Abstract
Photosynthetic characteristics and resource utilization efficiency of maize (Zea mays L.) and millet (Setaria italica L.) are studied in a loess region of China. Limited sampling shows that maize has a higher light-saturated CO2 assimilation rate, apparent quantum yield and carboxylation efficiency than does millet in the middle (July) and late (September) growth stage. Compared with millet, maize has lower transpiration rate (T r ) and stomatal conductance (G s ) in July and higher T r and G s in September. This leads to higher leaf water use efficiency in maize in July, but it is lower than for millet in September. Mass-based leaf nitrogen concentrations (N mass) of both crops are similar in both growth stages. Leaf mass per area (LMA) of maize is the same as of millet in July but lower in September. These results indicate differences in photosynthetic capacity and resource use efficiencies between these crops.
Introduction
The yield potential of a crop depends on its capacity to obtain resources from the environment and to use these resources to fix carbon dioxide (CO2) into biomass. For grain crops, this also depends on the proportions of its biomass partitioned to grain (Binkley et al. Citation2004; Long et al. Citation2006). Photosynthesis is the basis for crop growth and yield formation: the efficiency of converting resources into biomass depends on the net leaf photosynthetic rate of the crop canopy. Although some research has indicated that leaf photosynthetic rate is poorly correlated with crop yield (Evans Citation1998), more recent experiments have provided evidence that a sustained increase in leaf photosynthesis leads to an increase in crop yield (Ainsworth et al. Citation2002; Ainsworth & Long Citation2005).
Leaf photosynthetic efficiency mainly depends on simultaneous supplies of light, CO2, water and nitrogen (N). For most field crops, the low supply of resources frequently constrains leaf photosynthesis, thus lowering crop productivity. Drought and nitrogen deficiency are major constraints to crop production and yield stability in many arid and semi-arid regions (Jamieson et al. Citation1995; Shangguan et al. Citation2000, Citation2004; Hu et al. Citation2006; Shao et al. Citation2008). Desirable traits of crops in drought-susceptible and nitrogen-limited environments include efficient use of limited resources (Shangguan et al. Citation2000). Water use efficiency provides an indicator of the performance of a crop growing under any environmental constraint. An improvement in water use efficiency not only improves crop yield and profitability but also reduces environmental impacts on groundwater recharge (Sadras Citation2004). Efficient photosynthetic nitrogen use is believed to increase the fitness of plants because high photosynthetic nitrogen use efficiency (PNUE) is associated with high relative growth rate (Hikosaka Citation2004). Low soil nitrogen can limit both the leaf area intercepting light and the capacity of plants to fix CO2 photosynthetically (Reich et al. Citation2006). Improved crop water and nitrogen use efficiencies can therefore reduce the detrimental effects of drought and low soil nitrogen on leaf photosynthesis and crop yield.
Maize and millet have been extensively planted in the semi-arid Loess Plateau of China because of their high tolerance for drought and nitrogen deficiencies. Many studies have evaluated the effects of water and fertilizer inputs and their interactions on grain yield of the two crops there (Dang et al. Citation2003; Yan et al. Citation2006; Liu & Zhang Citation2007). Other research has investigated the water physiology and photosynthetic characteristics of maize and millet (Cheng et al. Citation1996; Kang et al. Citation1998; Liu et al. Citation2004; Shi et al. Citation2007). This research indicates that within a certain range, either irrigation or fertilizer markedly raise grain yield of the two crops as well as promote their resource use efficiency. By contrast, drought increases leaf stomatal resistance, decreases leaf water potential, transpiration rate and photosynthetic rate, thus reducing grain yield. Previous studies help us better understand how to control water and fertilizer to increase crop yield. However, mechanisms regarding the physio-ecological adaptations of maize and millet in this region have been poorly documented. Genetic differences between the two crop species would suggest different utilization efficiencies of light, CO2, water and nitrogen, which further determine their adaptability and reflect their adaptation strategies to the surrounding environment.
The objectives of this study were first, to examine the differences in the efficiency of resource use (light, CO2, water and nitrogen) of maize and millet and, second, to better understand the resource utilization strategies and physio-ecological adaptation mechanisms of the two crops.
Materials and methods
Location and climatic conditions
The study was conducted in the Kangjiagelao watershed (109°31′48″–109°32′31″E, 36°30′51″–36°31′09″N) of Yan'an, Shaanxi, China. The catchment area covers about 4 km2 with an altitude ranging from 1100–1320 m above sea level. It has a semi-arid continental climate with a mean annual precipitation of 550 mm. Most of the precipitation occurs from June to September. The average annual temperature is about 8–10°C and annual evaporation is about 900–1000 mm. The annual accumulated temperature above 10°C is about 3200°C and annual solar radiation is about 5000 MJ m−2 a−1. The frost-free period is 186 days and average annual sunshine amounts to 2445 h (Liu et al. Citation2003).
Study site
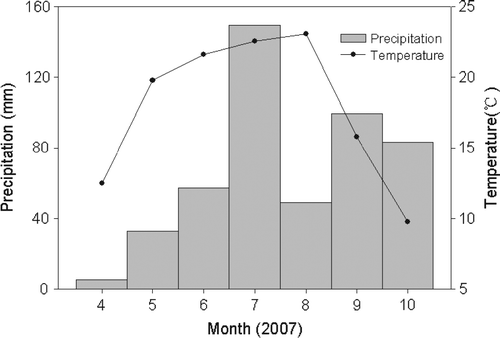
The experiment was conducted during the crop growing season from April to October 2007. Maize (Zea mays L. cv. Zhongdan 2) and millet (Setaria italica L. Beauv cv. Jinfen 7) were planted in two adjacent pieces of terraced farmland to ensure identical moisture and temperature conditions. The soil of the farmland to a depth of 40 cm was fine-loamy with an average total N content of 0.65 g kg−1, an average total phosphorus (P) content of 0.42 g kg−1 and an average organic matter content of 5.62 g kg−1. Maize was sown at about 28,000 plants per hectare on 20 April and was harvested on 5 October. Millet was planted at about 167,000 plants per hectare on 21 April and was harvested on 8 October. Crops were neither fertilized nor irrigated during the growth period. shows the distribution of rainfall and temperatures from April to October in the study area: rainfall was relatively plentiful during the 2007 growing season.
Fig. 1 Distribution of precipitation and temperature from April to October 2007 in the Yangou watershed
Photosynthetic gas exchange
We randomly selected three evenly grown maize and three evenly grown millet plants and then chose a healthy, fully grown, south-facing leaf at the upper-middle part of each plant for measurements. All measurements were taken between 8:30 a.m. and 11:30 a.m. on 24–25 July and 20–21 September 2007, which were sunny days.
Photosynthetic gas exchange parameters of both crops were measured with an open-system portable CO2/H2O analyser (LI-6400, LI-COR Inc, Lincoln, NE, USA) at middle (July) and late (September) growth stage in 2007. Photosynthesis light response curves were measured at 13 light intensities ranging from 2000–0 µmol m−2 s−1 with a CO2 concentration of 380 µmol mol−1 and a temperature of 30°C in the chamber. The responses of A (CO2 assimilation rate) to C i (intercellular CO2 concentration) were assessed in terms of the concentration of CO2 entering the leaf chamber: this varied from 1500–50 µmol mol−1 at a light intensity (cf. photosynthetically available radiation, PAR) of 1500 µmol m−2 s−1 and a temperature of 30°C.
The curve fitting application (Sigmaplot for Windows 10.0, Systat Software Inc, San Jose, CA, USA) was adopted to analyse the A-C i and A-PAR responses by the following three-component exponential function (Watling et al. Citation2000):
The parameters P n (photosynthetic rate) and T r (transpiration rate) as well as Gs (stomatal conductance) were also measured with the LI-6400 (LI-COR Inc, Lincoln, NE, USA). The WUE (water use efficiency) of the two crops was calculated as the ratio of P n to T r (Penuelas et al. Citation1998). After measurements were completed, the leaves were collected and used to determine LMA (leaf mass per area) and N mass (mass-based leaf nitrogen concentration).
LMA and Nmass
The sample leaves were cut into several small pieces, 5–10 leaf pieces were spread on a transparent rectangular plastic paper (21 × 29.7 cm) without overlapping. The plastic paper was then optically scanned at 200 dpi by a flatbed scanner (BenQ 5000 scanner, BENQ Inc, Taiwan, China). The images were recorded in tiff format and analysed using DELTA-T SCAN image analysis software (Delta-T Scan, Delta-T Devices Company, UK). Thus, the projected leaf area (cm2) could be quickly calculated. Each measurement was replicated five times. The scanner and software were calibrated with DELTA-T SCAN image standards for length and area before use. The scanned leaves were collected and placed in envelopes and oven-dried at 70°C for at least 48 h to constant weight. They were then weighed on a scale with a precision of <0.001 g, and LMA (g m−2) was calculated as the ratio of leaf dry weight to leaf area.
After LMA determination, the dried leaf samples were used for N mass determination. The samples were ground into a uniformly fine powder with a plant sample mill and then sieved with a 1 mm mesh screen before chemical determination. A 200 mg sample was used to determine the leaf nitrogen content by modified Kjeldahl analysis in which the ratio of concentrated sulphuric and salicylic acid with Na2SO4, K2SO4 and Se as catalysts was 62:1:1 (m m−1) (Page et al. Citation1982). The digestion products were used to determine N mass with a Kjeltec analyser (Kjeltec 2300 Analyzer Unit, Foss, Sweden). PNUE (µmol CO2 mol−1 s−1) was determined as P n / (N mass/14 × LMA) (Poorter and Evans Citation1998).
Data analysis
A one-way analysis of variance (ANOVA) was conducted with the SPSS 13.0 statistical package (SPSS 13.0 for Windows, Chicago, USA). The LSD test was used for multiple comparisons.
Results
Light utilization efficiency of maize and millet at different growth stages
In our limited sampling, we found values of A sat and AQY to be significantly higher in maize than in millet at the two growth stages. Compared with millet, the A sat of maize was 18.6% higher in July and 61.6% higher in September; the AQY of maize was 12.7% and 20% higher than that of millet in July and September, respectively. A sat and AQY in both crops were substantially higher in July than in September. Maize A sat and AQY decreased by 49.2% and 23.9%, respectively, whereas millet A sat and AQY decreased by 62.8% and 28.6%, respectively, over that period ().
Table 1 A sat, AQY and CE of maize and millet at two growth stages. Values are means ±SD (n = 3). Different letters in the same column indicate significant differences at P < 0.05 according to LSD tests
CO2 utilization efficiency of maize and millet at different growth stages
The CE of maize was 86.1% higher than that of millet in July and 36% higher in September. However, the CE of maize was 55.7% less in September than in July, a decrease that is greater than was the difference from millet ().
WUE and PNUE of maize and millet at different growth stages
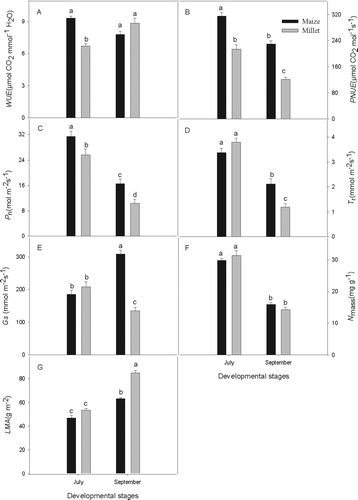
The WUE of maize was 1.4 times greater than that of millet in July (P < 0.01) but was not significantly different from that of millet in September (A). The PNUE of maize and millet differed significantly in both July and September (P < 0.01), that of maize being 1.5 and 1.9 times that of millet in July and September, respectively (B).
Fig. 2 WUE, PNUE, P n, T r, Gs, LMA and N mass of the maize and millet at the different growth stages. Values are means ±SD (n = 3). The same letters are not significantly different at P < 0.05 by LSD test
Discussion
The values of A sat, AQY and CE reflect potential plant photosynthetic capacity, energy use efficiency and CO2 use efficiency, respectively. Carbon assimilation capacity and effective energy capture efficiency greatly influence crop yield (Murchie et al. Citation1999). In our limited study, both maize and millet had high A sat, AQY and CE values at the middle growth stage; these were remarkably low at the late growth stage (). In maize, A sat, AQY and CE were higher than in millet in both growth stages, indicating that maize used light and CO2 more efficiently to reach a higher photosynthetic rate. The high carbon assimilation rate in maize could result in higher yields than in millet.
Both maize and millet had high WUE in both growth stages (A), reflecting a relatively high P n and low T r . To obtain a higher P n , plants must maintain a certain stomatal aperture, but any increase in stomatal aperture can lead to greater stomatal water loss. Plants can also increase WUE by simultaneously having high P n and low T r as we found to be the case in July for maize (it simultaneously had low Gs also) but not millet. In September, maize still had higher P n than millet, but higher Gs led to higher T r , resulting in slightly lower WUE than millet (A, 2C, 2D, 2E).
The parameter of PNUE is the ratio of the CO2 assimilation rate to leaf nitrogen content and represents the economy of leaf photosynthesis (Hikosaka Citation2004). Previous research has revealed significant differences in the relationship of CO2 assimilation rate and leaf nitrogen content between species. The observed PNUE tends to be higher in C4 species than in C3 plants (Anten et al. Citation1995), higher in herbaceous than in woody species (Hikosaka et al. Citation1998) and higher in deciduous than in evergreen species (Takashima et al. Citation2004). Generally speaking, species with a high PNUE tend to have higher growth rates and higher productivity than those with a low PNUE (Hikosaka Citation2004; Zheng & Shangguan Citation2007a). In our study, maize had much higher PNUE than millet in the two growth stages, indicating stronger resource capture and utilization capacities, especially with regard to nitrogen use.
The proportion of nitrogen distributed between leaf photosynthetic organs and non-photosynthetic organs indicates a plant's strategy of increasing PNUE to improve photosynthetic efficiency versus increasing LMA to enhance defence capabilities. Higher LMA means that plants allocate more carbon and nitrogen to cell wall components or other non-photosynthetic apparatus: this results in a low nitrogen content in photosynthetic organs, thus lower photosynthetic capacity and lower PNUE (Takashima et al. Citation2004; Warren & Adams Citation2004). In our study, maize had significantly higher P n than did millet at the two growth stages, but they had similar LMA and N mass in July (C, 2F, 2G). This indicates that higher P n was the main reason for PNUE being higher in maize than in millet at the middle growth stage. Although there was little difference in N mass between maize and millet in September, the P n of maize was significantly higher and the LMA of maize was substantially lower than of millet at this late growth stage—the combination of high P n and low LMA resulting in higher PNUE in maize than in millet.
Resource utilization efficiency is the capacity of a plant to capture and use light, warmth, water, soil nutrition, etc. during growth. Plants with high resource use efficiency often have high productivity and are able to adapt well to environmental conditions—in our case, enabling maize to have higher yields than millet in suitable conditions.
In addition to inherent genetic factors, the environment of a plant also greatly affects its resource use efficiency. For example, P n, WUE and PNUE of Robinia pseudoacacia were significantly higher than those of Pinus tabulaeformis in humid areas of the Loess Plateau; however, with decreasing rainfall, P n, WUE and PNUE of P. tabulaeformis were close to or higher than those of R. pseudoacacia (Zheng & Shangguan Citation2007b). So R. pseudoacacia may have a higher photosynthetic capacity only in favourable environments.
Similarly, to enable the high resource utilization by maize, appropriate surrounding conditions are needed. The arid and changeable climate of the Loess Plateau may create difficulties for maize to maintain sustainable and efficient resource utilization and stable and high yields. By contrast with the introduced maize, millet is a native species in the Loess Plateau and has long been cultivated here: it is thus likely to have evolved to be better adapted to the climate and environment of this region. The higher LMA of millet at the two growth stages indicates that millet is better than maize at tolerating stress. Therefore, stable yields of millet can be produced even in an extreme drought year. Zhang (Citation2007) showed that millet had modest inter-annual variation in terms of yield compared with maize. This reliability probably is the main reason why millet has been cultivated so widely in this region despite its lower yield.
Acknowledgements
This study was funded by the Major State Basic Research Development Projects of China (2007)CB407200), the Knowledge Innovation Project in Chinese Academy of Sciences (KSCX1-YW-12), the State Forestry Administration of China (2009)04056) and the Program for Changjiang Scholars and Innovative Research Team in University (IRT0749). Special thanks are also given to reviewers and editors for their valuable comments.
References
- Ainsworth , EA , Davey , PA , Bernacchi , CJ , Dermody , OC , Heaton , EA , Moore , DJ , Morgan , PB , Naidu , SL , Ra , HSY and Zhu , XG . 2002 . A meta-analysis of elevated CO2 effects on soybean (Glycine max) physiology, growth and yield . Global Change Biology , 8 ( 8 ) : 695 – 709 .
- Ainsworth , EA and Long , SP . 2005 . What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2 . New Phytologist , 165 ( 2 ) : 351 – 371 .
- Anten , NPR , Schieving , F and Werger , MJA . 1995 . Patterns of light and nitrogen distribution in relation to whole canopy gain in C3 and C4 mono- and dicotyledonous species . Oecologia , 101 ( 4 ) : 504 – 513 .
- Binkley , D , Stape , JL and Ryan , MG . 2004 . Thinking about efficiency of resource use in forests . Forest Ecology and Management , 193 ( 1–2 ) : 5 – 16 .
- Cheng , LM , Yan , JY , Zhang , YG and Zhang , H . 1996 . Study on drought resistant physiological characteristics of foxtail millet under moisture stress condition . Chinese Bulletin of Botany , 13 ( 3 ) : 56 – 58 .
- Dang , TH , Hao , MD , Guo , SL and Cai , GX . 2003 . Effect of plastic-film mulch on water and nitrogen use by spring maize and on fate of applied nitrogen in the southern Loess Plateau . Chinese Journal of Applied Ecology , 14 ( 11 ) : 1901 – 1905 .
- Evans LT 1998 Greater crop production In: Waterlow JC Armstrong DG Fowdenand L Riley R Feeding a world population of more than eight billion people – A challenge to science Cary, NC, , USA Oxford University Press 89 97
- Hikosaka , K , Hanba , YT , Hirose , T and Terashima , I . 1998 . Photosynthetic nitrogen-use efficiency in woody and herbaceous plants . Functional Ecology , 12 ( 6 ) : 896 – 905 .
- Hikosaka , K . 2004 . Interspecific difference in the photosynthesis–nitrogen relationship: patterns, physiological causes, and ecological importance . Journal of Plant Research , 117 ( 6 ) : 481 – 494 .
- Hu , YC , Shao , HB , Chu , LY and Wu , G . 2006 . Relationship between water use efficiency (WUE) and production of different wheat genotypes at soil water deficit . Colloids and Surfaces B: Biointerfaces , 53 ( 2 ) : 271 – 277 .
- Jamieson , PD , Martin , RJ and Francis , GS . 1995 . Drought influences on grain yield of barley, wheat, and maize . New Zealand Journal of Crop and Horticultural Science , 23 : 55 – 66 .
- Kang , SZ , Shi , WJ , Hu , XT and Liang , YL . 1998 . Effects of regulated deficit irrigation on physiological indices and water use efficiency of maize . Transactions of the Chinese Society of Agricultural Engineering , 14 ( 4 ) : 82 – 87 .
- Liu , GS , Guo , AH , Ren , SX , An , SQ , Yang , YY , Mao , F and Zhao , HR . 2004 . Effects of different mulching methods on diurnal variation of leaf photosynthesis and water use efficiency for summer maize . Journal of Soil and Water Conservation , 18 ( 2 ) : 126 – 131 .
- Liu , JH , Xu , XX , Yang , G , Mu , XM and Wang , W . 2003 . Study on biomass of secondary shrubbery community in small watershed of Loess Hill and Gully Region . Acta Botanica Boreali-Occidentalia Sinica , 23 ( 8 ) : 1362 – 1366 .
- Liu , WZ and Zhang , XC . 2007 . Optimizing water and fertilizer input using an elasticity index: A case study with maize in the Loess Plateau of China . Field Crops Research , 100 ( 2–3 ) : 302 – 310 .
- Long , SP , Zhu , XG , Naidu , SL , Ort , DR and Usda , ARS . 2006 . Can improvement in photosynthesis increase crop yields? . Plant. Cell and Environment , 29 : 315 – 330 .
- Murchie , EH , Chen , Y , Hubbart , S , Peng , S and Horton , P . 1999 . Interactions between senescence and leaf orientation determine in situ patterns of photosynthesis and photoinhibition in field-grown rice . Plant Physiology , 119 ( 2 ) : 553 – 564 .
- Page , AL , Miller , RH and Keeney , DR . 1982 . Methods of soil analysis. Part 2. Chemical and microbiological properties , 2nd edn , Madison : American Society of Agronomy Press .
- Penuelas , J , Filella , I , Llusia , J , Siscart , D and Pinol , J . 1998 . Comparative field study of spring and summer leaf gas exchange and photobiology of the Mediterranean trees Quercus ilex and Phyllyrea latifolia . Journal of Experimental Botany , 49 ( 319 ) : 229 – 238 .
- Poorter , H and Evans , JR . 1998 . Photosynthetic nitrogen-use efficiency of species that differ inherently in specific leaf area . Oecologia , 116 ( 1 ) : 26 – 37 .
- Reich , PB , Hobbie , SE , Lee , T , Ellsworth , DS , West , JB , Tilman , D , Knops , JMH , Naeem , S and Trost , J . 2006 . Nitrogen limitation constrains sustainability of ecosystem response to CO2 . Nature , 440 ( 7086 ) : 922 – 925 .
- Sadras , VO . 2004 . Yield and water-use efficiency of water- and nitrogen-stressed wheat crops increase with degree of co-limitation . European Journal of Agronomy , 21 ( 4 ) : 455 – 464 .
- Shangguan , ZP , Shao , MA and Dyckmans , J . 2000 . Nitrogen nutrition and water stress effects on leaf photosynthetic gas exchange and water use efficiency in winter wheat . Environmental and Experimental Botany , 44 ( 2 ) : 141 – 149 .
- Shangguan , ZP , Shao , MA , Ren , SJ , Zhang , LM and Xue , Q . 2004 . Effect of nitrogen on root and shoot relations and gas exchange in winter wheat . Botanical Bulletin of Academia Sinica , 45 ( 1 ) : 49 – 54 .
- Shao , HB , Chu , LY , Shao , MA and Zhao , CX . 2008 . Advances in functional regulation mechanisms of plant aquaporins: Their diversity, gene expression, localization, structure and roles in plant soil-water relations (Review) . Molecular Membrane Biology , 25 ( 3 ) : 179 – 197 .
- Shi , CL , Guo , JX , Yan , CR , Li , YZ and Mei , XR . 2007 . Diurnal variation of instantaneous gas-exchange over canopy and leaf level of spring corn in arid areas . Transactions of the Chinese Society of Agricultural Engineering , 23 ( 1 ) : 24 – 31 .
- Takashima , T , Hikosaka , K and Hirose , T . 2004 . Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduous Quercus species . Plant Cell and Environment , 27 ( 8 ) : 1047 – 1054 .
- Warren , CR and Adams , MA . 2004 . Evergreen trees do not maximize instantaneous photosynthesis . Trends in Plant Science , 9 ( 6 ) : 270 – 274 .
- Watling , JR , Press , MC and Quick , WP . 2000 . Elevated CO2 induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum . Plant Physiology , 123 ( 3 ) : 1143 – 1152 .
- Yan , CR , Mei , XR , Ju , H and Chi , J . 2006 . Effects of fertilization on the growth, development and water use efficiency of spring millet . Chinese Journal of Eco-Agriculture , 14 ( 1 ) : 142 – 144 .
- Zhang , X . 2007 . Drought adaptability of main minor crops in Loess Plateau . Journal of Arid Land Resources and Environment , 21 ( 8 ) : 111 – 115 .
- Zheng , SX and Shangguan , ZP . 2007a . Spatial patterns of photosynthetic characteristics and leaf physical traits of plants in the Loess Plateau of China . Plant Ecology , 191 ( 2 ) : 279 – 293 .
- Zheng , SX and Shangguan , ZP . 2007b . Photosynthetic physiological adaptabilities of Pinus tabulaeformis and Robinia pseudoacacia in the Loess Plateau . Chinese Journal of Applied Ecology , 18 ( 1 ) : 16 – 22 .