
Abstract
The intrinsic relationship between grape berry ripening and temperature is not well understood. The aim was to compare ripening in three cultivars—Vitis vinifera L. cv. Chardonnay, cv. Merlot and cv. Semillon—in response to temperature. In the post-veraison period (soluble solids 8–12 oBrix), whole vines were exposed to five controlled temperature regimes, with the day temperature ranging from 20–40 °C, and the berry attributes and the ripening processes followed. Temperature had little effect on the berry expansion process in all cultivars. However, there were marked effects of temperature on the accumulation of both dry matter and sugar in all varieties. The rate of ripening in all three cultivars was a curvilinear function of temperature. Rate of ripening of Chardonnay berries was highest at 25 °C and for Semillon and Merlot at 35 °C and 40 °C, respectively. Application of the temperature response to field-grown Semillon vines accurately predicted the seasonal progression of ripening.
Introduction
Grapevines (Vitis vinifera L.) are an economically important crop in Australia and elsewhere and there is a need to have a better understanding of the vegetative and reproductive growth and development processes in relation to the abiotic environment, particularly in relation to potential changes in climate (Jones et al. Citation2010). Greer and colleagues (Greer & Sicard Citation2009; Greer et al. Citation2011; Greer Citation2012; Greer & Weedon Citation2012) have undertaken an extensive study of the interactive effects of temperature and light on these growth and developmental processes in the grapevine cultivar, Semillon. This cultivar represents the second most important white grapevine within Australia (Greer & Weston Citation2010), but is especially important in some regional locations. In many locations within Australia, grapevines experience relatively high temperature conditions during summer (Gladstones Citation1992) as well as high incident radiation (Smart Citation1973; Smart & Sinclair Citation1976). Many studies have investigated the effects of high temperature/high light conditions on a range of cultivars, including Sultana (Sommer et al. Citation2000), Shiraz (Soar et al. Citation2009), Cardinal, Chardonnay and Chenin Blanc (Sepúlveda & Kliewer Citation1986b), Cabernet Sauvignon (Crippen & Morrison Citation1986; Smart et al. Citation1988) and Merlot (Spayd et al. Citation2002), to name a few.
Fruit ripening is one of the more important aspects of the reproductive process for the grower as it determines the time of harvest and has an influence on the berry composition, which can influence wine style and quality. High temperatures and/or high light intensities are known to have effects on the accumulation of soluble solids (Bergqvist et al. Citation2001). For example, Keller et al. (Citation1998) have shown post-veraison berry ripening of Cabernet Sauvignon vines declined markedly when the light exposure was decreased. A similar result occurred with post-veraison berries of Nebbiolo vines (Chorti et al. Citation2010). By contrast, Spayd et al. (Citation2002) compared berries from the eastern and western sides of the canopy of Merlot vines with some exposed and others shaded over the whole ripening period and showed berry mass and soluble solids content were largely unaffected by the treatments. In a comparison with the eastern and western (warmer) sides of the canopies of Merlot and Cabernet vines, Shellie (Citation2011) showed that berry growth across the whole growing season was higher in the east-facing berries which were reported to be cooler than west-facing berries, but actual berry temperatures were not given. There are several studies indicating that high temperatures can delay ripening and limit sugar accumulation in the late-ripening stage (Matsui et al. Citation1986; Sepúlveda & Kliewer Citation1986a). Many of these studies have been carried out in controlled environments.
Specific effects of temperature on the berry ripening process have also been well addressed. For example, Radler (Citation1965) compared the ripening of Sultana grapes at two temperature regimes imposed from bloom to harvest and noted that elevated temperatures (33 °C) caused a reduction in berry size but a more rapid accumulation of sugars. By contrast, Buttrose et al. (Citation1971) compared the effect of temperature on Cabernet Sauvignon vines over the whole season and found no effect on berry growth or on the rate of sugar accumulation. Yet again, Kliewer (Citation1977) observed with Cabernet Sauvignon and Tokay vines that berry size and soluble solids per berry declined progressively over the whole berry development stage as temperature increased from 25 to 40 °C, when exposed before veraison. There was also no effect on berry weight or on solid soluble solids accumulation when the table grape cultivar, Aki Queen, was grown at temperatures of 20 and 30 °C at different stages of berry development (Yamane et al. Citation2006). These results are, therefore, inconsistent across different cultivars but also the temperature range varies considerably and raises the question of whether the grape berry ripening process is a temperature-dependent process. However, it was noted by Greer & Weedon (Citation2012) with Semillon vines grown in exposed and shaded conditions, that the rate of berry ripening process increased sharply during a period of high temperature, this implied there was a high temperature response for berry ripening of this cultivar.
Although there have been a number of comparative studies of several grapevine cultivars, these have mostly focused on gas exchange and water relations. For example, Sepúlveda & Kliewer (Citation1986b) compared the stomatal behaviour of three cultivars when exposed to high temperatures. Similarly, Moutinho-Pereira et al. (Citation2007) compared the gas exchange attributes of three more cultivars grown in a Mediterranean climate, Costa et al. (Citation2012) compared the stomatal response to water deficit in five grapevine cultivars and Rogiers et al. (Citation2009) compared gas exchanges and leaf water potential in 10 common grapevine cultivars. However, Sadras et al. (Citation2008) compared the dynamics of soluble solids accumulation in 12 cultivars. In contrast, there is a paucity of work comparing the response of the berry developmental processes to temperature in different cultivars.
Accordingly, the objective of the present study was to compare the sugar accumulation process of three common grapevine cultivars, namely Vitis vinifera cv. Merlot, V. vinifera cv. Chardonnay and V. vinifera cv. Semillon and to assess the response of the berry ripening processes in each cultivar to temperature. The vines were grown under the same conditions in controlled environments to ensure the responses of the different cultivars could be readily compared. These cultivars were chosen primarily on the basis of availability as potted vines for the study. An additional objective was to model the ripening process on field-grown Semillon vines on the basis of the temperature-dependent response of ripening.
Materials and methods
Plant material and growth conditions
Fifteen potted Vitis vinifera cv. Chardonnay vines, 15 V. vinifera cv. Merlot vines and 10 V. vinifera cv. Semillon vines were placed in a cool room at 6 °C in late January (mid summer) for 2 months to ensure the vines became fully senesced and dormant (Weaver & Iwasaki Citation1977). All vines were 5 years old, of a similar size and grown in 25 L pots in a sand, soil and fertiliser potting medium. The vines were all pruned to two bud spurs and five current season's shoots were retained on each vine and each supported two bunches. The Chardonnay vines were kept in the cool room for an additional 8 days to enable matching of phenology with the other varieties. The shoots were grown as horizontally as possible tying them to bamboo supports. In late March, all the vines were transferred to and distributed among four controlled environment chambers (TH6000, Thermoline Scientific, Smithfield, NSW, Australia). In each chamber, the temperature was initially set at 15/10 °C (day/night) with a 12 h photoperiod and lighting supplied by 8 × 20 W compact fluorescent lamps (Cool White, Fairway, China) to allow for acclimation to commence. Three days later, the temperature was raised to 28/18 ± 0.5 °C, the atmospheric water vapour pressures were set and controlled at 1.5/1.2 ± 0.2 kPa and the photon flux density was set and maintained at 750 ± 10 µmol m−2 s−1 at canopy height (ca. 1.2 m above the floor), supplied by 4 × 1 kW high pressure discharge lamps (HQI-T1000, Osram, Germany) over a 12 h photoperiod. The vines were grown in these conditions for 140 days in total. An automatic watering system was used to supply water daily to ensure the pots were well watered. During the treatment periods, the pots were hand watered to run-off daily to ensure unlimited water, especially at high temperatures.
Treatment conditions
At approximately 86 days after budbreak (DAB), the whole vines of each cultivar were randomly allocated to five controlled environment chambers to ensure equal numbers of bunches were in each chamber for each cultivar but the growth conditions remained as above. After 10 days (96 to 107 DAB), the day/night temperatures were altered to 20/14, 25/17.5, 30/21, 35/24.5 and 40/28 ± 0.5 °C but all other growth conditions as above were maintained. These temperature regimes were based on vineyard canopy measurements across the growing season (Greer et al. Citation2010). These treatments were imposed, on the assumption that the high temperatures may be detrimental to the bunches and vines, thus for 6 days at 40 °C, 12 days at 35 °C and 18 days at 30–20 °C. The soluble solids concentration averaged 8.9 ± 0.9 oBrix for Merlot berries, 12.0 ± 0.1.4 oBrix for Chardonnay berries and 8.1 ± 1.6 oBrix for Semillon berries at the time the treatments were imposed, which was approximately at the start of the rapid increase in soluble solids concentration, i.e. veraison (Greer & Weston Citation2010; Greer & Weedon Citation2012). All bunches had between 150 and 200 berries. The berries all reached a soluble solids concentration of 18–20 oBrix at the completion of the treatments.
Vine measurements
Phenology
Using the modified E-L scheme (Coombe Citation1995), the phenology of each cultivar was recorded at weekly intervals to assess budbreak and time of flowering.
Berry attributes
Soluble solids concentration
The concentration of soluble solids was measured at the start of each treatment by detaching three berries per bunch, from the top, middle and bottom segments, from four to five bunches on each cultivar in each temperature treatment (12–15 in total on each occasion) and measuring the soluble solids concentration with a refractometer (PR-101, Atago, Tokyo, Japan). Thereafter, soluble solids concentration was measured at 1–3 day intervals by the same procedure until the treatments ended. Soluble solids concentrations were subsequently converted to sugar concentration through a linear relationship between oBrix and sucrose concentration (Iland et al. Citation2004).
Berry diameter and dry weight
Prior to each measurement of soluble solids concentration, the diameter of each berry used in the assay above was measured with a microcaliper. This enabled berry volume to be determined assuming the berry was spherical and using the appropriate mathematical formula. This determination was subsequently used along with the sugar concentration to determine the sugar content of each berry, by taking into account the volume of the berry.
At the same time, for each cultivar and treatment, three berries were also removed from the selected bunches (12–15 in total on each occasion) and dried at 60 °C for 2 weeks in a drying oven to determine the dry weight.
Data analysis
All data were analysed using generalised linear models with SAS ver. 9.1.3 (SAS Institute, Cary, NC, USA) as well as using linear regression in some instances. Least-squares means and standard errors were determined and all data were analysed assuming a fully randomised design.
To determine the rates of dry matter accumulation and ripening as a function of temperature, linear regressions were fitted to the berry dry weights and sugar contents against sampling date, using the SAS regression procedure for each bunch, shoot, vine, cultivar and temperature. The slopes of each regression were screened to ensure linearity of response, by selecting those regressions that had a probability (P) <0.05 and variance (r2) >0.75. The slope coefficients were averaged across all bunches, shoots and vines to determine the mean rates of dry matter accumulation and ripening at each temperature for each cultivar. In this context, day temperatures were used because berry expansion occurs during the day (Greer & Rogiers Citation2009) and is highly dependent on photosynthesis (Greer & Weston Citation2010).
Modelling the ripening response
For the temperature-dependent response of ripening in Semillon berries, an empirical cubic regression was fitted to the data of rate of ripening as a function of temperature. Berry ripening and berry diameter in Semillon vines grown in a commercial vineyard were followed at regular intervals from about when the soluble solids concentration was at 4 oBrix (see Greer et al. (Citation2010) for further details of vineyard) by destructively harvesting three berries, located at the top, middle and basal segments of each of 12 bunches and measuring berry diameter and soluble solids concentration, and calculating sugar content, as above. In addition, bunch temperatures in four bunches were measured at hourly intervals with thermocouples inserted inside the bunch shortly after fruit set had occurred. These thermocouples were averaged at hourly intervals throughout the growing season with a data logger (CR1000, Campbell Scientific, Logan, UT, USA). The bunch temperature data were subsequently applied to the temperature-dependent empirical function to estimate the seasonal progression of the ripening process.
Results
Phenology
Budbreak occurred on 7 April in the Semillon vines and on 13 and 14 April in the Chardonnay and Merlot vines, respectively. Merlot vines flowered on 6 May (29 DAB), Chardonnay vines on 7 May (24 DAB) and Semillon vines on 11 May (27 DAB).
Responses of berry growth to temperature
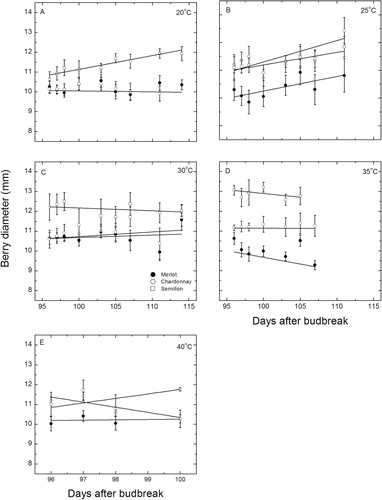
In the post-veraison stage of development when soluble solids increase rapidly, berry diameters changed over time more or less linearly () for some of the cultivars and temperatures. Only the Semillon berries expanded at 20 °C (P = 0.0011, r2 = 0.82) and were significantly (P < 0.01) larger (11–12 mm) than in both the other two cultivars (10 mm) which did not expand at this temperature. At 25 °C, Merlot berries expanded slightly (P = 0.047), whereas the change for Chardonnay was not significant but there were marked increases in berry diameter for Semillon (P = 0.0096, r2= 0.73). There was little evidence of expansion at 30 °C in any cultivar although Semillon berries remained the largest. Some berry shrinkage tended to occur at 35 °C but the changes were not significant; however, there were significant differences in berry size, with Merlot smallest at 9.3 mm, Chardonnay at 11.2 mm and Semillon largest at 13.8 mm. There was much less difference between berries of the different cultivars exposed to 40 °C (), although the Chardonnay berries tended to shrink (P = 0.098, r2 = 0.72) and Semillon berries tended to increase slightly but not significantly.
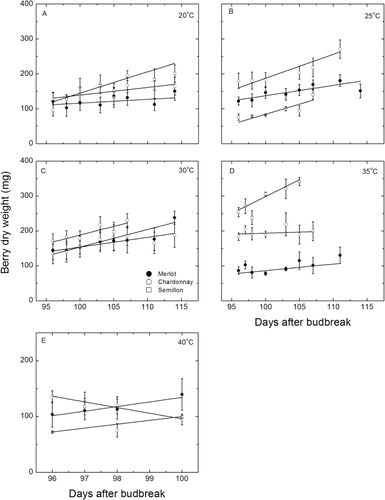
Accumulation of dry weight () in these berries also followed a general linear pattern in each cultivar and temperature, and gains in dry weight occurred over time. At 20 °C, only the Semillon berries increased significantly (P = 0.045, r2 = 0.4) in dry weight (97–201 ± 20 mg) whereas at 25 °C, all cultivars increased significantly (P = 0.0013–0.028; r2 = 0.67–0.87) in dry weight by 80 mg, on average. The same was true at 30 °C, with all the regressions significant (P = 0.003–0.019, r2 = 0.72–0.75) and dry matter increased by 69 mg, on average. Chardonnay and Merlot berries did not increase in dry weight at 35 °C but Semillon berries increased (by 87 mg) significantly in dry weight (P = 0.018, r2 = 0.73). By contrast, only Semillon berries did not change in dry weight when exposed to 40 °C, while the Merlot berries increased (by 35 mg) significantly (P = 0.032, r2 = 0.90) but the Chardonnay berries lost significant amounts (28 mg) of dry weight (P = 0.0096, r2 = 0.97).
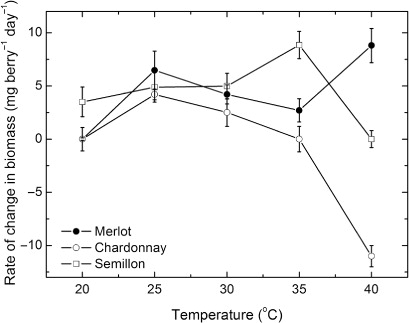
The response of the rate of change in dry matter content to temperature varied between the three cultivars () but was curvilinear in all cases. Rates of accumulation of dry matter in Chardonnay berries varied in a consistent pattern with temperature, with a reasonably distinct maximum of 4.2 mg day−1 at 25 °C, with the rates declining to a negligible rate at 35 °C and to strongly negative values at 40 °C. By contrast, temperatures between 20 and 30 °C seemed to have little impact on the rate of dry matter accumulation in Semillon berries and the rates were about the same magnitude as those for Chardonnay berries. However, Semillon berries accumulated dry matter maximally at 35 °C at a rate of 8.9 mg day−1 and significantly faster than at the lower temperatures and, furthermore, faster than Chardonnay berries. The rate of dry matter accumulation in Semillon berries was negligible at 40 °C. By contrast, the rate of dry matter accumulation in Merlot berries was apparently maximal at 40 °C, while at a similar rate to the maximum rate for Semillon berries. At lower temperatures, the rate of dry matter accumulation in Merlot berries seemed to be also suboptimal at 25 °C, as the rates declined significantly at 20 and 30 °C but the rates at 25 and 40 °C were not significantly different. Thus, the temperature response of the rate of dry matter accumulation in Merlot berries was not distinctive and appeared to be bimodal.
Response of berry sugar accumulation to temperature
Consistent with changes in berry diameter and dry weight accumulation, the accumulation of sugar in the berries across all cultivars and temperatures followed a linear pattern (). At 20 °C, sugar accumulation in Semillon berries increased significantly (P < 0.001, r2 = 0.95) from about 52 to 206 mg berry−1, while for the other two cultivars, there was an increase from 50 to 120 mg berry−1 for Merlot (not significant) and a significant increase from 120 to 150 mg berry−1 for Chardonnay berries (P < 0.001, r2 = 0.78). The sugar content of the Merlot berries was, therefore significantly lower than for the other two cultivars. Across all cultivars, the sugar content increased from 64% to 92% of the biomass.
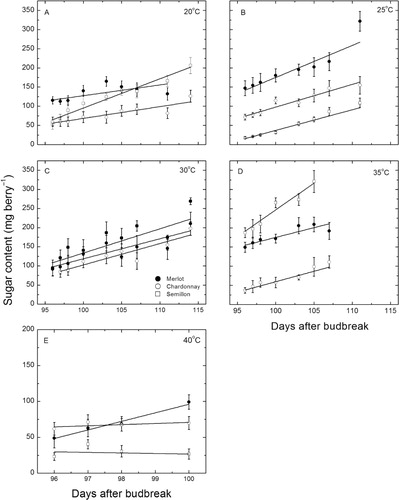
At 25 °C, there were highly significant (P = 0.0001–0.0057, r2 = 0.70–0.97) linear increases in sugar content. Both Merlot and Chardonnay berries approximately doubled the amount of sugar per berry (71 to 144 and 147 to 320 mg berry−1, respectively), whereas in the Semillon berries sugar increased about six-fold (17 to 122 mg berry−1). Thus, again there were significant (P < 0.01) differences between the cultivars in the amount of sugar at the start of the treatment. The sugar content increased from 54% to 88% of the biomass for all cultivars.
By contrast, at 30 °C () there were no differences between the cultivars, but all increased berry sugar content significantly (P < 0.03–0.001, r2 = 0.43–0.88) by about the same amount, from 80–94 ± 16 mg berry−1 to 197—268 ± 18 mg berry−1, over about 20 days and this represented a change from 59% to 92% of the biomass.
At 35 °C, there were linear (P < 0.05–0.0002, r2 = 0.46–0.93) increases in the berry sugar content across all cultivars. Merlot berries increased the most, with sugar content increasing from 37 to 110 mg berry−1; although Chardonnay and Semillon had significantly (P < 0.01) higher amounts (149 and 187 mg berry−1, respectively), the increase was less than two-fold (208 and 320 mg berry−1, respectively). For all cultivars, the sugar content ranged from 61%–93% of the biomass.
At 40 °C, there was no significant (P = 0.35–0.69) change in the sugar content of Chardonnay and Semillon berries (). Across the whole treatment period, berries of these two cultivars averaged 65.5 and 32.7 mg berry−1, respectively, and the difference between them was significant (P < 0.01). However, the Merlot berries increased sugar content significantly (P = 0.017, r2 = 0.95) from 50 to 100 mg berry−1 across the 5 days of treatment. The sugar content of these berries was lower than for the other temperatures at 41% of the biomass and increased to only 62% during the exposures.
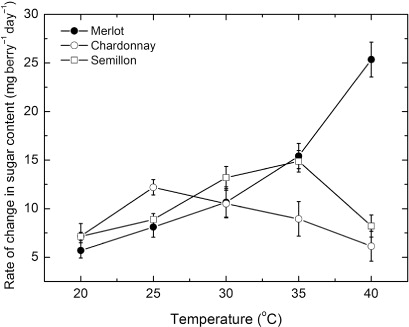
Rate of ripening as a function of temperature
The rates of ripening in response to all temperatures () varied markedly between the three cultivars, with both Chardonnay and Semillon having a distinct but different optimum temperature for ripening. Chardonnay berries ripened maximally at 25 °C and the rate was 12.2 ± 0.8 mg sugar berry−1 day−1 while the Semillon berries ripened maximally at 35 °C and at a rate of 14.8 ± 1.0 mg sugar berry−1 day−1. By contrast, the rate of ripening of the Merlot berries increased exponentially with temperature, and was maximal at 40 °C and at a rate of 25 ± 1.8 mg sugar berry−1 day−1.
Modelling the ripening process
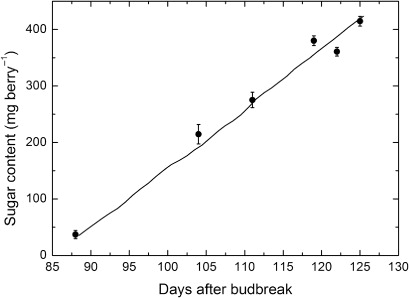
There was a highly significant (P = 0.0102, r2 = 0.999) cubic regression fitted to the rates of ripening and temperature for the Semillon vines (not shown, cf. ). Using the hourly average bunch temperatures, the berry sugar content was estimated from this cubic regression, integrated over the day and then summed over each day of the growing season. This followed a highly linear progression (, slope 10.56 ± 0.03 mg sugar berry−1 day−1, P < 0.0001, r2 = 0.999) which closely matched the measured estimates of sugar content, which also increased linearly (slope 10.04 ± 0.59 mg sugar berry−1 day−1, P < 0.001, r2 = 0.985).
Discussion
Phenology
Growing the three grapevine cultivars in controlled environment conditions has enabled a comparison of their phenology and berry growth traits that otherwise would be difficult to achieve. Comparison of different cultivars in vineyard conditions is the most desirable but because of the phenotype × genotype interaction and the coupling of the growth and development to the prevailing climate, it is difficult to separate differences in performance being cultivar-driven from expressions of the phenotype × environment interaction. The vines of the present study were treated from before leaf fall in the previous season to ensure full dormancy was achieved under identical conditions. This also facilitated a comparison of the phenology of each cultivar, including the time of budbreak and flowering, as well as berry ripening.
On the assumption that V. vinifera cv. Chardonnay was an early maturing variety (McIntyre et al. Citation1982), the dormancy period was extended by about 1 week for these vines in comparison with the other two cultivars in an attempt to synchronise their phenology. However, the Semillon vines broke bud first and the Chardonnay and Merlot vines broke bud 7 days later but almost on the same day as each other. If the extra dormancy treatment was not given to the Chardonnay vines, their budbreak may have been more consistent with that of Semillon but overall, there was little apparent difference in the time of budbreak between the cultivars. Across 11 different cultivars, including Semillon, Riesling, Shiraz and Cabernet Sauvignon and for one region (Riverina, NSW), budbreak varied by 11 days (Coombe & Dry Citation1988). In the study of a large number of cultivars, McIntyre et al. (Citation1982) showed that Chardonnay broke bud 3 days before Semillon, and Merlot was 8 days later. Tomasi et al. (Citation2011) found a 5-day difference, on average, between Chardonnay and Merlot. Thus, the differences in budbreak for the cultivars of the present study fit well within the range for vineyard comparisons of times of budbreak.
The dates of flowering of the three cultivars in the present study were not consistent with those for budbreak. Semillon was the last cultivar to flower but the first to break bud whereas Merlot was the first to flower and the last to break bud. The times for budbreak and flowering of Chardonnay were comparable to that for Merlot. However, Chardonnay flowered at 24 DAB but Semillon and Merlot flowered at 27 and 29 DAB, respectively, and therefore the differences in flowering dates, that is, phenology were extended, with 3–5 days between Chardonnay and the other two cultivars. Consistent with these results, McIntyre et al. (Citation1982) indicated that Chardonnay flowered earlier, by 8 days, than both Semillon and Merlot; however, these authors suggested management practices may have influenced their data. Furthermore, the durations between budbreak and flowering for these cultivars were much shorter than occurs across a number of different cultivars and regions, with the duration more typically over 50 days (McIntyre et al. Citation1982; Coombe & Dry Citation1988; Jones & Davis Citation2000). It is also known that phenology of berry development is faster in controlled environment conditions than in the field (Greer & Rogiers Citation2009). The results of the present study suggest phenology of the three cultivars was increased by the growth conditions and, furthermore, suggests phenology is temperature controlled, certainly for budbreak (Moncur et al. Citation1989) and time to flowering (Tomasi et al. Citation2011).
Berry growth and development
Berry growth and development occurs shortly after fruit set but the major ripening process occurs after veraison and, thus, the temperature treatments in the present study were applied close to just or after veraison (97–107 DAB). This period is when soluble solids and the fresh and dry weights increase in a more or less linear pattern (Coombe Citation1992; Keller et al. Citation1998; Rogiers et al. Citation2006; Shellie Citation2006; Greer & Weedon Citation2012). Certainly, this was the case for all treatments, when berry growth of each cultivar was measured over a relatively short period. It was clear, however, that the cultivars responded differently to temperature, with Merlot berry diameter remaining static over the five temperatures whereas Semillon berries expanded strongly at 20 and 25 °C but not at the higher temperatures. By contrast, the Chardonnay berries expanded rapidly at 25 °C only but perhaps more significantly, berry size declined even more rapidly at 40 °C. Berry volume of Cabernet and Tokay vines was also affected by temperature, with the most rapid increase occurring at 25 °C and the slowest increase at 40 °C (Kliewer Citation1977) and indicates there may be considerable cultivar differences in berry expansion in response to temperature.
Despite the lack of berry expansion, there was still considerable accumulation of dry matter in the post-veraison period, but the rates of accumulation were highly dependent on temperature. There was also considerable variation in the temperature response between the different cultivars, although there was no consistent pattern. Between 25 and 30 °C, nevertheless, the rates for all cultivars were generally about the same at 4.2–5.2 mg (dry weight) berry−1 day−1. For two of the cultivars, Chardonnay and Semillon, there was also a general increase in the rate of accumulation to an optimum and a subsequent decrease in the rate as the temperature increased from 20 to 40 °C. The optimum temperature for dry matter accumulation varied between 25 °C for Chardonnay berries and 35 °C for Semillon berries. The temperature response for the Merlot berries was much less distinct, with markedly lower rates at 30 and 35 °C compared with 25 and 40 °C and, thus, apparently bimodal. Although not expressly calculated, rates of dry matter accumulation in Cabernet Sauvignon berries were estimated to be maximal at about 30 mg berry−1 day−1 at 25 °C and the rates declined thereafter with increasing temperatures to about 19 mg berry−1 day−1 at 40 °C (Kliewer Citation1977). This is in general accordance with the present results. However, it is not known if growth conditions affect the temperature response or the actual rates of dry matter accumulation.
Sugar accumulation by the berries of the different cultivars generally followed a linear pattern across all temperatures and was therefore consistent with the accumulation of dry matter. A similar pattern of glucose accumulation was shown for Cabernet Sauvignon berries on vines grown outdoors (Keller et al. Citation1998) and by about the same magnitude. For the present study, underlying the process of sugar accumulation was the influence of temperature on the rate of ripening. This response varied markedly between the three cultivars, with Chardonnay ripening rates maximal at 25 °C, Semillon maximal at 35 °C and the Merlot berries appeared to be maximal at 40 °C. The patterns of response to temperature for the Chardonnay and Semillon berries were the typical bell-shaped curves of other temperature-dependent processes such as photosynthesis (Taiz & Zeiger Citation2010), although tended towards cooler temperatures for the Chardonnay berries and tended towards the higher temperatures for the Semillon berries. By contrast, the rate of ripening of Merlot berries appeared to increase exponentially in response to temperature, more akin to the response of other processes such as respiration (Sale Citation1974). It is not clear why the ripening process of the different cultivars should vary so much in response to temperature. Chardonnay vines may have originated in the Burgundy region of France (Galet Citation2002). This has a semi-continental climate, with average summer temperatures of about 20 °C. Merlot and Semillon may have originated in the Bordeaux region, where average summer temperatures range from 21 °C in Bordeaux to 25–26 °C in the Gironde region where Semillon is presently grown (Galet Citation2002). It is not certain if the climatic origins of these grapevine cultivars affected the ripening response to temperatures or the vines have acclimated to the local Australian conditions. Certainly, the relatively low temperature maximum for the Chardonnay berries conforms to its origins in the semi-continental climate of France, but the relatively high maxima of the other two cultivars would appear to reflect more the growth conditions because the temperature maxima for berry ripening were too high for their climatic origins. Nevertheless, because the cultivar differences in temperature response for ripening were assessed in common growth conditions, the variation must have at least some genotypic origins. Further study is certainly required to assess what contribution acclimation to the growth conditions might have occurred in determining these ripening responses to temperature.
The rates of ripening of the three cultivars were typically in the range of 5–15 mg sugar berry−1 day−1. Previous estimates of the rate of ripening for Semillon were about 8 mg berry−1 day−1 at 25 °C (Greer & Weston Citation2010) and well in keeping with the rates of the present study. Similarly, rates estimated for Cabernet Sauvignon and Tokay berries at about 6 and 16 mg berry−1 day−1 and maximal at 32.5 °C and 35 °C, respectively (Kliewer Citation1977), and for the cultivar Chasselas grown in outdoor conditions, estimated from Dreier et al. (Citation2000), at 11 mg berry−1 day−1, were also in accord. In contrast, for Cabernet Sauvignon vines grown outdoors, the rate of glucose accumulation was estimated to be 2.8 mg berry−1 day−1 (Keller et al. Citation1998) and perhaps indicative of growth conditions having an influence on the rates of ripening. Sadras et al. (Citation2008) in their study of dynamics of soluble solids, nevertheless, showed that Chardonnay, in comparison with Merlot and Semillon, ripened earlier and was physiologically mature sooner.
A quantitative relationship between the rates of ripening and temperature was established for the Semillon vines, although based on an empirical function. Nevertheless, it was clear that applying this relationship to an independent set of bunch temperatures collected in a vineyard was used successfully to determine the accumulation of sugar in the berries in field-grown vines. In both the modelled accumulation and the measured content of berry sugar, the patterns of ripening followed linear progressions, with a close match between them, confirming a good fit. This model clearly confirms that the ripening process across the growing season is driven by an intrinsic relationship between temperature and rate of ripening. It seems likely that this would hold for the other cultivars in the study and possibly more generally in the Vitis genus. We can also conclude more generally, that the use of controlled environments to develop underlying temperature-dependent functions of biological properties is vindicated as a highly amenable approach and, furthermore, such functions can be readily translated to field-grown plants in varying temperature conditions.
In conclusion, this study has established that the ripening process of selected grapevine cultivars was a temperature-dependent and curvilinear response. Furthermore, the actual quantitative response to temperature differed between the cultivars, with the optimum temperature for ripening varying widely from 25 to 40 °C. The accumulation of dry matter, of which the sugar is a major component (50%–90% over time) was also influenced by temperature and generally affirmed the different temperature responses between the three cultivars. While there may be genotypic differences between the cultivars in their response to temperature, it is likely that the growth conditions also had a major influence on the response and certainly warrants further investigation. However, these temperature-dependent responses provide a basis for assessing the impact of the changing climate on the phenology and ripening process of grapevines in different regions. Furthermore, the rates of ripening for the three cultivars were comparable with other varieties. The results for Semillon provided a basis for successfully modelling the ripening process over the growing season and will provide the basis for the more extensive modelling currently being undertaken at our institute. The results also affirm that Chardonnay is more likely to be a cool season cultivar and is generally intolerant of very high temperatures, whereas Semillon and Merlot can be concluded to be warm season cultivars and well tolerant of high temperatures in the ripening and growth processes of their berries.
Acknowledgements
This project was funded from a Winegrowing Futures grant to the National Wine and Grape Industry Centre from the Grape and Wine Research and Development Corporation. We acknowledge the contribution of Pete Carey for technical assistance with the project. We also thank the vineyard owners for allowing the research to be conducted on their vines. The senior author also acknowledges support from SAS Australia.
References
- Bergqvist J, Dokoozlian N, Ebisuda N 2001. Sunlight exposure and temperature effects on Berry Growth and composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. American Journal of Enology and Viticulture 52: 1–7.
- Buttrose MS, Hale CR, Kliewer WM 1971. Effect of temperature on the composition of ‘Cabernet Sauvignon’ berries. American Journal of Enology and Viticulture 22: 71–75.
- Chorti E, Guidoni S, Ferrandino A, Novello V 2010. Effect of different cluster sunlight exposure levels on Ripening and Anthocyanin Accumulation in Nebbiolo Grapes. American Journal of Enology and Viticulture 61: 23–30.
- Coombe BG 1992. Research on development and Ripening of the Grape Berry. American Journal of Enology and Viticulture 43: 101–110.
- Coombe BG 1995. Growth stages of the grapevine: adoption of a system for identifying grapevine growth stages. Australian Journal of Grape and Wine Research 1: 104–110. 10.1111/j.1755-0238.1995.tb00086.x
- Coombe BG, Dry PR 1988. Viticulture volume I resources. Adelaide, Winetitles. 211 p.
- Costa JM, Ortuño MF, Lopes CM, Chaves MM 2012. Grapevine varieties exhibiting differences in stomatal response to water deficit. Functional Plant Biology 39: 179–189. 10.1071/FP11156
- Crippen DD, Morrison JC 1986. The effects of sun exposure on the phenolic content of cabernet Sauvignon Berries during development. American Journal of Enology and Viticulture 37: 243–247.
- Dreier LP, Stoll GS, Ruffner HP 2000. Berry Ripening and Evapotranspiration in Vitis vinifera L. American Journal of Enology and Viticulture 51: 340–346.
- Galet P 2002. Grape varieties. London, UK, Cassell Illustrated. 159 p.
- Gladstones J 1992. Viticulture and environment. Underdale, South Australia, Winetitles.
- Greer DH 2012. Modelling leaf photosynthetic and transpiration temperature-dependent responses in Vitis vinifera cv. Semillon grapevines growing in a hot irrigated vineyard conditions. AoB Plants: pls009. doi:10.1093/aobpla/pls009
- Greer DH, Rogiers SY 2009. Water flux of Vitis vinifera L. cv. Shiraz bunches throughout development and in relation to late-season weight loss. American Journal of Enology and Viticulture 60: 155–163.
- Greer DH, Sicard SM 2009. The net carbon balance in relation to growth and biomass accumulation of grapevines (Vitis vinifera cv. Semillon) grown in a controlled environment. Functional Plant Biology 36: 645–653. 10.1071/FP09037
- Greer DH, Weedon MM 2012. Interactions between light and growing season temperatures on growth and development and gas exchange of Semillon (Vitis vinifera L.) vines grown in an irrigated vineyard. Plant Physiology and Biochemistry 54: 59–69. 10.1016/j.plaphy.2012.02.010
- Greer DH, Weedon MM, Weston C 2011. Reductions in biomass accumulation, photosynthesis in situ and net carbon balance are the costs of protecting Vitis vinifera ‘Semillon’ grapevines from heat stress with shade covering. AoB Plants: plr023. doi:10.1093/aobpla/plr023
- Greer DH, Weston C 2010. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Functional Plant Biology 37: 206–214. 10.1071/FP09209
- Greer DH, Weston C, Weedon MM 2010. Shoot architecture, growth and development dynamics of Vitis vinifera cv. Semillon vines grown in an irrigated vineyard with and without shade covering. Functional Plant Biology 37: 1061–1070. 10.1071/FP10101
- Iland P, Bruer N, Edwards G, Weeks S, Wilkes E 2004. Chemical analysis of grapes and wine: techniques and concepts. Campbelltown, South Australia, Patrick Iland Wine Promotions PTY Ltd. 110 p.
- Jones GV, Davis RE 2000. Climate influences on grapevine phenology, grape composition, and wine production and quality for Bordeaux, France. American Journal of Enology and Viticulture 51: 249–261.
- Jones GV, Duff AA, Hall A, Myers JW 2010. Spatial analysis of climate in winegrape growing regions in the Western United States. American Journal of Enology and Viticulture 61: 313–326.
- Keller M, Arnink KJ, Hrazdina G 1998. Interaction of nitrogen availability during bloom and light intensity during veraison. I. effects on grapevine growth, fruit development, and ripening. American Journal of Enology and Viticulture 49: 333–340.
- Kliewer WM 1977. Effect of high temperatures during the bloom-set period on fruit-set, ovule fertility, and berry growth of several grape cultivars. American Journal of Enology and Viticulture 28: 215–222.
- Matsui S, Ryugo K, Kliewer WM 1986. Growth inhibition of Thompson seedless and Napa Gamay berries by heat stress and its partial reversibility by applications of growth regulators. American Journal of Enology and Viticulture 37: 67–71.
- McIntyre GN, Lider LA, Ferrari NL 1982. The chronological classification of grapevine phenology. American Journal of Enology and Viticulture 33: 80–85.
- Moncur MW, Rattigan K, Mackenzie DH, McIntyre GN 1989. Base temperatures for budbreak and leaf appearance of grapevines. American Journal of Enology and Viticulture 40: 21–26.
- Moutinho-Pereira J, Magalhães N, Gonçalves B, Bacelar E, Brito M, Correia C 2007. Gas exchange and water relations of three Vitis vinifera L. cultivars growing under Mediterranean climate. Photosynthetica 45: 202–207. 10.1007/s11099-007-0033-1
- Radler F 1965. The effect of temperature on the ripening of Sultana grapes. American Journal of Enology and Viticulture 16: 38–41.
- Rogiers SY, Greer DH, Hatfield JM, Orchard BA, Keller M 2006. Solute transport into Shiraz berries during development and late-ripening shrinkage. American Journal of Enology and Viticulture 57: 73–80.
- Rogiers SY, Greer DH, Hutton RJ, Landsberg JJ 2009. Does night-time transpiration contribute to anisohydric behaviour in a Vitis vinifera cultivar? Journal of Experimental Botany 60: 3751–3763. 10.1093/jxb/erp217
- Sadras VO, Collins M, Soar CJ 2008. Modelling variety-dependent dynamics of soluble solids and water in berries of Vitis vinifera. Australian Journal of Grape and Wine Research 14: 250–259.
- Sale P 1974. Productivity of vegetable crops in a region of high solar input. III. Carbon balance of potato crops. Functional Plant Biology 1: 283–296.
- Sepúlveda G, Kliewer WM 1986a. Effect of high temperature on grapevines (Vitis vinifera L.). II. Distribution of soluble sugars. American Journal of Enology and Viticulture 37: 20–25.
- Sepúlveda G, Kliewer WM 1986b. Stomatal response of three grapevine cultivars (Vitis vinifera L.) to high temperature. American Journal of Enology and Viticulture 37: 44–52.
- Shellie KC 2006. Vine and berry response of merlot (Vitis vinifera L.) to differential water stress. American Journal of Enology and Viticulture 57: 514–518.
- Shellie KC 2011. Interactive effects of deficit irrigation and berry exposure aspect on merlot and cabernet Sauvignon in an arid climate. American Journal of Enology and Viticulture 62: 462–470. 10.5344/ajev.2011.10103
- Smart RE 1973. Sunlight interception by vineyards. American Journal of Enology and Viticulture 24: 141–147.
- Smart RE, Sinclair TR 1976. Solar heating of grape berries and other spherical fruits. Agricultural Meteorology 17: 241–259. 10.1016/0002-1571(76)90029-7
- Smart RE, Smith SM, Winchester RV 1988. Light quality and quantity effects on fruit ripening for cabernet sauvignon. American Journal of Enology and Viticulture 39: 250–258.
- Soar CJ, Collins MJ, Sadras VO 2009. Irrigated Shiraz vines (Vitis vinifera) upregulate gas exchange and maintain berry growth in response to short spells of high maximum temperature in the field. Functional Plant Biology 36: 801–814. 10.1071/FP09101
- Sommer KJ, Islam MT, Clingeleffer PR 2000. Light and temperature effects on shoot fruitfulness in Vitis vinifera L. cv. Sultana: influence of trellis type and grafting. Australian Journal of Grape and Wine Research 6: 99–108. 10.1111/j.1755-0238.2000.tb00167.x
- Spayd SE, Tarara JM, Mee DL, Ferguson JC 2002. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot Berries. American Journal of Enology and Viticulture 53: 171–182.
- Taiz L, Zeiger E ed. 2010. Plant physiology. 5th edition. Sunderland, Massachusetts, USA, Sinauer Associates Inc.
- Tomasi D, Jones GV, Giust M, Lovat L, Gaiotti F 2011. Grapevine phenology and climate change: relationships and trends in the Veneto Region of Italy for 1964–2009. American Journal of Enology and Viticulture 62: 329–339. 10.5344/ajev.2011.10108
- Weaver RJ, Iwasaki K 1977. Effect of temperature and length of storage, root growth and termination of bud rest in Zinfandel Grapes. American Journal of Enology and Viticulture 28: 149–151.
- Yamane T, Jeong ST, Goto-Yamamoto N, Koshita Y, Kobayashi S 2006. Effects of temperature on anthocyanin biosynthesis in grape berry skins. American Journal of Enology and Viticulture 57: 54–59.