leste, and Implications for Human-Bird Interactions during the Pleistocene.” Archaeological and Anthropological Sciences 11 (12): 6325–6337. doi:10.1007/s12520-017-0568-4.
- Hawkins, S., S. O’Connor, T.R. Maloney, M. Litster, S. Kealy, J.N. Fenner, K. Aplin, et al. 2017. “Oldest Human Occupation of Wallacea at Laili Cave, Timor-Leste, Shows Broad-Spectrum Foraging Responses to Late Pleistocene Environments.” Quaternary Science Reviews 171: 58–72. doi: 10.1016/j.quascirev.2017.07.008.
- Hawkins, S., S.C. Samper Carro, J. Louys, K. Aplin, S. O’Connor, and Mahirta. 2018. “Human Palaeoecological Interactions and Owl Roosting at Tron Bon Lei, Alor Island, Eastern Indonesia.” The Journal of Island and Coastal Archaeology 13 (3): 371–387. doi:10.1080/15564894.2017.1285834.
- Heaney, L.R. 1991. “A Synopsis of Climatic and Vegetational Change in Southeast Asia.” Climatic Change 19 (1–2): 53–61. doi:10.1007/BF00142213.
- Hope, G. 2001. “Environmental Change in the Late Pleistocene and Late Holocene at Wanda Site, Soroako, South Sulawesi, Indonesia.” Palaeogeography, Palaeoclimatology, Palaeoecology 171: 129–145. doi:10.1016/S0031-0182(01)00243-7.
- Hope, G. 2005. “The Quaternary in Southeast Asia.” In The Physical Geography of Southeast Asia, edited by A. Gupta, 24–37. Oxford: Oxford University Press.
- Hurlbert, S.H. 1971. “The Nonconcept of Species Diversity: A Critique and Alternative Parameters.” Ecology 52: 577–586. doi:10.2307/1934145.
- IBM SPSS Statistics for Windows. 2021. Windows. Armonk, NY: IBM Corp.
- Kealy, S., J. Louys, and S. O’Connor. 2016. “Islands under the Sea: A Review of Early Modern Human Dispersal Routes and Migration Hypotheses through Wallacea.” The Journal of Island and Coastal Archaeology 11 (3): 364–384. doi:10.1080/15564894.2015.1119218.
- Kealy, S., J. Louys, and S. O’Connor. 2018. “Least-Cost Pathway Models Indicate Northern Human Dispersal from Sunda to Sahul.” Journal of Human Evolution 125: 59–70. doi:10.1016/j.jhevol.2018.10.003.
- Keegan, W.F., S.M. Fitzpatrick, K. Sullivan Sealey, M.J. LeFebvre, and P.T. Sinelli. 2008. “The Role of Small Islands in Maritime Subsistence Strategies: Case Studies from the Caribbean.” Human Ecology 36: 635–654. doi:10.1007/s10745-008-9188-z.
- Kuchling, G., A.G.J. Rhodin, B.R. Ibarrondo, and C.R. Trainor. 2007. “A New Subspecies of the Snakeneck Turtle Chelodina Mccordi from Timor-Leste (East Timor) (Testudines: Chelidae).” Chelonian Conservation and Biology 6 (2): 213. doi:10.2744/1071-8443(2007)6[213:ANSOTS]2.0.CO;2.
- Lambeck, K., and J. Chappell. 2001. “Sea Level Change through the Last Glacial Cycle.” Science 292: 679–686. doi:10.1126/science.1059549.
- Lambeck, K., H. Rouby, A. Purcell, Y. Sun, and M. Sambridge. 2014. “Sea Level and Global Ice Volumes from the Last Glacial Maximum to the Holocene.” Proceedings of the National Academy of Sciences 111 (43): 15296–15303. doi:10.1073/pnas.1411762111.
- Langley, M.C., C. Clarkson, and S. Ulm. 2019. “Symbolic Expression in Pleistocene Sahul, Sunda, and Wallacea.” Quaternary Science Reviews 221: 105883. doi:10.1016/j.quascirev.2019.105883.
- Legendre, P., and L. Legendre. 1998. Numerical Ecology. Elsevier Science.
- Louys, J., T.J. Braje, C.-H. Chang, R. Cosgrove, S.M. Fitzpatrick, M. Fujita, S. Hawkins, et al. 2021. “No Evidence for Widespread Island Extinctions after Pleistocene Hominin Arrival.” Proceedings of the National Academy of Sciences 118 (20): e2023005118. doi:10.1073/pnas.2023005118.
- Louys, J., S. Kealy, S. O’Connor, G.J. Price, S. Hawkins, K. Aplin, Y. Rizal, et al. 2017. “Differential Preservation of Vertebrates in Southeast Asian Caves.” International Journal of Speleology 46 (3): 379–408. doi:10.5038/1827-806X.46.3.2131.
- Louys, J., S. O’Connor, Mahirta, P. Higgins, S. Hawkins, and T.R. Maloney. 2018. “New Genus and Species of Giant Rat from Alor Island, Indonesia.” Journal of Asia-Pacific Biodiversity 11 (4): 503–510. doi:10.1016/j.japb.2018.08.005.
- Louys, J., G.J. Price, and S. O’Connor. 2016. “Direct Dating of Pleistocene Stegodon from Timor Island, East Nusa Tenggara.” PeerJ 4: e1788. doi:10.7717/peerj.1788.
- Maxwell, S. E., and H.D. Delaney. 2004. ”Designing Experiments and Analyzing Data: A Model Comparison Perspective. 2nd ed.)” ed. Mahwah: Lawrence Erlbaum Associates.
- McColl, H., F. Racimo, L. Vinner, F. Demeter, U.G. Wilken, J.V. Moreno Mayar, A. Seguin-Orlando, et al. 2018. “The prehistoric peopling of Southeast Asia.” Science 361 (6397), 88–92.
- Mix, A.C., E. Bard, and R. Schneider. 2001. “Environmental Processes of the Ice Age: Land, Oceans, Glaciers (EPILOG).” Quaternary Science Reviews 20 (4): 627–657. doi:10.1016/S0277-3791(00)00145-1.
- Norman, K., J. Inglis, C. Clarkson, J. Tyler Faith, J. Shulmeister, and D. Harris. 2018. “An Early Colonisation Pathway into Northwest Australia 70–60,000 Years Ago.” Quaternary Science Reviews 180: 229–239. doi:10.1016/j.quascirev.2017.11.023.
- O’Connor, S. 2006. “Continuity in Shell Artefact Production in Holocene in East Timor.” In Uncovering Southeast Asia’s Past. edited by E.A. Bacus, I. Glover, V.C. Pigott, and P.D. Sharrock, 218–233. Selected papers from the 10th International Conference of the European Association of Southeast Asian Archaeologists, NUS Press.
- O’Connor, S. 2007. “New Evidence from East Timor Contributes to Our Understanding of Earliest Modern Human Colonization East of Sunda Shelf.” Antiquity 81: 523–535. doi:10.1017/S0003598X00095569.
- O’Connor, S. 2015. “Rethinking the Neolithic in Island Southeast Asia, with Particular Reference to the Archaeology of Timor-Leste and Sulawesi.” Archipel 90: 15–47. doi:10.4000/archipel.362.
- O’Connor, S., and K. Aplin. 2007. “A Matter of Balance: An Overview of Pleistocene Occupation History and the Impact of the Last Glacial Phase in East Timor and the Aru Islands, Eastern Indonesia.” Archaeology in Oceania 42: 82–90. doi:10.1002/j.1834-4453.2007.tb00021.x.
- O’Connor, S., A. Barham, M. Spriggs, P. Veth, K. Aplin, and E. St Pierre. 2010. “Cave Archaeology and Sampling Issues in the Tropics: A Case Study from Lene Hara Cave, A 42,000 Year Old Occupation Site in East Timor, Island Southeast Asia.” Australian Archaeology 71: 29–40. doi:10.1080/03122417.2010.11689382.
- O’Connor, S., J. Louys, S. Kealy, and S.C. Samper Carro. 2017. “Hominin Dispersal and Settlement East of Huxley’s Line. The Role of Sea Level Changes, Island Size, and Subsistence Behavior.” Current Anthropology 58 (17): S567–82. doi:10.1086/694252.
- O’Connor, S., Mahirta, S. Kealy, C. Boulanger, T. Maloney, S. Hawkins, M.C. Langley, et al. 2018. “Kisar and the Archaeology of Small Islands in the Wallacean Archipelago.” The Journal of Island and Coastal Archaeology 1–28. doi:10.1080/15564894.2018.1443171.
- O’Connor, S., R. Ono, and C. Clarkson. 2011. “Pelagic Fishing at 42,000 Years before the Present and the Maritime Skills of Modern Humans.” Science 334: 1117–1121. doi:10.1126/science.1207703.
- O’Connor, S., G. Robertson, and K.P. Aplin. 2014. “Are Osseous Artefacts a Window to Perishable Material Culture? Implications of an Unusually Complex Bone Tool from the Late Pleistocene of East Timor.” Journal of Human Evolution 67: 108–119. doi:10.1016/j.jhevol.2013.12.002.
- O’Connor, S., M. Spriggs, and P. Veth. 2002. “Excavation at Lene Hara Cave Establishes Occupation in East Timor at Least 30,000–35,000 Years Ago.” Antiquity 76: 45–49. doi:10.1017/S0003598X0008978X.
- O’Connor, S., and P. Veth. 2000. “The World’s First Mariners: Savannah Dwellers in an Island Continent.” In East of Wallace’s Line. Modern Quaternary Research in Southeast Asia, Vol. 16, edited by S. O’Connor and P. Veth, 99–137. Rotterdam, Netherlands: Balkema Publishers. Modern Quaternary Research in Southeast Asia 16. A.A.
- O’Connor, S., and P. Veth. 2005. “Early Holocene Fish Hooks from Lene Hara Cave, East Timor Establish Complex Fishing Technology Was in Use in Island South East Asia Five Thousand Years before Austronesian Settlement.” Antiquity 79: 249–256. doi:10.1017/S0003598X0011405X.
- Ono, R., R. Fuentes, A. Pawlik, H.O. Sofian, Sriwigati, N. Aziz, N. Alamsyah, and M. Yoneda. 2020. “Island Migration and Foraging Behaviour by Anatomically Modern Humans during the Late Pleistocene to Holocene in Wallacea: New Evidence from Central Sulawesi, Indonesia.” Quaternary International 554: 90–106. doi:10.1016/j.quaint.2020.03.054.
- Ono, R., S. Soegondho, and M. Yoneda. 2009. “Changing Marine Exploitation during the Late Pleistocene in Northern Wallacea: Shell Remains from Leang Sarru Rockshelter in Talaud Islands.” Asian Perspectives 2: 318–341.
- Purnomo, G.A., K.J. Mitchell, S. O’Connor, S. Kealy, L. Taufik, S. Schiller, A. Rohrlach, et al. 2021. “Mitogenomes Reveal Two Major Influxes of Papuan Ancestry across Wallacea following the Last Glacial Maximum and Austronesian Contact.” Genes 12 (7): 965. doi:10.3390/genes12070965.
- Roberts, P., J. Louys, J. Zech, C. Shipton, S. Kealy, S.C. Samper Carro, S. Hawkins, et al. 2020. “Isotopic Evidence for Initial Coastal Colonization and Subsequent Diversification in the Human Occupation of Wallacea.” Nature communications 11 (1): 2068.
- Samper Carro, S.C., F. Gilbert, D. Bulbeck, S. O’Connor, J. Louys, N. Spooner, D. Questiaux, et al. 2019. “Somewhere beyond the Sea: Human Cranial Remains from the Lesser Sunda Islands (Alor Island, Indonesia) Provide Insights on Late Pleistocene Peopling of Island Southeast Asia.” Journal of Human Evolution 134:102638. doi:10.1016/j.jhevol.2019.07.002.
- Samper Carro, S.C., J. Louys, and S. O’Connor. 2017. “Methodological Consideration for Ichtyoarchaeology from the Tron Bon Lei Sequence, Alor, Indonesia.” Archaeological Research in Asia 12: 11–22. doi:10.1016/j.ara.2017.09.006.
- Samper Carro, S.C., J. Louys, and S. O’Connor. 2018. “Shape Does Matter: A Geometric Morphometric Approach to Shape Variation in Indo-Pacific Fish Vertebrae for Habitat Identification.” Journal of Archaeological Science 99: 124–134. doi:10.1016/j.jas.2018.09.010.
- Samper Carro, S.C., S. O’Connor, J. Louys, S. Hawkins, and M. Mahirta. 2016. “Human Maritime Subsistence Strategies in the Lesser Sunda Islands during the Terminal Pleistocene-Early Holocene: New Evidence from Alor, Indonesia.” Quaternary International 416: 64–79. doi:10.1016/j.quaint.2015.07.068.
- Samper-Carro, S.C., S. O’Connor, Mahirta, S. Kealy, and C. Shipton. 2022. “Talking Dead. New Burials from Tron Bon Lei (Alor Island, Indonesia) Inform on the Evolution of Mortuary Practices from the Terminal Pleistocene to the Holocene in Southeast Asia.” PLOS ONE 17 (8): e0267635. doi:10.1371/journal.pone.0267635.
- Samper Carro, S.C., C. Raymond, V. Weisbecker, and S. O’Connor. 2022. “From Little Things, Big Things Grow: Investigating Human Involvement in the Accumulation of Small Vertebrates in Matja Kuru 2 (Timor-Leste) during the Late Pleistocene.” Journal of Taphonomy 16 (1–4): 177–178.
- Samper Carro, S.C., T.J. Stewart, Mahirta, R. Wood, and S. O’Connor. 2021. “Burial Practices in the Early Mid-Holocene of the Wallacean Islands: A Sub-Adult Burial from Gua Makpan, Alor Island, Indonesia.” Quaternary International 603 (November): 125–138. doi:10.1016/j.quaint.2020.10.004.
- Szabo, K., and J.R. Amesbury. 2011. “Molluscs in a World of Islands: The Use of Shellfish as a Food Resource in the Tropical Island Asia-Pacific Region.” Quaternary International 239: 8–18. doi:10.1016/j.quaint.2011.02.033.
- Trainor, C.R. 2005. “Species Richness, Habitat Use and Conservation of Birds of Alor Island, Lesser Sundas, Indonesia.” Emu 105: 127–135. doi:10.1071/MU03030.
- Trainor, C.R. 2007. “Changes in Bird Species Composition on a Remote and Well-Forested Wallacean Island, South-East Asia.” Biological Conservation 140: 373–385. doi:10.1016/j.biocon.2007.08.022.
- Trainor, C.R., P.J. Benstead, K. Martin, D. Lesmana, D. Agista, M.C. Benstead, R. Drijvers, and I. Setiawan. 2006. “New Bird Records for Nusa Tenggara Islands: Sumbawa, Moyo, Sumba, Flores, Pulau Besar and Timor.” Kukila 13: 6–22.
- Trainor, C.R., F. Santana, P. Pinto, F.X. Almeida, R. Safford, and R. Grimmet. 2008. “Birds, Birding and Conservation in Timor-Leste.” Birding Asia 9: 16–45.
- Veth, P., M. Spriggs, and S. O’Connor. 2005. “Continuity of Cave Use in the Tropics: Examples from East Timor and the Aru Islands, Maluku.” Asian Perspectives 44 (1): 180–192. doi:10.1353/asi.2005.0015.
- Yokoyama, Y., K. Lambeck, P. De Deckker, P. Johnston, and L.K. Fifield. 2000. “Timing of the Last Glacial Maximum from Observed Sea-Level Minima.” Nature 406 (6797): 713–716.