Abstract
Quantifying diurnal patterns of water use efficiency (WUE) and radiation use efficiency (RUE) for wheat and maize is important for assessing water use by plants and crop productivity. Water and carbon dioxide fluxes from an irrigated wheat-maize double-crop field from November 2002 to October 2003 were measured using the Eddy Covariance method. Evident differences were observed between the diurnal patterns of WUE for wheat and maize. The WUE values of wheat peaked near 9, 15 and 12 mg CO2 g H2O in the morning, and then decreased linearly with time and recovered in the late afternoon (4:00pm) before sunset in March, April and May, respectively. The WUE of maize increased after sunrise and retained stable values of 6, 14 and 12 mg CO2 g H2O from mid-morning to mid-afternoon (10:00am –2:00pm) and then decreased slowly with time until sunset in July, August and September, respectively. Similar patterns were observed in the RUE of wheat and maize. Over the three months of the study, averaged RUE was 1.76 g C MJ−1 for the wheat crop and 1.87 g C MJ−1 for the maize crop. A coupled photosynthesis and transpiration model was used to simulate the diurnal variations in WUE under variable climate conditions. Measurement results and sensitivity analysis show that the difference in the diurnal variation pattern in WUE between wheat and maize resulted from the different carbon fixing mechanisms of wheat and maize.
Introduction
The North China Plain (NCP), which has a dominant wheat-maize double-crop system, is the largest agricultural production area in China. It provides more than 50% of the nation's wheat and about 33% of its maize (State Statistics Bureau Citation1999). However, precipitation decreases from 910 mm y−1 in the south to 470 mm y−1 in the north and about 70% is concentrated in the summer (Wu et al. Citation2006). Winter wheat needs irrigating especially in the northern part of the NCP. Agriculture irrigation uses about 70–80% of the total water resources in the NCP (Wang et al. Citation2008). Increased irrigation has led to a rapid decline of the groundwater table. Water shortage has become a major limiting factor of wheat-maize production in the NCP (Zhang et al. Citation2005). Therefore, there is an urgent need to improve resource use efficiency to ensure the sustainability of agriculture production in the NCP.
WUE and RUE are two important parameters that reflect the resource usage of cropping systems, and they are of special importance in the assessment of resource use efficiency and agro-ecosystem productivity. A number of studies have analysed WUE and RUE in wheat and maize in the NCP (Wang et al. Citation2001; Fang et al. Citation2006; Li et al. Citation2008; Qiu et al. Citation2008). These studies focused on WUE and RUE at the leaf level by monitoring the ratios of photosynthesis to transpiration and photosynthesis to intercepted photosynthetically active radiation (PAR), and at the field level by monitoring the ratios of biomass production to evapotranspiration and biomass production to intercepted photosynthetically active radiation. Few studies have described continuous canopy WUE and RUE due to limited measurement data (Campbell et al. Citation2001; Zhao et al. Citation2007). With the development of measurement techniques, the FluxNet network now provides a powerful platform for long-term and continuous measurements of carbon dioxide (CO2), water vapour and energy fluxes between various ecosystems and the atmosphere (Baldocchi et al. Citation2001).
In the NCP, continuous flux data have been measured at the Yucheng Comprehensive Experiment Station since 2002. These data enable a coupled analysis of CO2 and water fluxes and quantification of crop radiation and water use efficiency. Li et al. (Citation2006) studied the annual variation and control mechanism of carbon fluxes in farmland using data collected over two years. Zhao et al. (Citation2007) studied seasonal variations in the WUE of wheat and analysed the effects of weather variables on WUE. The results showed that photosynthetically active radiation and vapour pressure deficit (VPD) had a negative effect on WUE. However, few studies have assessed the difference between diurnal variations in WUE and RUE of irrigated wheat and maize fields in the NCP and analysed its control mechanism. Baldocchi (Citation1994) compared the WUE of a closed wheat canopy with that of an open maize canopy and found similar diurnal changes in wheat and maize. However, this conclusion may be not applicable to the NCP due to the different climatic background and crop growth season. Analysis of the difference helps to understand the mechanism of water and carbon dioxide exchange processes between C3 and C4 plants and the atmosphere at the canopy level. Simultaneously, mathematical models have been widely used to understand the biophysical and physiological processes of leaf and canopy photosynthesis and water use (Wang et al. Citation2007). Here, we used a coupled photosynthesis and transpiration model to analyse the response of WUE to the change in environment and crop factors.
Therefore, the objectives of this study were: first, to quantify the diurnal patterns of water, CO2 fluxes and resource use efficiency in the wheat-maize double cropping system in the NCP using continuous flux measurements; second, to compare the resource (radiation and water) use efficiencies of wheat and maize and analyse factors contributing to the diurnal variations; and third, to simulate the response of the diurnal variations in WUE to variable climate and crop factors.
Materials and methods
Site description
Field experiments were conducted from November 2002 to October 2003 at Yucheng Comprehensive Experiment Station in the NCP (36°40′ N, 116°22′ E, 28 m above sea level), which is a farmland site of ChinaFLUX. It has a warm-temperate, semi-humid and monsoon climate. The annual mean air temperature is 13.1°C and the mean temperatures in January and July are −3°C and 26.9°C, respectively. The mean annual precipitation is about 610 mm, and nearly 70% of it is concentrated in the summer months from June to August. The dominant soil type is a silt loam with a bulk density of 1.28 g cm−3 at 0–20 cm depth, 1.39 g cm−3 at 20–65 cm and 1.41 g cm−3 at 65–104 cm. The main cropping system in the region is a wheat-maize rotation with summer maize from late June to late September, followed by winter wheat from middle October to early June.
Experimental design and measurements
Winter wheat (variety Keyu 13) was planted on 15 October and harvested on 8 June. Summer maize (variety Ludan 981) was planted on 14 June and harvested on 27 September. During the wheat growth season, 225 mm of irrigation was applied using the surface flood method through plastic pipes and recorded with a water meter to prevent drought. Before planting winter wheat, 84.4 kg ha−1 of nitrogen (N) fertilizer as urea was applied and another 172.5 kg ha−1 of N fertilizer was applied in stem elongation. During the maize growth season, 281.3 kg ha−1 of N fertilizer was applied in stem extension. During the growing season of wheat and maize, 20 plants were randomly harvested at 5-day intervals. The leaf area index (LAI) was measured using an LI-COR 3100 instrument and crop height was measured using a metre ruler. Biomass samples were weighed after the material of each component had been dried in an oven at 80°C for 20 hours. Crops were harvested at maturity. More detailed descriptions of the experimental results are given in Li et al. (Citation2006).
An energy, water vapour and CO2 flux measurement system was set up in the field. The Eddy Covariance method was used to measure the CO2, sensible and latent heat fluxes at 2.1 m above the ground, and the height of eddy covariance system was adjusted during the growing season of maize. The system consisted of a fast-responding infrared gas analyser (LI-7500, LI-COR Inc, Lincoln, NE, USA) to measure CO2 flux and a three-dimensional sonic anemometer (CSAT-3, Campbell Scientific Inc, UT, USA) to measure sensible and latent heat fluxes. Data were recorded with a data-logger (CR23X, Campbell Scientific Inc, USA) and the sampling frequency was 20 Hz for each channel. Average values were calculated and recorded every 30 minutes. There was a fetch of more than 200 m for winds from all directions in the growing periods of winter wheat and maize.
A radiometer (CNR-1, KIPP&ZONEN, The Netherlands) was installed at a height of 1.86 m above the ground to measure downward and reflected components of shortwave and longwave radiation. Incident photosynthetically active radiation was measured at the same height with a quantum sensor (LI-190SB, LI-COR Co, USA). Air temperature and relative humidity were measured with temperature/humidity probes (HMP45C, VAISALA Co, Finland) and wind speed was measured with an anemometer (A100R, Vector Ins, England) at a height of 2.05 m above the ground.
Two heat flux plates (HFP01SC, Hukseflux, The Netherlands) were set 0.05 m below the ground at row and aisle positions to measure soil heat flux. Soil water content was measured with TDR sensors (CS615-L, Campbell Scientific Inc, USA) at soil depths of 0.01 m and 0.03 m, and soil temperatures were measured with five thermocouple sensors (TSAV, Campbell Scientific Inc, USA) at depths of 0.02, 0.05, 0.10, 0.20 and 0.50 m below the ground.
Data processing
Corrections were made in CO2 and latent heat flux measurements for density effects due to heat and water vapour transfer (Webb et al. Citation1980). To investigate the influence of the underlying terrain on fluxes, 3-D coordinate rotation was conducted during the growing seasons of winter wheat and summer maize. The threshold of friction velocity (u*) was determined according to the correlation between nocturnal CO2 flux and friction velocity (Falge et al. Citation2001) and nocturnal flux data were discarded when u*<0.15 m s−1. Moreover, abnormal flux data (such as data measured with Net Ecosystem Exchange [NEE] greater than 2.5 mg m2 s−1) as a result of instrument malfunction or inaccurate calibration of the system were discarded, as were flux data collected during the precipitation periods.
Calculation of canopy resource use efficiency
WUE (mg CO2 g H2O) was calculated as the ratio of net ecosystem productivity (NEP) and latent heat flux (LE):
Coupled photosynthesis and transpiration model
The framework of the coupled canopy photosynthesis and transpiration model was originally developed by Wang et al. (Citation2006), which included a canopy evapotranspiration module, a canopy photosynthesis module, and a soil water and heat transfer module. Several processes have been revised and simplified for this study as set out below.
Canopy evapotranspiration module
Two-layer water and heat transfer submodel The evapotranspiration was calculated by the S-W equation, which separates soil evaporation and plant transpiration into two layers (Shuttleworth and Wallace Citation1985).
The resistance submodel Aerodynamic resistance from the top of canopy to the reference height (z) was calculated for neutral conditions (Monteith & Unsworth Citation1990):
Aerodynamic resistance between the substrate and canopy source height was calculated by (Shuttleworth & Wallace Citation1985):
The soil resistance in the model is given in the form (Lin & Sun Citation1983):
Canopy photosynthesis module
Coupled photosynthesis and stomatal conductance model for C 3 and C 4 plants Collatz et al. (Citation1991) developed a coupled leaf photosynthesis and stomatal conductance model for C3 plants based on the biochemical model of leaf photosynthesis for C3 plants (Farquhar et al. Citation1980) and the leaf stomatal conductance model (Ball et al. Citation1987). Furthermore, Collatz et al. (Citation1992) proposed a coupled leaf photosynthesis and stomatal conductance model for C4 plants. In these models, the gross photosynthetic rate (P) is expressed as a function of intercellular partial pressure of CO2 (P i), the incident photosynthetic photon flux density (Q p) and leaf temperature (T l):
Because the transition from the limitation by one factor to the limitation by another appears to be gradual, to allow for some co-limitation between J c, J e and J s, Collatz et al. (Citation1991) solved the following quadratics for their smaller roots:
RuBP regeneration is controlled by the rate of electron transport/photo-phosphorylation which is given by (Collatz et al. Citation1991, Citation1992):
The capacity for the export or utilization of the products of photosynthesis in the case of C3 and the CO2 limited capacity for C4 photosynthesis are defined as (Collatz et al. Citation1991, Citation1992):
Dark respiration (R d) is proportional to V m (Collatz et al. Citation1991):
Stomatal conductance (g s) was calculated by the expression (Wang et al. Citation2007):
The main input data of the model included air temperature, water vapour pressure, wind speed, net radiation, soil heat flux, surface soil water content, averaged soil temperature at 0–10 cm depth, leaf area index and crop height. The main outputs were CO2 flux, soil evaporation, crop transpiration and canopy water use efficiency.
We verified the model by comparing the measured values and the simulated values of CO2 flux, latent heat flux and WUE. Sensitivity analyses were performed to see the response of the model to variable climate and crop factors.
Results
Diurnal variation of weather variables, carbon dioxide and water vapour fluxes
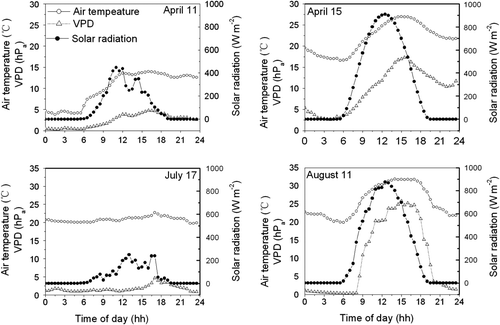
displays the diurnal variation of half-hour averaged solar radiation (Q), air temperature (T a) and VPD on typical clear and cloudy days over the winter wheat and summer maize growing seasons. On typical clear days, the lowest air temperature (10.4°C on 15 April and 11.9°C on 11 August) occurred at 6:00am, and increased over the daytime with solar radiation and reached the highest value (27°C on 15 April and 31.8°C on 11 August) at 3:00pm. The difference between the highest and the lowest values of VPD on 15 April and 11 August was 14.8 hPa and 25.5 hPa, respectively. On typical cloudy days, the diurnal variation in VPD was not as significant as on clear days. The difference between the highest and the lowest values of VPD was 4.5 hPa on 11 April and 3 hPa on 17 July, respectively.
Fig. 1 Diurnal variations of half-hourly averaged solar radiation (Q), air temperature (T a), vapour pressure deficit (VPD) on typical clear and cloud days over the growing season of winter wheat and summer maize.
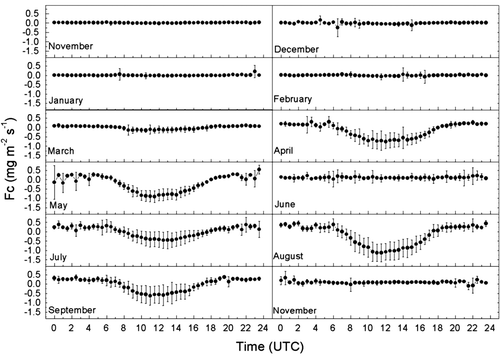
shows the monthly average diurnal variation of CO2 flux (F c) over the wheat-maize double-cropping field. At the early growth stage of winter wheat (November to February), F c was mainly soil respiration due to low crop LAI and canopy photosynthesis. Soil respiration was also very low because of low soil temperature. F c remained close to zero during the night-time and showed a small diurnal variation with a net CO2 release during the daytime. In the vigorous wheat growth stage (March to May), F c was positive during the night due to soil and dark respiration, and reached a maximum negative value between 11:00am and 12:00pm in accordance with incoming PAR. The minimum values of F c were −0.157, −0.756 and −0.911 mg CO2 m−2 s−1 in March, April and May, respectively. The diurnal variations of F c during the maize season were similar to those of winter wheat. F cwas positive during the early growth stage (after 25 June) and the post-growth stage (October). At the vigorous maize growth stage, the minimum values of F c were −0.454, −1.129 and −0.631 mg CO2 m−2 s−1 in July, August and September, respectively.
Fig. 2 Diurnal variations of monthly averaged half-hourly CO2 flux (F c) from November 2002 to October 2003 in a wheat-maize field at Yucheng. Bars indicate the standard deviation of particular half-hourly average values.
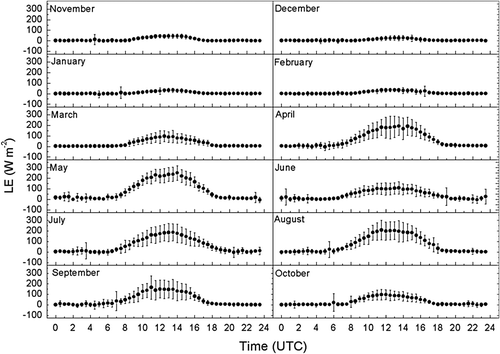
The monthly averaged half-hourly latent heat fluxes (LE) are shown in . From November to February, the diurnal variation range of LE was small because of the small daily changes in R n, T a and wind speed. With the increase in LAI, T a and R n from March onwards, LE showed a clear diurnal variation pattern, with a value close to zero in the night-time, and then increasing sharply after sunrise to a maximal value between 1:00pm and 3:00pm. On average, the maximum value of LE ranged from 2.4 MJ m−2 d−1 in December to 21.6 MJ m−2 d−1 in May and 18 MJ m−2 d−1 in August, corresponding to evapotranspiration rates of 1.0, 8.8 and 7.3 mm d−1, respectively.
Fig. 3 Diurnal variations of monthly averaged half-hourly latent heat flux (LE) from November 2002 to October 2003 in a wheat-maize field at Yucheng. Bars indicate the standard deviation of particular half-hourly average values.
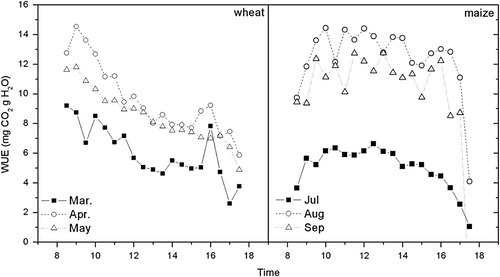
Diurnal variations in wheat and maize WUE
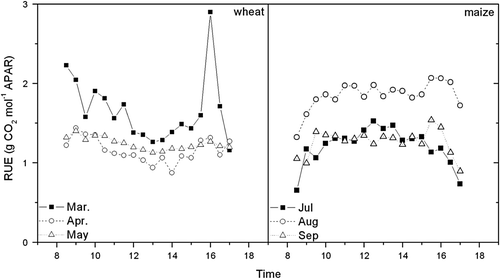
Monthly averaged diurnal changes in canopy WUE during the wheat season (March to May) and the maize season (July to September) are presented in . The WUE of wheat peaked near 9, 12 and 15 mg CO2 g H2O in the morning (8:00–9:00am) and then decreased with the increase in solar radiation and VPD. It then recovered in the late afternoon (4:00pm) before sunset in March, April and May. The WUE of maize increased after sunrise and retained a stable value of 6, 12 and 14 mg CO2 g H2O from mid-morning to mid-afternoon (10:00am–2:00pm) and then decreased slowly with time until sunset. The difference between the WUE of wheat and maize was caused by physiological differences in the response of canopy photosynthesis to environmental factors. PAR reached the light saturation point of wheat very soon after sunrise. High air temperature and VPD increased crop transpiration and led to a decline in stomatal conductivity, and therefore WUE decreased significantly. Because of maize's higher light saturation point and higher optimal temperature of canopy photosynthesis, the WUE of maize changed gently during the daytime. Baldocchi (Citation1994) found similar patterns of diurnal WUE variation in wheat and maize at Boardman, OR (45°40′ N, 119°40′ E). The diurnal WUE variation of wheat was close to the results of our study. But there was a significant difference in the WUE of maize between the two studies. Because the latitude of the study sites and water and fertilizer management practices were similar in the two studies, this difference was caused by the different development stage of maize. In Baldocchi's study, the maize was planted in April and measurements were taken in May (Julian Day #165), at which time the maize absorbed less PAR due to sparse canopy than in the growth season of maize in this study.
Fig. 4 The diurnal variations of monthly averaged WUE during the vigorous growth periods of wheat (March, April and May) and maize (July, August and September) from 2002 to 2003 at Yucheng.
Diurnal variation of RUE of wheat and maize
Monthly averaged diurnal changes of canopy RUE during the wheat season (March to May) and the maize season (July to September) are presented in . The RUE of wheat reached nearly 2.3, 1.5 and 1.4 g CO2 mol−1 APAR in the morning (8:00–9:00am) and then decreased with the increase in solar radiation. It reached its lowest value of the day around noon (12:00–1:00pm) and then increased with the decreasing PAR in March, April and May. However, the RUE of maize increased after sunrise and retained a stable value of 1.3, 1.9 and 1.3 g CO2 mol−1 PAR from mid-morning to mid-afternoon (10:00am–4:00pm). It then decreased slowly with time until sunset. The difference was caused by the different physiological response of canopy photosynthesis to environmental factors. The difference between the amplitude of the RUE of maize and wheat was due to the different photosynthetic pathways—C4 in maize and C3 in wheat (Tesfaye et al. Citation2006).
Fig. 5 Diurnal variations of monthly averaged radiation use efficiency (RUE) during the vigorous growth periods of wheat (March, April and May) and maize (July, August and September) from 2002 to 2003 in a wheat-maize field at Yucheng.
The three-month average RUE was 1.35 g CO2 mol−1 APAR for the wheat crop, corresponding to 1.76 g C MJ−1 APAR. For the maize crop it was 1.44 g CO2 mol−1 APAR, corresponding to 1.87 g C MJ−1 APAR. These results were similar to those of other studies (Sinclair and Muchow Citation1999).
The relationship of WUE in wheat and maize to PAR and VPD
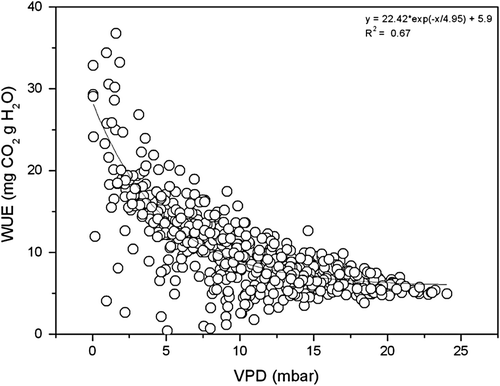
shows the relationship between the WUE of wheat and VPD on typical clear days. WUE decreased exponentially with increasing VPD and retained a stable low value when VPD was above 1.5 kPa, suggesting that VPD was the control factor for diurnal WUE variation on typical clear days.
Fig. 6 The relationship between VPD and WUE of wheat on typical clear days during the growth season of wheat at Yucheng.
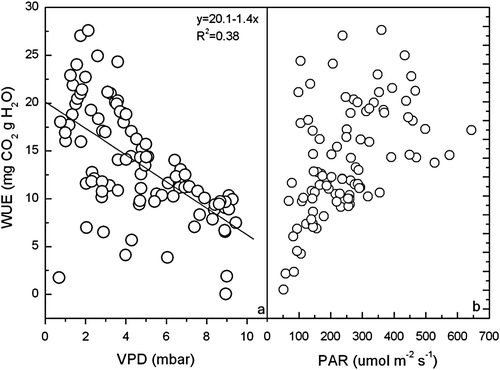
The WUE of wheat decreased linearly with increasing VPD on cloudy days but the R2 value was only 0.38 (a). b shows that the variation of WUE was also sensitive to light intensity on cloudy days. WUE increased with increasing light intensity when PAR was lower than 300 µmol m−2 s−1.
Fig. 7 The relationship between VPD, PAR and WUE of wheat on typical cloudy days during the growth season of wheat at Yucheng.
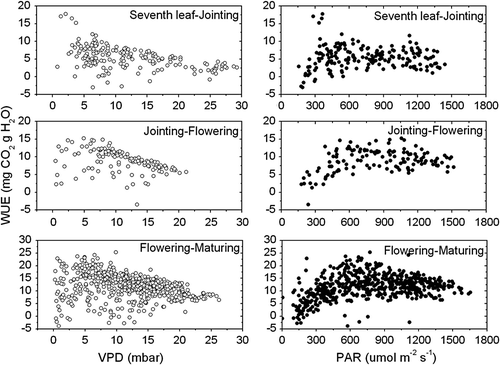
presents the relationship between the WUE of maize and PAR and VPD on typical clear days during three development stages of maize. The WUE of maize increased with light intensity when PAR was lower than 600 µmol m−2 s−1. The variation in the WUE of maize was relatively insensitive to VPD on typical clear days, which was consistent with the finding by Baldocchi (Citation1994). This result shows that other environmental factors also played significant roles in determining the diurnal variation of canopy WUE of maize. Higher optimal temperature and light saturation point of canopy photosynthesis for C4 crop (maize) than C3 crop (wheat) may lead to the difference.
Fig. 8 The relationship between PAR, VPD and WUE on typical clear days during the growing season of maize at Yucheng.
Test and sensitivity analysis of the model
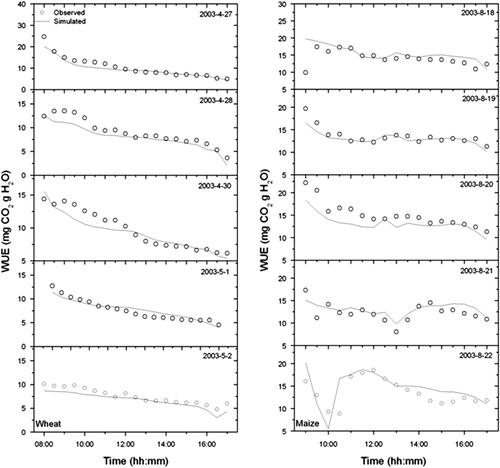
The comparisons between simulated and measured half-hourly averaged CO2 flux, latent heat flux and WUE on typical clear days during the growing season of winter wheat and summer maize are shown in and . The observed and the simulated CO2 flux, latent heat flux and WUE are in good agreement indicating that the model could effectively simulate diurnal variation of water, carbon fluxes and WUE in wheat-maize cropping system.
Fig. 9 Comparison between simulated and observed values of CO2 flux and latent heat flux (LE) over winter wheat and summer maize canopy at typical clear days in 2003.
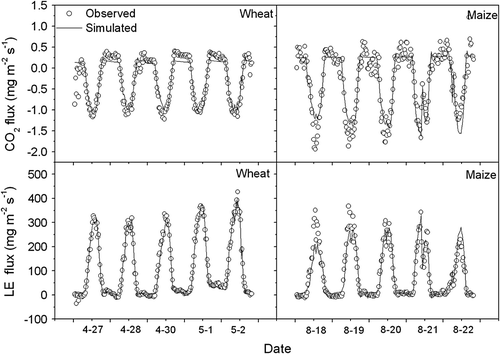
Fig. 10 Comparison between simulated and observed values of WUE over winter wheat and summer maize canopy at typical clear days in 2003.
shows the simulated response of CO2 flux, LE and WUE of winter wheat and summer maize to change in air temperature (5–40°C), PAR (100–1900 µmol m−2 s−1) and wind speed (0–16 m s−1) when other weather variables were unchanged.
Fig. 11 The simulated response of CO2 flux, latent heat flux (LE), WUE of winter wheat and summer maize to the change in air temperature (5–40°C) at net radiation 200 W m−2, relative humidity 60% and wind speed 2 m s−1, in PAR (100–1900 µmol m−2 s−1) at air temperature 10°C, relative humidity 60% and wind speed 2 m s−1, and wind speed (0–16 m s−1) at air temperature 10°C, net radiation 200 W m−2, relative humidity 60%.
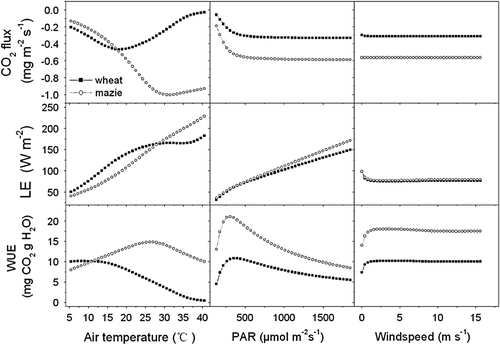
The response characteristics of WUE to the variation in PAR and wind speed between wheat and maize were similar. However, the response of canopy photosynthesis of wheat and maize to air temperature was significantly different. Air temperature affects canopy photosynthesis in two ways: one effect is on the intrinsic speed of the biochemical process of photosynthesis, and the other is on vapour pressure deficit (Wang et al. Citation2006). The optimum temperature of maize photosynthesis was higher than wheat photosynthesis, which resulted in a different response curve of WUE to air temperature. The WUE of wheat decreased with the increase in air temperature. However, the WUE of maize increased with the increase in air temperature and reached the maximal value when the air temperature was about 28°C, and then decreased with increasing air temperature.
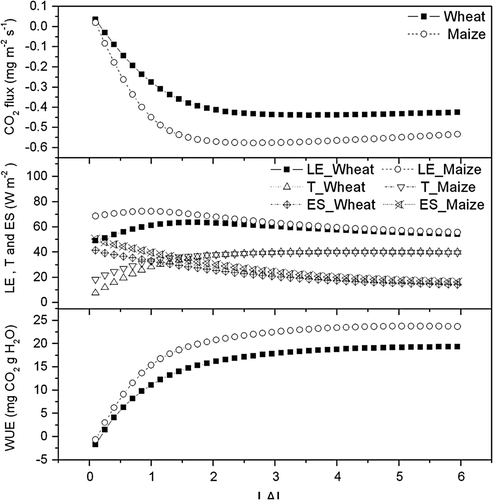
shows the simulated response of CO2 flux, LE, crop transpiration (T), soil evaporation (ES) and WUE of winter wheat and summer maize to the variation in LAI (0–6) when other input variables were held unchanged. An increase in LAI resulted in a nearly linear increase in canopy photosynthesis especially when LAI was low. But the change in canopy evapotranspiration with increasing LAI was not significant compared with canopy CO2 flux. Thus, WUE increased when LAI increased, which suggests that reducing soil evaporation at the early stage of wheat and maize would improve water use efficiency (Deng et al. Citation2003; Zhang et al. Citation2005).
Fig. 12 The simulated response of CO2 flux, latent heat flux (LE), crop transpiration (T), soil evaporation (ES), WUE of winter wheat and summer maize to the change in leaf area index (LAI) (0–6) at net radiation 200 W m−2, relative humidity 60% and wind speed 2 m s−1, air temperature10°C.
Discussion and conclusion
The NCP is one of the most important agricultural production areas in China. However, high yield in the NCP greatly depends on large amounts of irrigation, 64% of which comes from groundwater (Yang and Zehnder Citation2001). The NCP has been one of the most developed agricultural irrigation areas (Geng et al. Citation2001). Over-irrigation has led to a rapid decline in the groundwater table and water shortages (Zhang et al. Citation2005). Improving resource use efficiency by breeding new cultivars and applying agronomic technologies is an urgent task to ensure the stability of agricultural production in the NCP. The average WUE of the wheat crop in this study was 8.05 mg CO2 g−1 H2O and it was 9.22 mg CO2 g−1 H2O for the maize crop, which was lower than the results estimated from measured grain yield and actual evapotranspiration measured by large weighing lysimeter or remote-sensing technology in other regions of the NCP (Mo et al. Citation2005; Zhang et al. Citation2005). The main reason was that a great amount of irrigation applied in the wheat-growing season reduced the WUE. In addition, many field cultivation management practices for high yield and high WUE were not used in the experiment field in this study, such as selecting high-yield cultivars, optimizing irrigation scheduling and reducing soil evaporation by straw mulching (Zhang et al. Citation2005). Therefore, there is great potential to save water to increase WUE in the study region. shows the comparison between the water consumption of wheat and maize under different WUE levels in the production of 4965 kg ha−1 wheat and 5121 kg ha−1 maize, which were average yields of wheat and maize from 2000 to 2008 in the North China Plain. Rainfall ranges from 470–910 mm from the north to the south in the NCP, and about 70% is concentrated in the growth period of maize. If the WUE of wheat and maize reaches 1.5 and 2 kg m−3, respectively, wheat would require about 58–150 mm water from the south to the north, while rainfall could meet the water demands of maize in most years.
Table 1 The comparison between water consumption of wheat and maize under different WUE levels in the production of 4965 kg ha−1 wheat and 5121 kg ha−1 maize
Our study assumed that the extinction coefficient was a constant during the whole growth season of wheat and maize. However, the extinction coefficient changed with the development stages of wheat and maize. In addition, much research has shown that planting patterns have a significant influence on the RUE due to the variation in the vertical distributions of LAI (Han et al. Citation2008; Li et al. Citation2008). Therefore, more integrated models including robust radiation transfer processes over the canopy are required to analyse the control mechanism of diurnal and seasonal variation of RUE.
In summary, this study provides useful data that quantify the diurnal dynamics of WUE and RUE in a wheat-maize double-cropping field in the NCP. The results can assist in the analysis of resource use efficiency and productivity of agro-ecosystems. The results of this study are also helpful in the estimation of daily water use and radiation use efficiencies in crop growth models and for extrapolating instantaneous remote-sensing measurements to a daily scale.
Acknowledgements
This work is supported by a Special Fund for the Agriculture Profession (200803030) and National Basic Research Program of China (‘973’ Project, 2005CB121106) and National Basic Research Program of China (‘973’ Project, 2009CB118608). The authors acknowledge the anonymous referees for their valuable comments.
References
- Baldocchi , D . 1994 . A comparative study of mass and energy exchange rates over a closed C3 (wheat) and an open C4 (corn) crop: II CO2 exchange and water use efficiency . Agricultural and Forest Meteorology , 67 : 291 – 321 .
- Baldocchi , D , Falge , E , Gu , LH , Olson , R , Hollinger , D , Running , S , Anthoni , P , Bernhofer , C , Davis , K , Evans , R , Fuentes , J , Goldstein , A , Katul , G , Law , B , Lee , XH , Malhi , Y , Meyers , T , Munger , W , Oechel , WUKTP , Pilegaard , K , Schmid , HP , Valentini , R , Verma , S , Vesala , T , Wilson , K and Wofsy , S . 2001 . FLUXNET: a new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities . Bulletin American Meteorology Society , 82 : 2415 – 2434 .
- Ball JT , Woodrow IE , Berry JA , 1987 . A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions . In: J Biggins Progress in photosynthesis research . Dordrecht-Boston-Lancaster , Nijhoff . 4 : 221 227 .
- Campbell , CS , Heilman , JL , McInnes , KJ , Wilson , LT , Medley , JC , Wu , G and Cobos , DR . 2001 . Seasonal variation in radiation use efficiency of irrigated rice . Agricultural and Forest Meteorology , 110 : 45 – 54 .
- Choudhury , BJ and Monteith , JL . 1988 . A four-layer model for the heat budget of homogenous land surfaces . Quarterly Journal of Royal Meteorological Society , 114 : 373 – 398 .
- Collatz , GJ , Ball , JT , Grivet , C and Berry , JA . 1991 . Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: A model that includes a laminar boundary layer . Agricultural and Forest Meteorology , 54 : 107 – 136 .
- Collatz , GJ , Ribas-Carbo , M and Berry , JA . 1992 . Coupled photosynthesis-stomatal conductance model for leaves of C4 plants . Australia Journal of Plant Physiology , 19 : 519 – 538 .
- Deng , XP , Shan , L , Kang , SZ , Inanaga , S and Mohanmed , EK . 2003 . Improvement of wheat water use efficiency in semiarid area of China . Agricultural Sciences in China , 2 : 35 – 44 .
- Falge , E , Baldocchi , D , Olson , R , Anthoni , P , Aubinet , M , Bernhofer , C , Burba , G. , Ceulemans , R , Clement , R , Dolman , H , Granier , A , Gross , P , Grunwald , T , Hollinger , D , Jensen , NO , Katul , G , Keronen , P , Kowalski , A , Lai , CT , Law , BE , Meyers , T , Moncrieff , H , Moors , E , Munger , JW , Pilegaard , K , Rannik , U , Rebmann , C , Suyker , A , Tenhunen , J , Tu , K , Verma , S , Vesala , T , Wilson , K and Wofsy , S . 2001 . Gap filling strategies for defensible annual sums of net ecosystem exchange . Agricultural and Forest Meteorology , 107 : 43 – 69 .
- Fang , QX , Chen , YH , Li , QQ , Yu , SZ , Luo , Y , Yu , Q and Ouyang , Z . 2006 . Effects of soil moisture on radiation utilization during late growth stages and water use efficiency of winter wheat . Acta Agronomic Sinica , 32 : 861 – 866 .
- Farahani HJ , Bausch WC 1994 . Seasonal application of physical evapotranspiration models-resistance estimation . ASAE Paper No 94-2547 , ASAE , St Joseph, MI .
- Farquhar , GD , von Caemmerer , S and Berry , JA . 1980 . A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species . Planta , 149 : 78 – 90 .
- Geng , S , Zhou , YX , Zhang , MH and Smallwood , KS . 2001 . A sustainable agro-ecological solution to water shortage in the North China Plain (Huabei Plain) . Journal of Environmental Planning and Management , 44 : 345 – 355 .
- Han , H , Li , Z , Ning , T , Zhang , X , Shan , Y and Bai , M . 2008 . Radiation use efficiency and yield of winter wheat under deficit irritation in North China . Plant Soil Environment , 54 : 313 – 319 .
- Lee , XH and Hu , XZ . 2002 . Forest-air fluxes of carbon water and energy over non-flat terrain . Boundary-Layer Meteorology , 103 : 277 – 301 .
- Li , J , Yu , Q , Sun , XM , Tong , XJ , Ren , CY , Wang , J , Liu , EM , Zhu , ZL and Yu , GR . 2006 . Carbon dioxide exchange and the mechanism of environmental control in a farmland ecosystem in North China Plain . Science in China Series D: Earth Science , 49 ( suppl. II ) : 226 – 240 .
- Li , Q , Chen , Y , Liu , M , Zhou , X , Yu , S and Dong , B . 2008 . Effects of irrigation and planting patterns on radiation use efficiency and yield of winter wheat in North China . Agricultural Water Management , 95 : 469 – 476 .
- Lin JD , Sun SF 1983 . Moisture and heat flow in soil and their effects on bare soil evaporation . Transaction of Water Conservancy 7 : 1 7 (in Chinese) .
- Mehrez , BM , Taconet , O , Vidal-Madjar , D and Valencogne , C . 1992 . Estimation of stomatal resistance and canopy evaporation during the Hapex-Mobilhy experiment . Agricultural Forest Meteorology , 58 : 285 – 313 .
- Mo , X , Liu , S , Lin , Z , Xu , Y , Xiang , Y and McVicar , TR . 2005 . Prediction of crop yield, water consumption and water use efficiency with a SVAT-crop growth model using remotely sensed data on the North China Plain . Ecological Modelling , 183 : 301 – 322 .
- Monteith , JL and Unsworth , MH . 1990 . Principles of environmental physics , London : Edward Arnold .
- Olesen , JE , Jorgensen , LN and Mortensen , JV . 2000 . Irrigation strategy, nitrogen and application and fungicide control in winter wheat on a sandy soil. II. Radiation interception and conversion . Journal of Agricultural Science , 134 : 13 – 23 .
- Qiu , GY , Wang , L , He , X , Zhang , X , Chen , S , Chen , J and Yang , Y . 2008 . Water use efficiency and evapotranspiration of winter wheat and its response to irrigation regime in the North China Plain . Agricultural and Forest Meteorology , 148 : 1848 – 1859 .
- Shaw RH , Pereira AR 1981 . Aerodynamic roughness of vegetated surfaces. The effect of canopy structure and density . 15th Conference of Agriculture and Forest Meteorology and 50th Conference on Biometeorology , 1–3 April 1981 , Anaheim, California , American Meteorological Society .
- Shuttleworth , WJ and Gurney , RJ . 1990 . The theoretical relationship between foliage temperature and canopy resistance in a sparse crop . Quarterly Journal of Royal Meteorological Socienty , 116 : 498 – 519 .
- Shuttleworth , WJ and Wallace , JS . 1985 . Evaporation from sparse crops—an energy combination theory . Quarterly Journal of Royal Meteorological Society , 111 : 839 – 855 .
- Sinclair , TR and Muchow , RC . 1999 . Radiation use efficiency . Advances in Agronomy , 65 : 215 – 265 .
- State Statistics Bureau 1999 . Statistics yearbook of China . Beijing , Statistics Publishing House .
- Tesfaye , K , Walker , S and Tsubo , M . 2006 . Radiation interception and radiation use efficiency of three grain legumes under water deficit conditions in a semi-arid environment . European Journal of Agronomy , 25 : 60 – 70 .
- Wang , E , Yu , Q , Wu , D and Xia , J . 2008 . Climate, agricultural production and hydrological balance in the North China Plain . International Journal of Climatology , 28 : 1959 – 1970 .
- Wang , HX , Zhang , L , Dawes , WR and Liu , CM . 2001 . Improving water use efficiency of irrigated crops in the North China Plain –measurements and modeling . Agricultural Water Management , 48 : 151 – 167 .
- Wang , J , Yu , Q and Lee , X . 2007 . Simulation of crop growth and energy and carbon dioxide fluxes at different time steps from hourly to daily . Hydrological Processes , 21 : 2474 – 2492 .
- Wang , J , Yu , Q , Li , J , Li , L , Li , X , Yu , G and Sun , X . 2006 . Simulation of diurnal variations of CO2, water and heat fluxes over winter wheat with a model coupled photosynthesis and transpiration . Agricultural Forest Meteorology , 137 : 194 – 219 .
- Webb , EK , Pearman , G and Leuing , R . 1980 . Correction of flux measurements for density effects due to heat and water vapor transfer . Quarterly Journal of Royal Meteorological Society , 106 : 85 – 100 .
- Wu , DR , Yu , Q , Lu , CH and Hengsdijk , H . 2006 . Quantifying production potentials of winter wheat in the North China Plain . European Journal of Agronomy , 24 : 226 – 235 .
- Yang , H and Zehnder , A . 2001 . China's regional water scarcity and implications for grain supply and trade . Environment and Planning A , 33 : 79 – 95 .
- Yu , Q , Xu , S , Wang , J and Lee , X . 2007 . Influence of leaf water potential on diurnal changes in CO2 and water vapour fluxes . Boundary-Layer Meteorology , 124 : 161 – 181 .
- Zhang , XY , Chen , SY , Liu , MY , Pei , D and Sun , HY . 2005 . Improved water use efficiency associated with cultivars and agronomic management in the North China Plain . Agronomy Journal , 97 : 783 – 790 .
- Zhao , FH , Yu , GR , Li , SG , Ren , CY , Sun , XM , Mi , N , Li , J and Ouyang , Z . 2007 . Canopy water use efficiency of winter wheat in the North China Plain . Agricultural Water Management , 93 : 99 – 108 .