Abstract
Yellow lupin (Lupinus luteus L.) is a high-protein grain legume growing well in a range of acid soil types. However, the lack of herbicide tolerance for effective weed control has limited its adoption in broad acre farming systems. In order to breed herbicide-tolerant cultivars, a source of resistance needs to be identified. This paper reports the identification of a number of highly tolerant yellow lupin genotypes to metribuzin and carfentrazone-ethyl herbicides. Suitable dosages (LD50) of the two herbicides were identified as 150 g ai ha−1 (ai = active ingredient) for metribuzin and 200 g ai ha−1 for carfentrazone-ethyl. These dosages were used to screen diverse yellow lupin landraces and wild types to quantify the diversity of the germplasm. Considerable variation in response to the two herbicides was observed among the genotypes screened, ranging from complete plant death to no symptoms. Yellow lupin cv. Pootallong was susceptible to metribuzin, but three accessions (P20873, P23291 and P28800) were identified as tolerant with no plant damage. Four accessions (P20873, P20879, P23298 and P28832) showed higher tolerance to carfentrazone-ethyl than the standard cultivar Pootallong. A breeding line P20873 had the highest tolerance to both herbicides, indicating the possibility for developing yellow lupin cultivars with tolerance to both herbicides.
Introduction
Yellow lupin (Lupinus luteus L.) is a high-protein grain legume growing well in a range of hostile soil types. Its ability to tolerate higher aluminium toxicity in soils (French et al. Citation2001), lower phosphorus availability in the phosphate-fixing soils (Bolland et al. Citation2000, Citation2001) and transient water logging conditions (Davies et al. Citation2000) make it more suitable to a wider range of environments where other lupins struggle to grow. Yellow lupin grain has a high protein content (>38%) and twice the cysteine and methionine contents of most other lupins and pulses (Gladstones et al. Citation1998). The higher protein of yellow lupin offers significant advantages to intensive livestock industries as well as human food (Glencross et al. Citation2004).
Weed control is one of the major factors limiting yellow lupin production. The lack of herbicide options leads to poor control of broad-leaved weeds within the crop. Lupin plants are poor competitors against broad-leaved weeds in the field. Wild radish (Raphanus raphanistrum L.)—the predominant weed in lupin growing areas (Cheam & Code Citation1995)—causes up to 66% seed yield loss in lupin crops when densities exceed 28 wild radish plants m−2 (Hashem et al. Citation2006). Herbicide application has become the dominant method of weed control in the minimum tillage farming system which conserves soil moisture and prevents soil erosion and is widely practised in dry land agriculture (Perry et al. Citation1998; Bowran & Hashem Citation2008). So far, no herbicide-tolerant cultivars in yellow lupin have been developed.
Metribuzin and carfentrazone-ethyl are used for post-emergent application to control major broad-leaved weeds including wild radish, capeweed (Arctotheca calendula (L.) Levyns) and doublegee (Emex australis Steinh.). The two herbicides have different modes of action. Metribuzin—a triazinone herbicide inhibiting photosystem II—is the main post-emergent herbicide used in combination with diflufenican. Carfentrazone-ethyl—a member of phenyl triazolinone herbicides which inhibits protoporphyrinogen oxidase (Dayan et al. Citation1997)—is a relatively new herbicide in dry land agriculture. Carfentrazone-ethyl in a mix with 2-methyl-4-chlorophenoxyacetic acid (MCPA) is capable of controlling a wide range of broad-leaved weeds (Cumming Citation2000). Development of yellow lupin cultivars tolerant to either or both of these two herbicides would provide an effective tool for weed management.
Herbicide-tolerant cultivars have been bred for several major crops (Oxtoby & Hughes Citation1989) and identification of sources of tolerance is the first step in developing herbicide-tolerant cultivars. Differential tolerance to metribuzin between cultivars has been reported in soybean (Barrentine et al. Citation1982), barley (Caldwell & O'Sullivan Citation1985), field pea (Al-khatib et al. Citation1997), tomato (Stephenson et al. Citation1976), winter wheat (Schroeder et al. Citation1985) and narrow-leafed lupin (Si et al. Citation2006). Selecting tolerant genotypes has improved metribuzin tolerance in cultivars of soybean, wheat and narrow-leafed lupin (Barrentine et al. Citation1982; Schroeder et al. Citation1985; Si et al. Citation2006). Selectivity to carfentrazone-ethyl is also found between soybean and broad-leaved weeds (Thompson & Nissen Citation2000). This paper reports the genotypic variation and identification of sources of tolerance to metribuzin and carfentrazone-ethyl in yellow lupin germplasm.
Materials and methods
Materials
Yellow lupin cv. Pootallong and narrow-leafed lupin cvs Mandelup and Danja were used in dose response experiments to assess their tolerance to metribuzin and carfentrazone-ethyl. Mandelup is tolerant to metribuzin and Danja is susceptible. A biotype of broad-leaved weed wild radish WARR 25 was included in the carfentrazone-ethyl dose response experiment to determine the selectivity between lupins and the weed. This biotype was collected from the Western Australian lupin growing area.
Fifty yellow lupin germplasm accessions including cv. Pootallong were selected from the Australian yellow lupin germplasm collection at the Department of Agriculture and Food, Western Australia to determine the magnitude of genotypic variation in herbicide tolerance. These accessions of wild types, landraces, breeding lines and cultivars came from various parts of the world (), representing the diversity of yellow lupin germplasm.
Table 1 Survival (%) and plant damage score of 50 yellow lupin accessions in response to 150 g ai ha−1 metribuzin and 200 g ai ha−1 carfentrazone-ethyl.
Dose response experiments
Dose responses with metribuzin were assessed on cvs Pootallong, Mandelup and Danja, and dose responses with carfentrazone-ethyl were assessed on cvs Pootallong, Mandelup and wild radish. Seeds of each genotype were sown into 200 mm diameter pots containing the School of Plant Biology standard potting mix (2.5 parts composted pinebark, 1 part coco peat, 1.5 parts riversand, plus additional macro- and micro-nutrients). Pots were arranged in a completely randomized design with four replicates per herbicide treatment, each containing 10 plants. The pots were maintained in a glasshouse at 20 °C during day time and 12 °C at night for germination and growth. Seedlings were watered overhead at 2–3 day intervals. There were 14 treatments, which were applied when the seedlings were at the four-leaf stage. Plants were sprayed with one of seven doses of metribuzin or carfentrazone-ethyl (0, 50, 100, 200, 400, 800, 1600 g ai ha−1; ai = active ingredient) in a spray cabinet fitted with two flat fan nozzles. The herbicides were delivered in 112 L ha−1 water with two passes at 200 kPa. After herbicide application, plants were returned to the glasshouse immediately and watering resumed 48 hours afterwards. Measurements of herbicide damage were recorded 2 weeks after herbicide application. The purpose of this experiment was to determine suitable dosages of the two herbicides for germplasm screening.
Germplasm evaluation experiments
Seeds of 50 yellow lupin germplasm accessions and the tolerant narrow-leafed lupin cv. Mandelup, as a control, were sown into 200 mm pots containing standard potting mix as mentioned earlier. There were eight plants per pot and four pots (replicates) for each herbicide treatment. The pots were arranged in a completely randomized design and maintained in a glasshouse at 20 °C during day time and 12 °C at night for germination and growth. Seedlings were watered overhead at 2–3 day intervals. When the seedlings were at the four-leaf stage, they were sprayed either metribuzin or carfentrazone-ethyl with doses determined from the previous experiment using the same spraying cabinet as described for the dose response experiments. Plants were returned to the glasshouse immediately after herbicide application and watering resumed 48 hours afterwards. Herbicide damage was assessed on individual plants.
Measurements
Survival rate, shoot dry weight and leaf damage scores were measured in both experiments 2 weeks after herbicide application. Survival rate for each replicate was expressed as a percentage of live plants relative to plant numbers before treatment. Shoots were cut at soil level, dried in a fan-forced oven at 80 °C for 48 h, and weighed. Shoot dry weights (% of control) were expressed as percentages of each treatment in relation to the untreated control. Individual plants were visually rated for damage using the European Weed Research Council (EWRC) ordinal scale from 1 to 9 (Dear et al. Citation2003), where 1 represented no damage; 2, very slight damage with some stunting and yellowing just visible; 3, slight damage with stunting and yellowing obvious; 4, substantial chlorosis and/or stunting; 5, strong chlorosis/stunting and thinning of stand; 6, increasing severity; 7, increasing severity; 8, increasing severity; and 9, plant dead.
Statistical analysis
Regression analysis was undertaken for dose response curves using SigmaPlot version 10.1 (Systat Software Inc, Point Richmond, CA, US). The LD50 (rate for killing 50% plants) and GR50 (rate for 50% growth reduction) values were estimated using the four-parameter logistical curve regression (Seefeldt et al. Citation1995). These values were used to calculate LD50 and GR50 ratios, respectively. Analysis of variance using GenStat 9.2 software was conducted for each herbicide in the germplasm evaluation experiment.
Results
Variation in metribuzin tolerance
Susceptibility of yellow lupin cultivar to metribuzin
Yellow lupin cv. Pootallong was very susceptible to metribuzin. Pootallong had even less seedling survival than the susceptible narrow-leafed lupin cv. Danja at 200 g ai ha−1 metribuzin (A). Pootallong had the lowest shoot dry weight (B) and the highest damage scores (C) across all herbicide rates. The LD50 value for cv. Pootallong was 130 g ai ha−1 metribuzin, less than that of cv. Danja, and the LD50 value for the tolerant cultivar Mandelup was 1280 g ai ha−1 metribuzin (). The rate of 150 g ai ha−1 metribuzin, therefore, was chosen to screen the germplasm for tolerance.
Figure 1 Dose response of yellow lupin cultivar ‘Pootallong’ (▵) and narrow-leafed lupin cultivars ‘Mandelup’ (♦) and ‘Danja’ (▪) to metribuzin with plants grown in glasshouses. A, Survival (%). B, Shoot dry weight (percentage of control) excluding dead plants. C, Plant damage score, where 1 represents no symptoms and 9 represents dead plants. The vertical bars above and below the graphed points are standard errors.
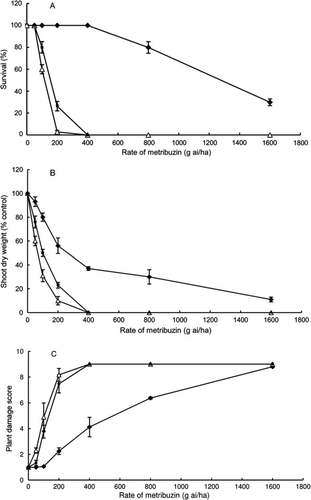
Table 2 LD50 and GR50 values (mean ±SE) of metribuzin for yellow lupin and narrow-leafed lupin cultivars.
Variation among germplasm accessions
The range of plant survival at 150 g ai ha−1 metribuzin among the 50 yellow lupin accessions varied from 15.6% to 100% (). All accessions had a higher survival rate than cv. Pootallong except for line P28900 (Poland) which had 15.6% survival. About half the accessions had 100% survival; the remainder ranged from 15.6% to 90%. Plant damage scores varied from a score of 1 (no damage) to 8.53 (most plants killed). The lowest surviving accession P28900 had the highest damage score of 8.53 and Pootallong had a damage score of 8.13. There were 43 accessions with damage scores significantly (P < 0.05) lower than Pootallong, indicating better tolerance. There were three accessions with no symptoms of damage to metribuzin with scores of 1 for all 32 plants examined. These three accessions (P20873, P23291 and P28800) had the same score as the tolerant narrow-leafed lupin cv. Mandelup.
Genotypic responses estimated from the plant damage score were classified into five categories from very tolerant to very susceptible (). The majority of the germplasm accessions (68%) were moderate to tolerant. The very tolerant genotypes accounted for 10% of accessions and 6% were very susceptible. Pootallong and two other accessions were very susceptible, having the highest damage scores with plants close to mortality. The very tolerant category included three accessions P20873, P23291 and P28800 with no damage symptoms and two accessions P20875 and P28329 with minimal damage.
Table 3 Tolerance categories of yellow lupin germplasm in response to metribuzin at 150 g ai ha−1.
Relationship between the origin and breeding status of the germplasm and metribuzin tolerance
There was no clear pattern between tolerance and country of origin in the germplasm. Both the most susceptible and the most tolerant accessions were from Poland. For the four Australian cultivars and breeding lines tested, metribuzin damage scores varied from 2.6 to 8.1. Tolerance was observed in accessions from Germany, Poland, Byelorussia and Ukraine. The highest tolerance was observed in cultivars or breeding lines () originating from diverse sources. Wild accessions and landraces were variable from moderate tolerance to very susceptible, but none had a high level of tolerance.
Table 4 Mean and range of plant damage score1 in the different categories of breeding status of 50 yellow lupin germplasm subjected to 150 g ai ha−1 metribuzin and 200 g ai ha−1 carfentrazone-ethyl.
Variation in carfentrazone-ethyl tolerance
Selectivity between wild radish and lupins
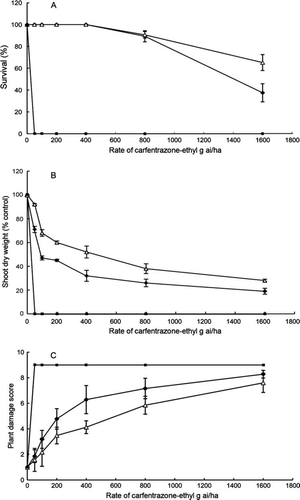
At rates ≥50 g ai ha−1 of carfentrazone-ethyl, all wild radish plants were killed (A), indicating its high efficacy on wild radish. Seedlings of cvs Mandelup and Pootallong had 100% survival at 400 g ai ha−1, although survival started to decrease when rates were ≥800 g ai ha−1. Mandelup seedlings had less shoot dry weight than Pootallong (B) and plant damage scores for Mandelup were higher than Pootalong at rates ≥200 g ai ha−1 carfentrazone-ethyl (C), indicating that Pootallong was more tolerant to carfentrazone-ethyl than Mandelup. The LD50 rate of 200 g ai ha−1 carfentrazone-ethyl—four times the rate to control wild radish—was chosen to screen the germplasm for tolerance.
Fig. 2 Dose response of yellow lupin cultivar Pootallong (▵) and narrow-leafed lupin cultivar Mandelup (♦) and wild radish (▪) to carfentrazone-ethyl with plants grown in glasshouses. A, Survival (%). B, Shoot dry weight (percentage of control) excluding dead plants. C, Plant damage score, where 1 represents no symptoms and 9 represents dead plants. The vertical bars above and below the graphed points are standard errors.
Variation among accessions
At 200 g ai ha−1 carfentrazone-ethyl, all yellow lupin accessions had 100% survival rate and plant damage scores varied between 3.4 and 6.7. Pootallong had a damage score of 4.8, significantly lower than that of Mandelup (score = 6.5), confirming the higher tolerance to carfentrazone-ethyl. Four accessions (P20873, P20879, P23298 and P28832) had significantly lower damage scores than Pootallong, indicating a higher tolerance to carfentrazone-ethyl than Pootallong. Interestingly, P20873 was the most tolerant accession to both metribuzin and carfentrazone-ethyl.
Discussion
This paper reports, for the first time, sources of herbicide tolerance in yellow lupin. Genotypic variability was large among the yellow lupin germplasm accessions tested for both metribuzin and carfentrazone-ethyl tolerance, with more variability for metribuzin tolerance than for carfentrazone-ethyl. Metribuzin tolerance in yellow lupin varied from very susceptible with almost all plants killed to very tolerant with all plants showing no sign of damage. This range of variation is supported by differential tolerance to metribuzin in various crops including soybean (Barrentine et al. Citation1982), field pea (Al-khatib et al. Citation1997), barley (Caldwell & O'Sullivan Citation1985) and wheat (Schroeder et al. Citation1985). The yellow lupin cultivar Pootallong is very susceptible; hence metribuzin has not been used for weed control in yellow lupin. However, most germplasm accessions had better tolerance than Pootallong. Three accessions (P20873, P23291 and P28800) were identified with no plant damage at 150 g ai ha−1 metribuzin, the same level of tolerance as the tolerant narrow-leafed lupin cv. Mandelup. Identification of tolerant sources makes it possible to develop herbicide-tolerant cultivars.
Breeding for metribuzin tolerance in yellow lupin may be relatively simple as the tolerance source is either an elite breeding line or cultivar from Europe. The conventional crossing method would be sufficient to transfer tolerance into elite breeding materials adapted in dry-land agricultural conditions. Metribuzin-tolerant lupin, soybean and wheat have already been developed from genotypic variations (Barrentine et al. Citation1982; Si et al. Citation2006; Kleemann & Gill Citation2007). Progress for developing metribuzin tolerant cultivars would be rapid as single plants of tolerant progenies in segregating populations generated from crosses could be selected visually by plant damage scores as demonstrated in this research. Further increases in herbicide tolerance than the three very tolerant genotypes identified could be possible by pyramiding genes from the various tolerant genotypes assuming that they possess tolerance genes at different loci. A further increase in metribuzin tolerance in narrow-leafed lupin was achieved by crossing two tolerant mutants, as tolerance in each mutant is controlled by a single gene and the effects of these two genes were additive (Si et al. Citation2010). Inheritance of metribuzin tolerance in yellow lupin is unknown and may be recessive or quantitatively controlled. We therefore suggest that inheritance studies of the herbicide tolerance trait and complementation analysis between the various sources of tolerance are performed before an appropriate course of breeding is determined.
Herbicide-tolerant cultivars are the most effective weed management strategy under the minimum tillage farming systems which prevent soil erosion and conserve soil moisture. Metribuzin is the main post-emergent herbicide used in narrow-leafed lupin to control the major broad-leaved weeds (Bowran & Hashem Citation2008). Pre-emergent application of metribuzin can also control brome grass (Kleemann & Gill Citation2007), hence the motivation for developing metribuzin tolerant wheat cultivars. The registered rate of metribuzin in narrow-leafed lupin is 75–112.5 g ai ha−1 in a mix with diflufenican. The narrow-leafed lupin cv. Mandelup is tolerant to this rate in the field (Dhammu & Nicholson Citation2005). We expect that the three tolerant accessions of yellow lupin identified at 150 g ai ha−1 metribuzin in this research would tolerate the registered rate in the field. Development of metribuzin-tolerant cultivars in yellow lupin would provide an effective option for weed control, encouraging farmers to grow yellow lupin in a wide range of soil types and hence improve productivity in hostile soil conditions. Although glyphosate-tolerant major crops such as soybean and canola developed through genetic modification are already grown widely, tolerance to other herbicides will complement glyphosate tolerance because usage of multiple herbicides in rotation will reduce the risk of weeds developing resistance (Duke Citation2005).
Tolerance to carfentrazone-ethyl would provide a useful additional herbicide option in yellow lupin as this herbicide has a different mode of action capable of controlling a wide range of broad-leaved weeds in dry-land agriculture (Cumming Citation2000). The large selectivity between yellow lupin cv. Pootalloong and wild radish by the dose responses is supported by the selectivity between soybean and morning glory (Dayan et al. Citation1997). Identification of four accessions with higher tolerance than cv. Pootallong is encouraging. Although these four accessions had some foliage damage at 200 g ai ha−1 carfentrazone-ethyl, they may tolerate the herbicide at 50 g ai ha−1, a rate registered for use in the mix to control weeds. Higher rates used in the germplasm screening were to ensure sufficient herbicide pressure to separate tolerant plants. Accession P20873, a breeding line from Ukraine, has the highest tolerance to both metribuzin and carfentrazone-ethyl and is the ideal source to develop cultivars tolerant to both herbicides. Stacking herbicide tolerance in one cultivar would provide more herbicide options for different weeds.
The identified genotypes responding differently to the application of the two herbicides can be useful material for the study of physiological and genetic mechanism of herbicide tolerance. Differences in tolerance among germplasm accessions are likely due to the different rates of metribuzin being metabolized by plants; tolerant genotypes of soybean have higher rates of metribuzin metabolism (Falb & Smith Citation1984). Carfentrazone-ethyl has a different mode of action from metribuzin (Dayan et al. Citation1997) and, as expected, the germplasm accessions responded differently to carfentrazone-ethyl. Although LD50 has been indicated as the best dosage for screening the germplasm, application of several different dosages to further characterize the identified tolerant genotypes will help us to understand better the tolerance capacity of these genotypes.
Conclusions
Large genotypic variation exists in yellow lupin germplasm for herbicide tolerance. Yellow lupin cv. Pootallong is very susceptible to metribuzin. Three elite breeding lines from Europe are very tolerant and they can be used as sources to develop metribuzin-tolerant cultivars. The considerable selectivity between yellow lupin and wild radish in response to carfentrazone-ethyl suggests the potential use of this herbicide in yellow lupin. One accession, P20873, a breeding line, is tolerant to both herbicides. Therefore, it is possible to develop yellow lupin cultivars tolerant to both herbicides for the effective control of a wide range of weeds.
Acknowledgements
The authors thank Dr Michael Walsh for providing wild radish seed, the glasshouse staff for their support during the experiment and the Western Australian Herbicide Resistance Initiative for allowing us to use their dedicated herbicide sprayer. We also thank Dr Xuanli Ma for her help with producing high-quality figures.
References
- Al-khatib , K , Libbey , C , Kadir , S and Boydston , R . 1997 . Differential varietal response of green pea (Pisum sativum) to metribuzin . Weed Technology , 11 : 775 – 781 .
- Barrentine WL , Hartwig EE , Edwards CJ Jr , Kilen TC 1982 . Tolerance of 3 soybean (Glycine max) cultivars to metribuzin . Weed Science 30 : 344 – 348 .
- Bolland , MDA , Sweetingham , MW and Jarvis , RJ . 2000 . Effect of applied phosphorus on the growth of Lupinus luteus, L. angustifolius and L. albus in acidic soils in the south-west of Western Australia . Australian Journal of Experimental Agriculture , 40 : 79 – 92 .
- Bolland , MDA , Sweetingham , MW and Jarvis , RJ . 2001 . Effect of Pleiochaeta setosa on field responses of Lupinus angustifolius and L. luteus to applications of phosphorus . Australian Journal of Experimental Agriculture , 41 : 549 – 556 .
- Bowran D , Hashem A 2008 . The role of weed management in sustaining systems for lupin production . Palta JA , Berger JD . Proceedings of the 12th International Lupin Conference , Fremantle , , Australia , 14–18 September 2008 . Pp. 11 – 14 .
- Caldwell , CD and O'Sullivan , PA . 1985 . Differential tolerance of 2 barley cultivars to metribuzin . Canadian Journal of Plant Science , 65 : 415 – 421 .
- Cheam , AH and Code , GR . 1995 . The biology of Australian weeds 24. Raphanus raphanistrum L . Plant Protection Quarterly , 10 : 1 – 13 .
- Cumming G 2000 . AFFINITY 400DF. A new herbicide with a new mode of action (Group G) for broadleaf weed control in cereals . In: Stewart V . 2000 Weeds Updates—Western Australia . South Perth , Western Australian Department of Agriculture and Food . Pp. 91 – 94 .
- Davies , CL , Turner , DW and Dracup , M . 2000 . Yellow lupin (Lupinus luteus) tolerates waterlogging better than narrow-leafed lupin (L. angustifolius). III. Comparison under field conditions . Australian Journal of Agricultural Research , 51 : 721 – 727 .
- Dayan , FE , Duke , SO , Weete , JD and Hancock , HG . 1997 . Selectivity and mode of action of carfentrazone-ethyl, a novel phenyl triazolinone herbicide . Pesticide Science , 51 : 65 – 73 .
- Dear , BS , Sandral , GA , Spencer , D , Khan , MRI and Higgins , TJV . 2003 . The tolerance of three transgenic subterranean clover (Trifolium subterraneum L.) lines with the bxn gene to herbicides containing bromoxynil . Australian Journal and Agricultural Research , 54 : 203 – 210 .
- Dhammu HS , Nicholson D 2005 . Herbicide tolerance of lupin varieties . In: Douglas A . 2005 Weeds Update —Western Australia . South Perth , Western Australian Department of Agriculture and Food . Pp. 77 – 79 .
- Duke , SO . 2005 . Taking stock of herbicide-resistant crops ten years after introduction . Pest Management Science , 61 : 211 – 218 .
- Falb , LN and Smith , AE . 1984 . Metribuzin metabolism in soybeans. Characterization of the intraspecific differential tolerance . Journal of Agricultural Food and Chemistry , 32 : 1425 – 1428 .
- French , RJ , Sweetingham , MW and Shea , GG . 2001 . A comparison of the adaptation of yellow lupin (Lupinus luteus) and narrow-leafed lupin (L. angustifolius) to acid sand plain soils in low rainfall agricultural areas of Western Australia . Australian Journal of Agricultural Research , 52 : 945 – 954 .
- Gladstones JS , Atkins C , Hamblin J 1998 . Lupins as crop plants, biology, production and utilization . Wallingford , , UK , CAB International .
- Glencross , BD , Evans , D , Jones , JB and Hawkins , WE . 2004 . Evaluation of the dietary inclusion of yellow lupin (Lupinus luteus) kernel meal on the growth, feed utilization and tissue histology of rainbow trout (Oncorhynchus mykiss) . Aquaculture , 235 : 411 – 422 .
- Hashem A , Pathan S , French R 2006 . Wild radish-lupin competition: difference in the competitive ability of lupin cultivars . Preston C , Watts JH , Crossman ND . Proceedings of the 15th Australian Weeds Conference , Adelaide , Australia 24–28 September 2006 . Pp. 391 – 394 .
- Kleemann , SGL and Gill , GS . 2007 . Differential tolerance in wheat (Triticum aestivum L.) genotypes to metribuzin . Australian Journal of Agricultural Research , 58 : 452 – 456 .
- Oxtoby , E and Hughes , MA . 1989 . Breeding for herbicide resistance using molecular and cellular techniques . Euphytica , 40 : 173 – 180 .
- Perry MW , Dracup M , Nelson P , Jarvis R , Rowland I , French RJ 1998 . Agronomy and farming systems . In: Gladstones JS , Atkins C , Hamblin J . Lupins as crop plants, biology, production and utilization . Wallingford , , UK , CAB International . Pp. 291 – 338 .
- Schroeder J , Banks PA , Nicholas RL 1985 . Soft red winter wheat (Triticum aestivum) cultivar response to metribuzin . Weed Science 34 : 66 – 69 .
- Seefeldt , SS , Jensen , JE and Fuerst , EP . 1995 . Log-logistic analysis of herbicide dose response relationships . Weed Technology , 9 : 218 – 225 .
- Si , P , Sweetingham , MW , Buirchell , BJ , Bowran , DG and Piper , T . 2006 . Genotypic variation in metribuzin tolerance in narrow-leafed lupin (Lupinus angustifolius L.) . Australian Journal of Experimental Agriculture , 46 : 85 – 91 .
- Si P , Pan G , Sweetingham MW 2010 . Semi-dominant genes confer additive tolerance to metribuzin in narrow-leafed lupin (Lupinus angustifolius L.) mutants . Euphytica doi: DOI 10.1007/s10681-010-0278-9
- Stephenson , GR , McLeod , JE and Phatak , SC . 1976 . Differential tolerance of tomato cultivars to metribuzin . Weed Science , 24 : 161 – 165 .
- Thompson , WM and Nissen , SJ . 2000 . Absorption and fate of carfentrazone-ethyl in Zea mays, Glycine max and Abutilon theophrasti . Weed Science , 48 : 15 – 19 .