
ABSTRACT
Whilst transhumance has been researched as a livestock and land management system, its legacy on soil- nutrient levels is unknown. This study addresses changes in soil chemistry associated with historical transhumance grazing management with the aim of establishing if soil-nutrient levels can shed new insights into land-use legacies originating from past transhumance in Scotland. The study is focused on one aspect of transhumance, the shielings associated with milking livestock formerly operating in the uplands of the Scottish Midland Valley. A literature review is presented focusing on the history, functioning and geographic distribution of transhumance systems in Scotland set within a global context. This is followed by a detailed examination of two shieling sites in Menstrie Glen in the Ochil Hills. Their relationship to topography is considered, with vegetation survey and chemical analyses for plant macronutrients of soils collected in the vicinity also carried out. Underpinned by theories integrating biogeochemical fluxes and agropastoral activities it is established that there is moderate enrichment of local nutrient levels still evident at the Menstrie Glen shieling sites today. Review of historical and environmental sources suggests that local nutrient levels reflect temporal dynamics of shieling establishment, functioning and abandonment with limited evidence to suggest this was driven by population pressure and adaptation to Little Ice Age climate change. The analyses lead to insights about wider ecological impacts of different forms of land use in the face of climate change and population pressure, carrying implications for land use in Scotland and the challenges facing extant transhumance systems in countries where both these issues are pressing today.
INTRODUCTION
Transhumance is a globally widespread system of pastoralism. Whilst many differences in practice exist locally, a common element is the seasonal movement of livestock and herders over either horizontal, or more frequently, vertical, distances to access meadows becoming available and productive at a certain time of the year. Regularly this means the migration upwards into mountainous areas to take advantage of the plant growth on high pastures during summer. As such, transhumance opens up areas with prevailing environmental conditions unsuitable to stationary year-round agricultural or pastoral systems, be that due to low temperatures, lack of rain or seasonal floods. The practice can be differentiated from pure nomadism, with its more peripatetic lifestyle, by the existence of a permanent home base, in use exclusively during the winter or more commonly all year round by some members of the family who will stay behind at this base. The low-lying areas vacated by livestock are then used for the production of winter fodder or for arable farming, whilst the principal products of the higher altitude summer grazings are butter and cheese from the management of milking livestock or the regulated fattening of livestock. Transhumance can further be differentiated from common land grazing and agistment, a practice of driving livestock onto summer pastures where landowners are often paid per head of livestock and the latter is largely left to fend for itself (Winchester Citation2000, p. 93). Transhumance may thus be seen as a strategy to maximise and optimise the utilisation of natural resources in relation to pastoral activities, both over space and time, by ensuring animals are in the right place at the right time to access these resources and to not hinder arable farming or winter fodder production in the valleys. Alongside the animals, the herders, be it single individuals or whole extended families, shift their home in the same seasonal pattern, inhabiting semi-permanent or permanent buildings whilst doing so (Evans Citation1940, Citation1958; Davies Citation1941; Dixon Citation2007). Whilst in most cases the owners of the livestock are also those who will make this migration, Fox (Citation2012, pp. 46–107) describes what he terms impersonal transhumance, one where professional grazers cared for the livestock during the time they were kept at the summer pastures. A similar system can be found in the Alborz mountains today, where the Iranian owners of livestock employ Afghani immigrants to attend to the animals at their summer pastures high in the mountains (pers. observation SW, 2017, 2018, 2019).
Some authors separate transhumance into vertical and horizontal transhumance. The former being the already highlighted movement with the seasons to higher elevation, the latter — and some consider this to be the only form of ‘true’ transhumance — encompassing often longer distances over mostly horizontal ground, a practice especially prevalent in countries bordering the Mediterranean (cf. Hofmeister Citation1961; Costello & Svensson Citation2018; see also Fox Citation2012, pp. 27–42 who terms this ‘lesser’ for mainly vertical, and ‘greater’ for mainly horizontal transhumance). In this paper, both are considered as specific forms of the same practice, whilst it is acknowledged that transhumance must be seen as embedded within a continuum from nomadism to permanent settlement and that there is and has been in many instances a degree of admixture where these forms of living are morphing into each other. Shepherd (Citation2021) also points out for late medieval Buchanan in north-east Scotland that a sharp contrast between upland and lowland regimes simplifies a more fluid situation, especially prior to widespread enclosure.
Transhumance involves an often profound input of labour; accessing the high pastures alone requiring significant effort, yet the prevalence of this form of agropastoralism in mountain areas, asserts its effectiveness in ensuring sufficient returns in food and economic gain (Dodgshon & Olsson Citation2007).
Today, systems of transhumance can be found worldwide. Within Europe one of the most well- known examples is the still widespread Almwirtschaft in the Alps, with its elaborate traditions surrounding the yearly migration to and from the high meadows on the flanks of Alpine peaks. Vertical transhumance is also widespread in the high mountain ranges of Asia and is evident in Mongolia, parts of China, Bhutan, Nepal, northern India, Pakistan, and Afghanistan amongst others (Suttie & Reynolds Citation2003). Transhumance is present in the western USA, where it can still be found to operate on the level of individual farms (Huntsinger et al. Citation2010). Neither is the practice solely a surviving relict of former times. New areas of transhumance can for example be found in Armenia, where Yezidis displaced by Daesh from their homelands in the Sinjar-mountains of Iraq have resettled and imported transhumance to the slopes of Mount Aragats (pers. observation SW, 2019).
In the mountainous parts of Britain and Scotland in particular, transhumance is an element of the historical landscape and associated with the shieling. This modern name is derived from Middle English shale and Old Norse skjol, defining the building for seasonal human occupation as well as the unenclosed upland pasture associated with it. According to the current Canmore dataset, the national record of historic monuments of Scotland, there are 7,267 shielings (Canmore 2020). Many of these were first mapped by the Ordnance Survey, whilst a substantial increase in known sites occurred through the Rapid Afforestable Land Survey (ALS), initiated in 1989, following an acknowledgement by the government to map any sites of archaeological or other historical interest prior to granting permission for afforestation (Dixon Citation2018). In order to separate the remains of shielings from the remains of huts of different usage an arbitrary size cut-off was used by ALS, together with other evidence, such as the presence of midden-mounds, the absence of compartmentalisation of the buildings and the occurrence of them in a group (Dixon Citation2018). This approach must, to a certain degree, lead to the misidentification of some structures and does not consider complexities arising from the reuse and modification of former shieling sites at a later date.
Whilst their location has been extensively documented and changes in function recognised, the agropastoral, economical and socio-ecological practices supporting the shieling-system have been less well studied, as has the influence shielings had on their environment, both in terms of the effects which ensued at the time of active use and regarding the long-term ecological impacts (cf. Hamilton & Davies Citation2007).
Furthermore, whilst many predictions about future changes of ecosystems or landscapes are still based solely on extrapolating from the current state, a growing body of work highlights the necessity to also consider the socio-ecological past to better predict the future. These considerations of land-use legacies frequently consider species composition, richness and abundance as well as geomorphology (Swetnam et al. Citation1999; Foster et al. Citation2003; Perring et al. Citation2016), whilst recent reviews have highlighted the importance of legacy effects on soilscapes and the long-lasting signatures human activity can impose upon the biogeochemical properties and cycles therein (McLauchlan Citation2006; MacDonald et al. Citation2012).
Despite this, and despite an increasing demand for meat worldwide and thus an increasing import- ance of pastures, the impact of livestock upon these pastures remains largely understudied (cf. Erb et al. Citation2016). The few studies considering the long- term legacies of past pastoral practices have mostly focused on the aspects of erosion and to a lesser degree deforestation (for example Brown et al. Citation2012 and EddudÓttir et al. Citation2020 for sites in Iceland; Nedelea (Citation2009) for a discussion of erosion impacts of contemporary transhumance in Romania). Some limited research on soil nutrients in Austria hints at a surprisingly high land-use intensity and possible N-depletion in traditional high alpine valley systems (Gingrich et al. Citation2015). Battaglini et al. (Citation2014) also highlight the high variability of soil nutrient stresses within the same agropastoral system.
The influence of former shielings on the environ- ment presented itself primarily in the form of biomass alteration, i.e., the uptake of vegetation by grazing livestock and fertilization through the excreta of the animals (cf. McLauchlan Citation2006). The aim of this study is thus to identify legacy-effects of these two influences on soilscapes, soil chemistry and vegetation associated with shielings in upland areas of the Scottish Midland Valley during the post-medieval period. From this environmental evidence one can deduct that whilst transhumance can be environmentally sustainable, the opposite may also hold true, with its impact intensity especially dependent on many local and farm- level components. Insights about the edaphic and vegetational sustainability of former shielings can thus help to inform models for environmentally sustainable (organic) low-impact (low input, low output) pastoralism at the present day (cf. Casado & Molina Citation2009; cf. Antrop Citation2005; Schiavon et al. Citation2019). Set within the broader human ecology of transhumance activity it is anticipated that the soil and vegetational legacies examined here will provide a new tool to study the wider environmental impact and sustainability of pastoral activities in rangeland- anthromes and their resilience against changing climates and population pressures.
The literature on transhumance in general, and shielings in particular, is first reviewed to build an up-to-date understanding about this form of agropastoralism. A second section describes the methods used to conduct a locational analysis of shielings in the Ochils and Menstrie Glen, an upland area of igneous bedrock in the Scottish Midland Valley where there has been little study of shielings, enabling an overview of their position in the agropastoral and environmental history of the area. This is followed by a description of how the relationship of two selected shielings to topography, vegetation and pedology was assessed. The results of these analyses are presented in a third section. In the fourth and discursive section, the paper discusses the assessment of land-use legacies of these shielings, as evidenced in the signatures of soil fertility resulting from grazing pressure and animal nutrient input in their vicinity. The paper goes on to consider the human ecology of former shielings, their relation to population pressure and climatic factors during the Little Ice Age and the implications this generates for a wider discussion about past and current land- use practices.
LITERATURE REVIEW
transhumance: the historical evidence
Despite being a widespread practice today, the historical aspects of transhumance have been the subject of relatively few systematic studies. This can be seen in the light of a general neglect of the study of the culture of populations living amongst mountains, which are sometimes seen as a poorer subset of populations living in the lowlands, both in terms of their perceived socio-cultural practices and the perceived value of these (cf. Dodgshon Citation2009). Evidence has been obtained for the seasonal movement of livestock to high pastures in the European Alps as early as the late third millennium b.c. (Reitmaier 2012; Reitmaier et al. Citation2018). The research of these distant beginnings of transhumance is complicated by the often very fragmentary nature of any kind of remains suitable for gleaning information about the agropastoral practices at the time. Akeret and Jacomet (Citation1997) have highlighted in their study of faecal macrofossils from Neolithic low-level farms at Horgen Scheller, Switzerland, the absence of certain kinds of plant remains, such as anthers (the part of a flower carrying the pollen), which would be associated with the summer months, in addition to the remains coming from species accessible in winter. One possible explanation for this could be transhumance removing the animals from the site during summer, yet more evidence is needed to rule out other possibilities. Other researchers have used zooarchaeological and geoarchaeological methods to reconstruct the presence and movement of livestock on high ground in the French Pyrenees during the Neolithic. From the available evidence it seems likely that these practices were in conjunction with hunting wild animals on higher ground and this may indeed have been one of the prime reasons for venturing there, together with the degradation of the lower- lying ground and an expansion of the population, the latter however being very difficult to ascertain (Geddes Citation1983). Some of the best evidence for the presence of regular seasonal rather than sporadic utilisation of upland Alpine pastures comes from a high-altitude mire on Mont Fallère, Italy, where palaeoecological multi-proxy studies were compared to nearby 14C-dated archaeological sites and led to the conclusion that from as early as c. 3600 b.c. humans were responsible for the transformation of high alpine woodland into pastures (Pini et al. 2017). Given that these would have been covered in snow during the winter one may imply the presence of some form of transhumance.
In Britain, palynological and geoarchaeological examinations at Greater Langdale in the English Lake District brought to light indirect and tentative evidence for the presence of domesticated animals in these uplands during the Neolithic, from which some authors deduct the presence of seasonal upland grazing regimes (Bradley & Edmonds Citation1988). However, Bradley (Citation1978) and Chambers et al. (1988), highlight, in several case studies, the difficulty of clearly separating archaeological and palynological evidence pointing at seasonal but irregular usage of areas from transhumance with a consistent seasonal regime. Clearer evidence for transhumance is given by new research analysing historic place-names in Cumbria, suggesting a prominent role for Scandinavian settlers establishing settlements in the central Lake District with the help of transhumance (Foster Citation2021).
Shieling establishment in Scotland is found in Norse setr, sætr and ærgi (adopted from Scottish Gaelic). Norse place-names indicate Norse period origins (Foster Citation2018). Archaeological evidence is scarce (cf. Dickinson Citation2020) and in Scotland is based on single cases, such as the remains of a thirteenth– fifteenth-century shieling on Barra. However, this lack of evidence may be because dwellings built prior to the seventeenth century were most frequently constructed from turf, with the obvious implications for the preservation of such structures (cf. Atkinson Citation2010). In addition, there is a parallel lack of documentary evidence, which, prior to the late medieval period is commonly only in the form of grants given to grazing rights and similar resources from which the actual practice of transhumance may or may not be inferred (Mather Citation1970; Dixon Citation2018). Dixon further points out that the most extensive studies of transhumance in Scotland, foremost that of Bil (Citation1990), are, due to the more readily available documentary evidence in the form of estate-papers, focusing on land use from the seventeenth century onwards, a time which saw the waning of the shieling system and the emergence of year-round sheep farming, made possible by the introduction of hardier breeds, such as Cheviots and Blackfaces (Miller Citation1967). However, Bil (Citation1990, p. 3) also traces references to shielings back to the twelfth century, which coincides with the expansion of population observed by others (Anderson Citation2001; cf. Flinn et al. Citation1977).
What we know about the specifics of animal husbandry occurring at shielings has in many instances been deducted from documentary evidence. The time the animals spent at the shieling was usually quite short, with the barony courts establishing the dates at which livestock had to be removed from the farm and brought to the shieling, usually mid to end June, and the time when the animals were supposed to be brought back at the end of the growing season. The latter was often undertaken in increments, with some cattle being brought back as early as early August, so as to be available for the tathing of outfields with their dung. The last animals were brought down by late September, hence the maximum time the animals spent at the shieling would be about four months (Dodgshon & Olsson Citation2006) whilst periods of one to two and a half months have been reported for Perthshire (Bill 1990, p. 177). Documentary evidence for shielings on the flanks of Ben Lawers in the Central Highlands suggests periods from the beginning of May to the middle of July (Atkinson Citation2016). Census data also reveals that stocking levels in pre-clearance shieling sites were very low compared to those established thereafter in year-round upland grazing. Contrary to a widespread belief that pre-clearance stocks were solely composed of cattle, stocks were actually mixed, with sheep, goats and, to a lesser degree, horses all being present (Dodgshon & Olsson Citation2006; cf. Campbell Citation2008 for the importance of sheep for the Scottish economy during the thirteenth and fourteenth century before a later transition to cattle). Identifying the presence of domestic livestock type at individual former shieling sites is yet to be achieved although faecal biomarkers in soils offer novel science-based approaches to this issue (Simpson et al. Citation1999).
The shieling huts themselves were built in varying designs, from beehive corbelled structures and roundhouses to cottage-like buildings in more recent times. Construction would typically involve a stone footing with turf walls and roofs (Wilkinson Citation2009). Shielings are frequently located at a well- drained spot of relatively level ground and close to a water source, usually a burn, commonly in groups of a handful to a dozen huts. In some cases however, such as at a site near Ben Lawers, up to eighty to ninety of them can be found in close proximity (Dixon Citation2018). It has not been studied to what degree this was influenced by the density of the population in the valleys.
What little reconstructed information we have for shielings at different locations in Scotland paints a picture of disparity regarding how the shieling was embedded within the wider agropastoral system. For the Isle of Skye, for example, it has been noted that indeed not all townships held shieling sites, and that at those having enough low-lying ground to support their milch-cows, the latter were kept on that instead, whilst there are indications that cattle were left on the high ground in a system better described as agistment (MacSween Citation1959). In Assynt, by contrast, shielings seem to have frequently been in essence outfields, or turned into these over the course of time, respectively (Miller Citation1967). Elsewhere, the extensive proliferation of shielings can be seen as a direct result of increasing population pressure, such as on Rum, where the population reached 445 at the end of the eighteenth century and where calculations show that every family had to utilise at least two shieling sites in order to gain enough feed for their livestock from the poor soils of the island (Miller Citation1967).
On the slopes of Ben Lawers, shielings occurred in great abundance at different altitudes. The evidence here hints at frequent re-allocation of shielings to different households and the moving of livestock from one shieling to another midway through the grazing season (Atkinson Citation2016). Whilst shielings show a general decline in the seventeenth century and their usage was dying out in many parts of Scotland soon thereafter (Smith Citation1986), they survived longest in the Western Isles, where the last shielings gradually disappeared during the interbellum (Grant Citation1977, pp. 25ff).
These examples show that the roles of shielings within the human ecology of the specific place in which they were embedded, whilst not funda- mentally dissimilar to each other, were far from uniform; equally the location of shielings was not the same everywhere, with those on the west coast often close to the sea, whilst in eastern parts of Scotland shielings could be at an altitude in excess of 700 metres above sea level (Smith Citation1986). It remains to be studied to what degree these observable differences translate into different kinds of land use, soilscape and sustainability legacies.
METHODS
site selection
Knowledge about shielings in the hill areas of the Scottish Midland Valley is limited, being confined to an understanding of their distribution. For this study, Menstrie Glen in the Ochils, a range of Devonian Age igneous hills rising steeply out of the Midland Valley north of the River Forth, was chosen as an example from this geographical region. The historical monuments within Menstrie Glen, including shielings sites, are well mapped and a general study about the history of human settlement of the glen had already been conducted (RCAHMS 2001) providing important baseline information on which this present study could draw. Menstrie Glen has eleven shieling sites (, ). Considering these with the information provided by the Canmore dataset (Canmore 2020a–k) and the impact of recent land use changes, such as afforestation (cf. Gallacher et al. Citation2013), confirmed by a field visit to all shieling sites within the glen, sites A and H were selected for in-depth examination.
Fig. 1. Location of all shieling sites within Mentrie Glen and Scotland. Canmore data on OS base map.
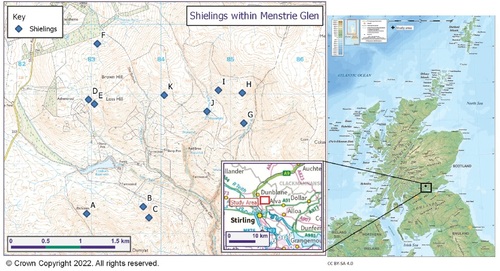
TABLE 1. OVERVIEW OF ALL ELEVEN SHIELINGS WITHIN MENSTRIE GLEN (CANMORE 2020A–K).
vegetation survey
Vegetation-cover at the shieling sites was surveyed according to the JNCC Phase 1 habitat survey classifications (JNCC 2010) and based on on-the- ground survey by transect walking. Plants were identified with standard field-guides (Hubbard Citation1984 for monocots; Fitter et al. Citation1996 for dicots).
soil sampling
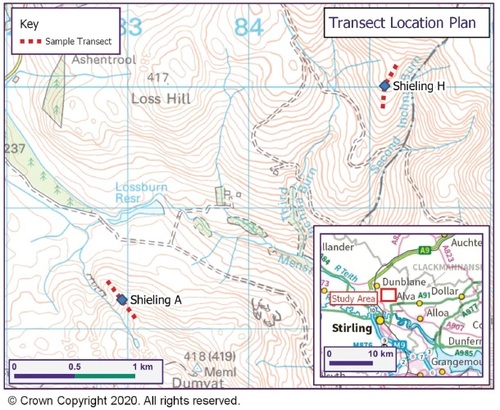
Sites A and H were located using GPS and OS maps. A centre point for the shieling structures was established and a 400 m transect was placed (), 200 m uphill and 200 m downhill. At site H the uphill transect was later expanded to 300 m to gain more information on the tailing off of nutrient levels in this particular place. Soil was sampled and placed in plastic bags at ten points per transect, 10 m, 50 m, 100 m, 150 m and 200 m in each direction. (Noted as -10 etc. for downslope and as +10 etc. for upslope sampling points.) Distances were paced out and if necessary corrected by GPS. The stratigraphy of the soil was examined using a Dutch soil auger. Soil-colour was established with a Munsell-chart. Soil samples were taken at 10 cm soil depth, which was chosen for the main analysis in order to access the mineral soil underneath the more recent thin but highly organic topsoil and to limit the influence of later stage nutrient inputs on the test-results ( and ). At the respective +10 sampling point at each site samples were taken at 2 cm, 10 cm and then every 10 cm downwards to establish the nutrient range within the soil-column. All sampling was carried out between October 2020 and May 2021.
Fig. 2. Location of shieling transects A and H within Menstrie Glen
chemical analysis
A Hanna Instruments field test-kit was used to measure plant available nutrient levels for the soils sampled to reflect intensities of past human activity (Holliday & Gartner Citation2007; Eidt Citation1984; Zlateva et al. Citation2018). A colorimetric approach was applied to measure pH (Barium Sulfate, Calcium Chloride reagent), N (Barium Sulfate, Citric Monohydrate, Potassium Disulfate reagent) and P (Potassium Disulfate reagent) levels. For K levels a turbidimetric approach was applied (EDTA Tetrasodium salt, Sodium Tetraphenylborate, Lithium Hydroxide reagent) (cf. Hanna Instruments Citation2018). This field- based approach allowed the grouping of results on an ordinal scale (seven increments from trace to high), facilitating the emergence of general trends in nutrient-loads.
RESULTS
vegetation survey
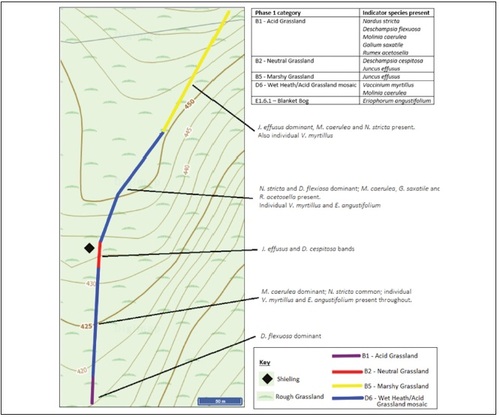
Site A has a relatively species-poor vegetation cover of acid grassland, heathland and a patch of neutral grassland occupied by Juncus effusus and Deschampsia cespitosa, species resistant to grazing (). The vegetation cover at site H presents a slightly higher diversity (ten versus only eight species at site A) and a mosaic of habitat types (). Acid grassland/ heath mosaic dominates, morphing into marshy grassland at the upper end of the transect. The forestry plantation nearby is likely reducing the moisture-level of the soil and will in the future due to shading and the water- needs of the growing trees further influence the vegetation.
Fig. 3. Phase 1 mapping and occurrence of indicator species with specific locations for selected species at shieling site A.
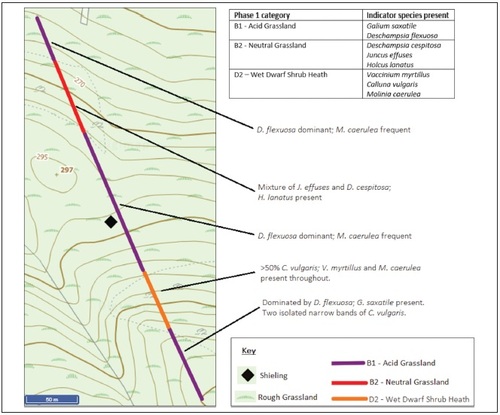
Fig. 4. Phase 1 mapping and occurrence of indicator species with specific locations for selected species at shieling site H.
topographical and pedological analysis
Site A () occupies a position just underneath a steep promontory with its topographic position possibly suggesting that it is located on a created platform. Apart from this, the general slope angle is between 15 and 20 degrees throughout, with the main slope facing north-west. A shallow A-horizon of reddish hued peaty loam (Munsell colour: 7.5 YR 4/2) overlies a B-horizon of sandy clay loam (Munsell colour: 7.5 YR 2.5/3) resting directly on the Devonian Age igneous bedrocks. The A-horizon is moderately well drained, with slightly more water being present towards the downslope end of the transect and some gleying taking place. The B-horizon is well drained throughout. The soil type at site A is Umbrisol according to the World Reference Base for Soil Resources (WRB) or Brown Earth according to the Scottish Soil Classification.
Fig. 5. Combined slope diagram, soil column diagram and nutrient analysis report for shieling site A. The slope diagram shows the topography along the transect of site A. Soil columns visualise the stratigraphy of the soil including different soil horizons and their respective colour. Both tables visualise the results of the chemical analysis at the sampling points along the transect at a soil depth of 10 cm and at the +10 m sampling point for an in-depth analysis of the soil column.
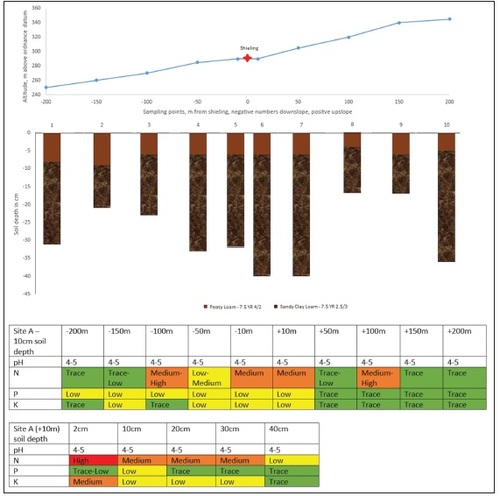
The soil has a uniform pH in the range of 4–5. Nutrient levels are clearly higher closer to the shieling. This is especially marked for N, which shows a clear decline after the sampling point 100 m at either side of the shieling. The gradient for P and K is less pronounced yet still noticeable.
The in-depth testing of the +10 m sampling point shows N concentrations highest in the topsoil and decreasing steadily with soil depth. P concentration is highest in the samples taken at 10 cm soil depth and K concentration is again highest at the top and then decreases rapidly lower down the soil profile.
Site H () similarly occupies a position underneath a steep promontory, although of lower height. The shieling sits on a south-facing slope at an altitude of 440 m, 150 m higher than site A, making it the highest shieling in Menstrie Glen. The slope below the shieling is at an angle of 15 degrees whilst the shieling itself sits on a nearly level section, again possibly indicating a created platform. Above the promontory the slope continues mainly on level ground. (The slope angle steepens beyond the +250 m sampling point.) An A-horizon of reddish peat (Munsell colour: 7.5 YR 4/4) forms the topsoil, followed by a black B1-horizon of peaty loam (Munsell colour: 10 YR 2/1). This horizon reaches the surface at the site of the actual shieling. At further depth there is a B2-horizon of reddish peaty loam (Munsell colour: 7.5 YR 4/3). At two sampling points this is followed by a Cg-horizon of gleyed clay of grey appearance with reddish mottling (Munsell colour: 7.5 YR 4/1), possibly indicating glacial drift. The soil at site H can be classed as Podzol (WRB) or Peaty Podzol according to the Scottish system.
Fig. 6. Combined slope diagram, soil column diagram and nutrient analysis report for shieling site H. The slope diagram shows the topography along the transect of site H. Soil columns visualise the stratigraphy of the soil including different soil horizons and their respective colour. Both tables visualise the results of the chemical analysis at the sampling points along the transect at a soil depth of 10 cm and at the +10 m sampling point for an in-depth analysis of the soil column.
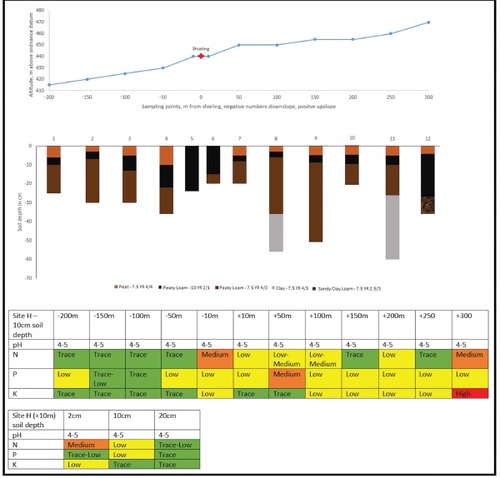
As at site A, the soil has a pH between 4 and 5. Nutrient levels are generally lower at site H than A, exhibit however again a marked increase in N close to the shieling, with levels declining rapidly immediately downslope from it and beyond the +100 m mark. P is mostly uniform at low levels throughout, K slightly enriched in an upslope direction. Between the +250 m and +300 m sampling points the angle of the slope steepens. A wire fence runs right along this break and cattle are being grazed on the land above it. Nutrient levels at the +300 m point are markedly increased, likely from this modern-day activity. The in-depth testing of the +10 m sampling point has a broadly similar pattern to that at site A. N is highest at the top and decreases with depth. P is highest at a depth of 10 cm and K decreases with depth, although more rapidly here than at site A.
DISCUSSION
vegetation dynamics at shieling sites at menstrie glen
A first question arising from the study is whether former shieling sites and the transhumance activities which took place there have influenced the vegetation in such a way as to be visible today. The study of legacies of agropastoral practices often focuses on woodland loss. Transhumance has for example been shown to have greatly contributed to the loss of tree cover in the Pyrenees (Garcia-Ruiz et al. Citation2020). From the evidence available, it is however not possible to make any informed postulations about when this process exactly happened in Menstrie Glen. Whilst shielings might well have been a kind of pioneer settlement, it is unlikely that the shielings studied here were exactly that. As post-medieval sites they came at a time when the human impact on the Scottish uplands would have already been extensive (cf. Oosthoek Citation2013), locally also shown by the fact that most of the other shielings in Menstrie Glen are potentially from an earlier time (cf. ). Palynology was used in one study aiming to reconstruct vegetational change at a different location within Menstrie Glen, reaching 300 years into the past. Evidence for oats and grasses could be found, not however for woodland (Tipping et al. Citation2001) supporting the picture of a landscape already highly modified by earlier settlement.
The present-day vegetation at both sites, whilst not uniform, shows no obviously different pattern closer to the shieling compared to further away on the transect. Whilst some small patches of J. effusus and D. cespitosa may be the result of a combination of nutrient enrichment and later preferential grazing (both are avoided by sheep) this does not present a conclusive evidence-base to develop robust theories about land-use legacies. This lack of conclusiveness stemming from the vegetational evidence furthers the case for a detailed investigation of nutrient dynamics within the soilscapes.
soil-nutrient-dynamics and land-use legacies at menstrie glen
Our primary aim was to determine whether former shieling activities left behind a legacy effect imprinted in soilscapes and soil-chemistry, reflected in the levels of plant macronutrients. and clearly show that this is indeed the case.
First, one can note that the pH at all sampling points at both locations is uniform and within the range of 4–5. Such acidic conditions reduce the mobilisation of nutrients within and from the soil. This also inhibits the process of denitrification, facilitated by bacteria, which reduces NO3 via intermediate steps to molecular N2, which is then lost to the atmosphere. The same is true for volatilisation, the process of transforming urea to NH3, which is also lost to the atmosphere (Cook Citation1983; Stevenson Citation1986). Higher pH may however have been a factor at the shieling sites whilst they were actively used. Experiments in Canada have shown that the application of cattle manure elevates soil pH levels of acidic soils (Whalen et al. Citation2000; cf. Bellows Citation2001). Cattle manure at the shieling sites may have thus accelerated N-dynamics in the past. Whilst the soil pH would revert to background levels over time, reductions or enhancements in nutrients could become imprinted in the local soilscape.
The uniform pH over both sites also makes them, notwithstanding the presence of slightly different soil- types, directly comparable to each other regarding their biogeochemical fluxes. The distribution of nutrients along the transect of site A suggests an enrichment of the soil still clearly discernible at the present day. Two features are particularly striking: the sharp drop off of enrichment beyond 100 m to both sides of the shieling site and the fact that the maxima of enrichment are not closest to the shieling but at some distance to either side of it. Our interpretation of these findings is that they are reflecting the distribution of livestock around the shieling site: As shielings were above all used for cheese and butter production, cows and ewes would have been kept in proximity to the shieling for the purpose of milking, where the milk would have had to be carried for processing. However, as the area of available land increases quadratically with the radius from the shieling, there would be insufficient grazing area available per livestock-unit within the immediate vicinity of the shieling, other than for the smallest of herds. Thus, the maxima of nutrient levels to either side may mark the main area in which the animals were kept, compared to the areas beyond the shieling into which animals would only occasionally stray (cf. Dodgshon Citation2009). At site A this boundary would have been approximately between 100 m and 150 m from the shieling, both in a downslope and upslope direction.
Another explanation for the trough in nutrient levels encircling shieling A could be overgrazing in its immediate vicinity, again brought about due to driving livestock to near the shieling for milking. That high stocking rates may lead to a local depletion of nutrients due to overgrazing has long been considered traditional knowledge among livestock farmers. It has also been shown in experiments conducted in the dry savannahs of Argentina (Abril & Bucher Citation1999) and new research in China suggest this may be due to the disruption of the normally positive effect of soil biodiversity upon N-availability where overgrazing occurs (Wang et al. Citation2020). Arbuscular mycorrhizal fungi (AMF) are some of the most important elements of this biodiversity for plant growth by forming a mutualistic symbiosis with plant roots and enhancing plant water and nutrient uptake (Douds & Millner Citation1999). Cavagnaro et al. (Citation2019) show in research conducted in Patagonia that AMF are significantly disturbed in heavily grazed landscapes when compared to moderately grazed and ungrazed areas. This is observed for plant species both preferentially grazed by livestock as well as those not normally consumed. The retention of soil nutrients is significantly influenced by the cycling occurring between plant nutrient uptake and plant dieback bringing nutrients back into the soil. If nutrient uptake is compromised through AMF-inhibition caused by overgrazing a larger proportion of nutrients will be left in the soil in forms which are more likely to be lost through water, air or direct soil erosion.
In contrast to N, P does not have a gaseous state and is not lost to the atmosphere but primarily in soluble form through leaching and in particulate form through direct erosion together with soil particles (Sharpley et al. Citation2001). One can see how both these pathways can be greatly enhanced by overgrazing. Due to reducing vegetation cover through plant removal at rates greater than the time needed for regrowth as well as trampling, which can do extensive damage at high stocking rates, overgrazing exposes the soil. Heavy rains are common in Scotland at all times of the year and cause loss of P downslope through overland and subsurface flow and eventual leaching into watercourses, groundwater or lower soil strata. Trampling also leads to a compaction of the soil, which in turn leads to a shrinkage and decrease of soil pore spaces, minimising nutrient availability for plants. Compaction through trampling in severely overgrazed pastures thus generates a feedback loop leading to more overgrazing elsewhere as livestock must move on from bare areas to find forage. In a sustainably grazed system, a high proportion of the nutrients is brought back to the soil via livestock excreta. Compaction also inhibits this process. Faecal matter deposited on a bare compact soil will be easily washed away during rain events and the nutrients it contains will consequently be lost from the area (cf. Bellows Citation2001). Evidence for such processes, such as a downslope accumulation of nutrients, could not be observed at either study site.
K is generally an understudied plant nutrient (Sardans & Peñuelas Citation2015), especially in relation to its role in grazed fields. What is known, is that K is principally lost through preferential flow and to a lesser extent matrix flow (Alfaro et al. Citation2004; Kayser & Isselstein Citation2005). Especially the former will be greatly increased where vegetation cover is decreased due to overgrazing.
The optimum distance effect, given local variations of grazing density underpinned by known soil responses to livestock grazing can explain the spatial distribution of nutrient levels at Site A. We suggest slight overgrazing immediately adjacent to the shieling despite increased manure input together with a nutrient gain through increased manuring with less grazing intensity further away from the shieling before background nutrient levels become evident. Integrating available documentary sources into the analyses reinforces the possibility of this explanation of the findings. Statutes from the Ben Lawers area from the year 1684 required shielings to be moved every five years as this ‘causes more grass to grow’ (Atkinson Citation2016). It is possible that these rules were introduced to countervail the cumulative effects of the overgrazing occurring every summer as well as the motivation of the ameliorating effect the shieling would have had on the soil through fertilization and the desire to expand the areas thus improved.
Nutrient levels at site H on the other hand portray a less clear picture. Discounting the +300 m sampling point, which has been influenced by modern-day grazing, one still sees an enhancement of nutrients in the area closer to the shieling, albeit on a lower magnitude.
This likely reflects the fewer habituations at this site indicating that it was less extensively used than site A. Furthermore, the Peaty Podzols at site H with their waterlogged A-horizon are more marginal than the Umbrisols at site A and any good grazing would have been spatially less continuous with the localised areas of enriched soil nutrients representing less wet areas than those nearby. It must be noted that vegetation on extensively waterlogged soils would have been very vulnerable to trampling by livestock. Indeed, some horizontal transhumance in coastal areas of Britain may have deliberately avoided waterlogged soils (Bradley Citation1978; Bil Citation1990, p. 38). Upland soils on slopes with only insubstantial acclivity, yet situated in areas of high rainfall, are especially vulnerable to waterlogging when underlain by impenetrable bedrock, as is clearly the case at Menstrie Glen. If primary settlement of the site entailed local deforestation, this would have increased waterlogging due to less water being retained in the biomass and reduced evapotranspiration. This would also lead to an increased rate of nutrient loss through leaching and increased soil-creep, even at very moderate angles (Vitousek Citation1983), as is evident downslope from site H.
The differences of site A and H are thus likely explained by their differences in geomorphology and hydrology leading to secondary pedogenic contrasts. Whilst site H is south facing, the much higher altitude compared to site A likely negatively affected its suitability for transhumance grazing, resulting in a more marginal and smaller shieling, in turn bequeathing less significant nutrient signatures when compared to site A. As nutrient levels are generally higher around the shieling than the background levels obtained from the outlying samples, one can suggest that overgrazing, although a possibility, did not occur at a significant scale at either site. When considering the localised spatial heterogeneity of nutrients at both sites one must also scrutinise how nutrients are cycled in grazed systems. Defecation and urination generally occur in a more concentrated way than the uptake of the plants during feeding. In short, whilst nutrients are removed from all areas of a grazed area, they are returned onto a much smaller area, leading to patches of high and low nutrient concentrations right next to each other (cf. Morton & Baird Citation1990; Whitehead Citation2000, p. 30), something which can be observed at both sites over a scale of only tens of metres.
For both sites one must also consider that enriched nutrient levels may at least in part have been preserved by a self-sustaining effect, whereby areas with higher nutrients would have had better grass and would thus have been preferentially grazed and fertilized by the sheep kept on the land of Menstrie Glen after the cessation of the shieling system (cf. Miller Citation1967). However, both sites A and H are not currently grazed in any form and it is unclear when they last were. It should further be noted that the mentioned concentrated defecation of sheep does not normally occur where grazing is highest but rather at the sites of their night encampments (cf. Arnold & Dudzinski Citation1978, p. 171), hence the self-sustaining effect might diffuse over time.
soils and vegetational sustainability of shielings in a changing environment
Our findings serve as a point of departure for a broader discussion on whether, from a soils perspective, the shieling system contributed to long-term environmental sustainability in the face of population and climate change. Whilst popular opinion often espouses the notion that traditional agropastoral practices must be inherently sustainable or by any means ‘better’ than modern-day methods, it is worth contemplating the possibility that this may not always be the case.
For our consideration of the shieling system it must be recognised that its establishment occurred over the tapestry of an already existing cultural landscape. In the Scottish uplands centuries of deforestation have created a rangeland-anthrome, where the regrowth of trees is primarily hampered by excess herbivore browsing. This situation leads to a floristically depleted landscape and a nutritionally highly modified, locally enriched, locally depleted soilscape (cf. Darling Citation1955 for one of the first comprehensive accounts of this). New work by Ellis et al. (Citation2021) highlights that traditional forms of low-intensity land use were rarely the primary drivers of environmental change. Whilst shieling systems can be classified as low intensity, the transformation of the Scottish upland forests into a rangeland-anthrome cannot. The question whether shielings were sustainable can thus only be addressed in relation to them preserving the status quo they were founded upon.
Considering the soils and vegetation at site A and H it can be proposed that these shielings were environmentally sustainable insofar as they did not deplete the soil any further than had already been the case and that they did not lead to a depletion of vegetation beyond that which must have occurred prior to the establishment of the post-medieval shielings (i.e. woodland loss). This can be inferred from the nutrients at both sites being higher closer to the shieling than at outlying sampling points. Similarly, inspection of aerial photography and investigations on the ground show the vegetation beyond the transects to be not substantially different from that along it, other than on a very localised scale for the occurrence of small patches of neutral grassland likely due to the enriching effect of the shielings.
Having established that soils at Menstrie Glen indeed show the imprint of a legacy effect of former shieling activity in the form of nutrient enrichment, one can consider the relationship this effect would have had on the suitability of shielings to become permanent farms and thus support an increase in the population due to their ‘priming’ effect on the soil. The findings of an increase of soil fertility at both sampling sites in Menstrie Glen demonstrate that the shieling system could to a certain degree be seen as an agricultural bridgehead, as a method of preparing marginal soils for permanent farming, albeit it remains unclear to what degree this was ever an intentional process. These findings are consistent with historical studies. Mather (Citation1970) points out in his study of the land use prior to 1745 in Strathfarrer that shielings there were some of the first elements of a population expanding into new lands and that those which subsequently became permanent farms in turn established their own shielings on ground of yet higher elevation. Similar trends can be observed in the Cairngorms, where shielings at a certain altitude represent an area of overlap with permanent settlements (Smith Citation1986). Sources contemporary with the shielings in Assynt speak of lush fields and of cultivation occurring on up to a third of all such sites, the enhancement of the soil through the shieling allowing limited arable farming (Miller Citation1967).
Whilst the enhanced nutrient levels found at the studied sites would have certainly worked in favour of any attempts at arable farming, there are no traces of cultivation directly at the studied sites A and H. Only sites D, E are overlain by later-date field systems (), which would have belonged to farms at lower elevation and might thus have been incidental. In Menstrie Glen the scant documentary evidence available supports the notion that the usage of shielings was beginning to decline as early as the mid-sixteenth century (RCAHMS 2001), i.e. prior to widespread agricultural expansion. Similar to what has been observed for shieling areas on Skye (MacSween Citation1959) the shielings at Menstrie Glen were never directly converted into farmsteads. These observations support the interpretation that an increase in soil nutrients as seen in Menstrie Glen did not automatically lead to an eventual transformation of shielings into farms and that other factors may have been the primary drivers for such developments elsewhere, the increased soil nutrients acting merely as a facilitator, not a main driver of agricultural expansion into areas of higher elevation.
Population developments and climatic factors may have been these primary drivers of both the waning and waxing of the shieling system and the transformation of former shielings into farms. The work by Parry (Citation1975) for example highlights the altitudinal decrease of c. 140 m of cropping for oats in the south-east Scottish Uplands, from 1300 to 1600 during the Little Ice Age (LIA). As shielings operated beyond the climatic margins of arable farming, long-term trends, especially in summer temperature, would have greatly affected the practice (cf. Parry 1976; Citation1978) and might have created an incentive for the expansion of transhumance during the LIA as other forms of land-use became unviable. Interestingly, one can find that the area of farm abandonment in the Lammermuir Hills in the Southern Uplands during the LIA (Parry Citation1976b) corresponds spatially with the existence of a number of shielings of post-medieval origin in the Canmore dataset. Elsewhere this process may have also been reverted once more as the climate was warming again. Dodgshon (Citation2005) reports of historical maps showing that two shielings near Loch Linnhe were farms in the 1580s, yet documents from 1767 refer to them as shielings having been converted into farms. It however remains to be studied to what degree these cases of exemplary evidence are typical for other sites across Scotland.
Research in Iceland has however shown that soil improvements due to manuring could have offset the impacts of the cooling climate during the LIA to allow continued farming on a subsistence level (Simpson et al. Citation2002). This might mean that drivers other than the climate were at least partly responsible for the renewed expansion of the shieling system during the LIA, such as the intensification of the cattle-trade with England (Bil Citation1990, p. 256; Cameron Citation2001). Evidence from soils, such as uncovered at Menstrie Glen, can thus help us to understand how specific agropastoral developments (such as farm establishment) took place, but it seems prohibitive to try to understand soil-nutrient levels removed from a wider context of environmental and human history and use them as narratives to reconstruct why these developments took place.
One possible explanation for the continued maintenance of shielings in Menstrie Glen is that they were the only practical option for utilising the unprepared soils at altitudes above that of the outfields with the local population pressures and climatic conditions at the time of their use. In such a scenario, site A can be seen as apposite for transhumance and would likely have been settled first. By contrast, the smaller shieling at site H on wetter soils, generally unwholesome to livestock, imply that it might have been a place of last resort, where a limited number of people conducted transhumance as all other suitable places in the glen might have already been occupied. The establishment of farms close to former shielings has rarely been clearly dated, both in Menstrie Glen and elsewhere, but is likely to fall into a period of altitudinal expansion of agricultural activity into outfields and beyond. Dodgshon (Citation1988) attributes this trend to a net population increase over the centuries from 1500–1800. Depopulating effects of LIA famines would have been obliterated after some decades. The dual socio-ecological stress of an expanding population and climate instability might have left the poorer elements of society no choice but to settle marginal areas and garner low-yield returns for high labour intensity.
Interestingly, at Menstrie Glen, despite soil nutrients in their favour, these farms were not established at the exact locations of former shieling sites. This may mean that the positive influence of the latter on the soil regarding its suitability for farming was small in comparison with the negative influence of higher elevation.
CONCLUSION
An overview on history, mechanisms and human ecology of transhumance can be developed from the existing literature; however, much remains to be studied on a global, regional and local level. Many regions of the world feature extant systems of transhumance, hitherto altogether overlooked by science. Frequently, systems have merely been examined en passant, whilst the main focus of studies was on other aspects, for example an integrated socio-economic description of a region. Although valuable insight may be obtained in this way, finer details of transhumance systems will often remain in the dark. Further focused studies on transhumance at a regional scale are needed to truly develop something akin to a world history and world theory of the practice. On a local scale the lack of knowledge is even more profound, especially when it comes to the environmental and land-use aspects in areas where transhumance has either ceased to exist or is conducted in locations which are hard to access for geographic or political reasons.
Scotland is a prime example of a region where this lack of knowledge is compounded by the challenge of having to reconstruct the practice from the evidence we can still uncover today. Thus far studies have largely focused on limited documentary evidence and archaeological work. No attention has been paid to land-use legacies. Aiming for a fuller picture, especially of the human ecology and the pressure exerted onto the land, this has been attempted here for a limited set of two shielings. As above-ground evidence in the form of vegetation surveys lacks univocal explanatory power, below-ground evidence was investigated in the form of soil-nutrient levels. Soil samples were taken along transects and analysed for N, P and K. Semi-quantitative nutrient levels were established using colorimetric test-kits. These highlighted spatially diverse nutrient enrichment in the vicinity of the shielings and allowed interpretation as to how transhumance may have been conducted at the shieling sites in Menstrie Glen and what this meant for the wider agropastoral developments in that location
primary sources
- Canmore, online catalogue of the National Record of the Historic Environment/ Historic Environment Scotland 2020. Shieling; https://canmore.org.uk/site/search/r esult?SIMPLE_KEY WORD=Shieling&SITE COUNTRY=1 [accessed 10/10/2020].
- 2020a. Enclosure (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/145355/dumyat [accessed: 29/10/2020].
- 2020b. Enclosure(S) (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/145039/dumyat [accessed: 29/10/2020].
- 2020c. Building(S) (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/145072/dumyat [accessed: 29/10/2020].
- 2020d. Bank (Earthwork)(S) (Period Unassigned), S hieling Hut(S) (Post Mediev al); https://canmore.org.uk/site/144998/loss-hill [accessed: 29/10/2020].
- 2020e. Field Boundary (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/145007/loss-hill [accessed: 29/10/2020].
- 2020f. Mound(S) (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/144981/the-balloch [accessed: 29/10/2020].
- 2020g. Bank (Earthwork) (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/144307/second-inchna-burn [accessed: 29/10/2020].
- 2020h. Enclosure (Period Unassigned), Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/144919/second-inchna-burn [accessed: 29/10/2020].
- 2020i. Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/144982/third-inchna-burn [accessed: 29/10/2020].
- 2020j. Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/145021/jerah [accessed: 29/10/2020].
- 2020k. Shieling Hut(S) (Post Medieval); https://canmore.org.uk/site/145026/crunie-burn [accessed: 29/10/2020].
Bibliography
- Abril, A., & Bucher, E. 1999. ‘The effects of overgrazing on soil microbial community and fertility in the Chaco dry savannahs of Argentina’, Applied Soil Ecol, 12 (2), pp. 159–67. doi: 10.1016/S0929-1393(98)00162-0
- Akeret, O., & Jacomet, S., 1997. ‘Analysis of plant macrofossils in goat/sheep faeces from the Neolithic lake shore settlement of Horgen Scheller – an indication of prehistoric transhumance?’, Vegetation Hist & Archaeobot, 6, pp. 235–9. doi: 10.1007/BF01370444
- Alfaro, M., Gregory, P., & Jarvis, S., 2004. ‘Dynamics of potassium leaching on a hillslope grassland soil’, J Environ Qual, 33, pp. 192–200. doi: 10.2134/jeq2004.1920
- Anderson, M., 2001. ‘Population patterns’, in The Oxford companion to Scottish History, ed. Lynch, pp. 487–91.
- Antrop, A., 2005. ‘Why landscapes of the past are important for the future’, Landscape & Urban Plann, 70, pp. 21–34. doi: 10.1016/j.landurbplan.2003.10.002
- Arnold, G., & Dudzinski, M., 1978. Ethology of Free- Ranging Domestic Animals (Amsterdam).
- Atkinson, J., 2010. ‘Settlement form and evolution in the Central Highlands of Scotland, ca. 1100–1900’, Int J Hist Archaeol, 14, pp. 316–34. doi: 10.1007/s10761-010-0110-1
- Atkinson, J., 2016. ‘Ben Lawers: an archaeological landscape in time’, Scott Archaeol Internet Rep 62.
- Battaglini, L., Bovolenta, S., Gusmeroli, F., et al., 2014. ‘Environmental sustainability of Alpine livestock farms’, Italian J Animal Sci, 13, pp. 431–43. doi: 10.4081/ijas.2014.3155
- Bellows, B., 2001. Nutrient Cycling in Pastures; http://pss.uvm.edu/pdpforage/Materials/SoilFert/nutrientcycling.pdf [accessed 26/3/2021].
- Bil, A., 1990. The Shieling 1600–1840 – the case of the central Scottish Highlands (Edinburgh).
- Birks, H., Birks, H. J., Kaland, P., & Moe, D. (eds), 1988. The Cultural Landscape – past, present and future (Cambridge).
- Bradley, R., 1978. The Prehistoric Settlement of Britain (London).
- Bradley, R., & Edmonds, M., 1988. ‘Fieldwork at Great Langdale, Cumbria 1985–1987: Preliminary Report’. Antiq J, 68, pp. 181–209. doi: 10.1017/S000358150006933X
- Brown, J., Simpson, I., Morrison, S., et al., 2012. ‘Shieling areas: historical grazing pressures and landscape responses in northern Iceland’, Human Ecol, 40, pp. 81–99. doi: 10.1007/s10745-011-9456-1
- Cameron, E., 2001. ‘Economy of the Highlands and Islands’, in The Oxford Companion to Scottish History, ed. Lynch, pp. 202–6
- Campbell, B., 2008. ‘Benchmarking medieval economic development: England, Wales, Scotand and Ireland’, Econ Hist Rev, 61 (4), pp. 896–945. doi: 10.1111/j.1468-0289.2007.00407.x
- Casado, G., & Molina, M., 2009. ‘Preindustrial agriculture versus organic agriculture: the land cost of sustainability’, Land Use Policy, 26, pp. 502–10. doi: 10.1016/j.landusepol.2008.07.004
- Cavagnaro, R., Pero, E., Dudinszky, N., et al., 2019. ‘Under pressure from above: Overgrazing decreases mycorrhizal colonization of both preferred and unpreferred grasses in the Patagonian steppe’, Fungal Ecol, 40, pp. 92–7. doi: 10.1016/j.funeco.2018.09.003
- Chambers, F., Kelly, R., & Price, S., 1988. ‘Development of the late-prehistoric cultural landscape in upland Ardudwy, North-West Wales’, in The Cultural Landscape – past, present and future, ed. Birks, Birks, Kaland & Moe (Cambridge), pp. 333–48.
- Cook, R., 1983. ‘The impact of acid deposition on the cycles of C, N, P and S’, in The Major Biogeochemical Cycles and Their Interactions. ed. B. Bolin & R. Cook (New York, US), pp. 345–64.
- Costello, E., & Svensson, E., 2018. ‘Transhumant pastoralism in historic landscapes – beginning of a European perspective’, in Historical Archaeologies of Transhumance Across Europe. ed. E. Costello &, Svensson (London), pp. 1–13.
- Darling, F. (ed.), 1955. West Highland Survey – an essay in human ecology (London).
- Davies, E., 1941. ‘The patterns of transhumance in Europe’, Geogr, 26 (4), pp. 155–68.
- Dickinson, S., 2020. ‘Bryant’s Gill: a Viking age shieling-farmstead in the English Lake District and its wider contexts’ (Pre-print, Viking Congress 2022). DOI:10.13140/RG.2.2.23635.40481.
- Dixon, P., 2007. ‘Hunting, summer grazing and settle- ment: competing land use in the uplands of Scotland’, in: Medieval Rural Settlement in Marginal Landscapes. Ruralia VII, ed. J. Klápšte & P. Sommer (Turnhout, Belgium), pp. 27–46.
- Dixon, P., 2018. ‘What do we really know about transhumance in medieval Scotland?’, in Historical Archaeologies of Transhumance across Europe, ed. E. Costello & E. Svensson (Abingdon, Oxon & New York, US), pp. 59–73.
- Dodgshon, R., 1988. ‘The ecological basis of Highland peasant farming, 1500–1800 A.D’, in: The Cultural Landscape – past, present and future, ed. Birks, Birks, Kaland & Moe (Cambridge), pp. 139–51.
- Dodgshon, R., 2005). ‘The Little Ice Age in the Scottish Highlands and Islands: documenting its human impact’, Scott Geogrl J, 12 1(4), pp. 321–37. doi: 10.1080/00369220518737243
- Dodgshon, R., 2009. ‘The environmental history of mountain regions’, in Nature’s End – history and the environment, ed. S. Sörlin & P. Warde (London), pp. 141–61.
- Dodgshon, R., & Olsson, G., 2006. ‘Heather moorland in the Scottish Highlands: the history of a cultural landscape, 1600–1880’, J Hist Geogr, 32, pp. 21–37. doi: 10.1016/j.jhg.2005.01.002
- Dodgshon, R., & Olsson, G., 2007. ‘Seasonality in European mountain areas: a study in human ecology’, in Seasonal Landscapes, ed. H. Palang, A. Printsmann & H. Soovali (Berlin), pp. 85–101.
- Douds, D., & Millner, P., 1999. ‘Biodiversity of arbuscular mycorrhizal fungi in agroecosystems’, ed. M. Paoletti (Amsterdam), pp. 77–93.
- Eddudóttir, S., Erlendsson, E., & Gísladóttir, G., 2020. ‘Landscape change in the Icelandic highland: a long- term record of the impacts of land use, climate and volcanism’, Quaternary Sci Rev, 240, article 106363. doi: 10.1016/j.quascirev.2020.106363
- Eidt, R. C., 1984. Advances in Abandoned Settlement Analysis (Milwaukee, Wisconsin, US).
- Ellis, E., Gauthier, N., Goldewijk, K., et al., 2021. ‘People have shaped most of terrestrial nature for at least 12,000 years’, PNAS: Proc Nat Acad Sci, 118 (17), e2023483118. doi: 10.1073/pnas.2023483118
- Erb, K., Fetzel, T., Kastner, T., et al., 2016. ‘Livestock grazing, the neglected land use’, in: Social Ecology – society nature relations across Time and Space, ed. H. Haberl, M. Fischer-Kowalski, F. Krausmann & V. Winiwarter (Berlin, Germany), pp. 295–313.
- Evans, E., 1940. ‘Tanshumance in Europe’, Geogr, 25 (4), pp. 172–80.
- Evans, E. 1958. ‘The Atlantic ends of Europe’, Adv of Sci, 15 (58), pp. 54–64.
- Fitter, R., Fitter, A., & Bramley, M., 1996. Wild Flowers of Britain and Northern Europe (London).
- Flinn, M., Gillespie, J., Hill, N., et al., 1977. Scottish Population History (Cambridge).
- Foster, D., Swanson, F., Aber, J., et al., 2003. ‘The importance of land-use legacies to ecology and conservation’, BioSci, 53 (1), pp. 77–88. doi: 10.1641/0006-3568(2003)053[0077:TIOLUL]2.0.CO;2
- Foster, R., 2018. ‘Norse shielings in Scotland: an interdisciplinary study of setr / sætr and ærgi- names’, Univ of Edinburgh Ph.D. thesis.
- Foster, R., 2021. ‘Reconstructing early shieling landscapes & land-use’, in Cumbria during the Viking Age’, Folk Life; 59 (1), pp. 1–17. doi: 10.1080/04308778.2021.1891729
- Fox, H., 2012, Darmoor’s Alluring Uplands (Exeter).
- Gallacher, J., Owen, J., Vaughan, A., et al., 2013. Environmental Statement – proposal for mixed woodland, Jerah Farm, Stirlingshire and Clackmannanshire; https://forestry.gov.scot/publications/202-jerah-farm-environmentalstatement/viewdocument [accessed: 20/12/2020].
- Garcia-Ruiz, J., Tomas-Faci, G., Diarte-Basco, P., et al., 2020. ‘Transhumance and long-term deforestation in the subalpine belt of the central Spanish Pyrenees: an interdisciplinary approach’, CATENA, 195, article 104744. doi: 10.1016/j.catena.2020.104744
- Geddes, D., 1983. ‘Neolithic transhumance in the Mediterranean Pyrenees’, World Archaeol, 15 (1), pp. 51–66. doi: 10.1080/00438243.1983.9979884
- Gingrich, S., Haidvogl, G., Krausmann, F., et al., 2015. ‘Providing food while sustaining soil fertility in two pre-industrial Alpine agroecosystems’, Human Ecol, 43, pp. 395–410. doi: 10.1007/s10745-015-9754-0
- Grant, J., 1977. Highland Villages (London).
- Hamilton, A., & Davies, A., 2007. ‘Written in the hill – an environmental history project in the Scottish Uplands’, Hist Scotland, May/June 2007, pp. 25–32.
- Hanna Instruments, 2018. HI-3895-010 Replacement Reagents for HI-3895 pH, Nitrogen, Phosphorus, Potassium; https://www.hannainstruments.co.uk/soil-analysisned-n-ascorbic-acid-p-tetraphenylborate-k.html#specifications [accessed: 07/01/2021].
- Hofmeister, B., 1961. ‘Wesen und Erscheinungsformen der Transhumance: Zur Diskussion um einen agrargeographischen Begriff’, Erdkunde, 15 (2), pp. 121–35. doi: 10.3112/erdkunde.1961.02.04
- Holliday, V.T., & Gartner, W. G., 2007. ‘Methods of soil P analysis in archaeology’, J Archaeol Sci, 34, pp. 301–33. doi: 10.1016/j.jas.2006.05.004
- Hubbard, C., 1984. Grasses – a guide to their structure, identification, uses and distribution in the British Isles (London).
- Huntsinger, L., Forero, L., & Sulak, A., 2010. ‘Trans- humance and pastoral resilience in the Western United States’, Pastoralism, 1 (1), pp. 9–36.
- JNCC (Joint Nature Conservation Committee), 2010. Handbook for Phase 1 Habitat survey (Peterborough).
- Kayser, M., & Isselstein, J., 2005. ‘Potassium cycling and losses in grassland systems: a review’, Grass & Forage Sci, 60 (3), pp. 213–24. doi: 10.1111/j.1365-2494.2005.00478.x
- Lynch, M. (ed.), 2001. The Oxford Companion to Scottish History (London).
- MacDonald, G., Bennett, E., & Taranu, Z., 2012. ‘The influence of time, soil characteristics, and land-use history on soil phosphorus legacies: a global meta-analysis’, Global Change Biol, 18 (6), pp. 1904–17. doi: 10.1111/j.1365-2486.2012.02653.x
- MacSween, D., 1959. ‘Transhumance in North Skye’, Scott Geogr Mag, 75(2), pp. 75–88.
- McLauchlan, K. K., 2006. ‘The nature and longevity of agricultural impacts on soil carbon and nutrients: a review’, Ecosystems, 9, pp. 1364–82. doi: 10.1007/s10021-005-0135-1
- Mather, A., 1970. ‘Pre-1745 land use and conservation in a highland glen: an example from glen Strathfarrar, north Inverness-shire’, Scott Geogr Mag, 86 (3), pp. 159–69.
- Miller, R., 1967. ‘Land use by summer shielings’, Scott Stud, 11, pp. 193–221.
- Morton, J., & Baird, D.. 1990. ‘Spatial distribution of dung patches under sheep grazing’, New Zealand J Agric Res, 33 (2), pp. 285–94. doi: 10.1080/00288233.1990.10428421
- Nedelea, A., 2009. ‘Effects of transhumance on the southern slope of the Fargas Mountains, GeoJ Tourism & Geosites, II (2,4), pp. 125–32.
- Oosthoek, J., 2013. Conquering the Highlands: a history of the afforestation of the Scottish Uplands’, World Forest History Series (Canberra, Australia).
- Parry, M., 1975. ‘Secular climatic change and marginal agriculture’, Trans Inst Br Geogr, 64, pp. 1–13. doi: 10.2307/621462
- Parry, M., 1976a. ‘The significance of the variability of summer warmth in upland Britain’, Weather, 31 (7), pp. 212–17. doi: 10.1002/j.1477-8696.1976.tb04442.x
- Parry, M., 1976b. ‘The mapping of abandoned farmland in upland Britain: an exploratory survey in south-east Scotland’, Geogr J, 142, pp. 101–10. doi: 10.2307/1796029
- Parry, M., 1978. Climatic Change, Agriculture and Settlement (Folkestone).
- Perring, M., De Frenne, P., Baeten, L., et al., 2016. ‘Global environmental change effects on ecosystems: the importance of land-use legacies’, Global Change Biol, 22, pp. 1367–1371. doi: 10.1111/gcb.13146
- Plini, R., Ravazzi, C., Raiteri, L., et al., 2017. ‘From pristine forests to high-altitude pastures: an ecological approach to prehistoric human impact on vegetation and landscapes in the western Italian Alps’, J Ecol, 105(6), pp. 1580–97. doi: 10.1111/1365-2745.12767
- RCAHMS (Royal Commission on the Ancient and Historical Monuments of Scotland), 2001. ‘Well Sheltered & Watered’ Menstrie Glen a Farming landscape near Stirling (Edinburgh).
- Reitmaier, T. (ed.), 2012. Letzte Jäger, erste Hirten. Hochalpine Archäologie in der Silvretta, Amt für Kultur, Archäologischer Dienst Graubünden (Chur, Switzerland).
- Reitmaier, T., Doppler, T., Pike, A., et al., 2018. ‘Alpine cattle management during the Bronze Age at Ramosch-Mottata, Switzerland’, Quaternary Int, 484, pp. 19–31. doi: 10.1016/j.quaint.2017.02.007
- Sardans, J., & Peñuelas, J., 2015. ‘Potassium: a neglected nutrient in global change’, Global Ecol & Biogeogr, 24, pp. 261–75. doi: 10.1111/geb.12259
- Schiavon, S., Sturaro, E., Tagliapietra, F., et al., 2019. ‘Nitrogen and phosphorus excretion on mountain farms of different dairy systems’, Agric Systems, 168, pp. 36–47. doi: 10.1016/j.agsy.2018.10.006
- Sharpley, A., McDowell, R., & Kleinman, P., 2001. ‘Phosphorus loss from land to water: integrating agricultural and environmental management’, Plant & Soil, 237, pp. 287–307. doi: 10.1023/A:1013335814593
- Shepherd, C., 2021. ‘The central uplands of Buchan – a distinctive agricultural zone in the 13th century: fact or fiction?’, Rural Hist,32 (1), pp. 41–75. doi: 10.1017/S0956793320000114
- Simpson, I. A., van Bergen, P. F., Perret, V., et al., 1999. ‘Lipid biomarkers of manuring practice in relict anthropogenic soils’, Holocene, 9 (2), pp. 223–9. doi: 10.1191/095968399666898333
- Simpson, I., Adderley, P., Gu∂mundsson, G., et al., 2002. ‘Soil limitations to agrarian land production in premodern Iceland’, Human Ecol, 30 (4), pp. 423–43. doi: 10.1023/A:1021161006022
- Smith, J., 1986. ‘Deserted farms and shealings in the Braemar area of Deeside, Grampian region, Proc Soc Antiq Scotland, 116, pp. 447–53. doi: 10.9750/PSAS.116.447.453
- Stevenson, F., 1986. Cycles of Soil (New York, US).
- Suttie, J., & Reynolds, S. (eds), 2003. Transhumant Grazing Systems in Temperate Asia (Rome, Italy).
- Swetnam, T., Allen, C., & Betancourt, J., 1999. ‘Applied historical ecology: using the past to manage the future’, Ecol Appl, 9(4), pp. 1189–206. doi: 10.1890/1051-0761(1999)009[1189:AHEUTP]2.0.CO;2
- Tipping, R., Waldron, R., & Cowley, D., 2001. ‘Pollen analysis and historic landscape change at Ashentrool, Menstrie Glen’, Forth Nat & Hist, 24, pp. 3–6.
- Vitousek, P., 1983. ‘The effects of deforestation on air, soil and water’, in The Major Biogeochemical Cycles and their Interactions, ed. B. Bolin & R. Cook (New York, US), pp. 223–46.
- Wang, L., Delgado-Baquerizo, M., Zhao, X., et al., 2020. ‘Livestock overgrazing disrupts the positive associations between soil biodiversity and nitrogen availability’, Functional Ecol, 34 (8), pp. 1713–20. doi: 10.1111/1365-2435.13575
- Whalen, J., Chang, C., Clayton, G., & Carefoot, J., 2000. ‘Cattle manure amendments can increase the pH of acid soils’, Soil Sci Soc America J, 64 (3), pp. 962–6. doi: 10.2136/sssaj2000.643962x
- Whitehead, D., 2000. Nutrient Elements in Grassland: Soil-plant-animal Relationships (Wallingford, Oxon).
- Wilkinson, B., 2009. ‘A study of turf: historic rural settlements in Scotland and Iceland’, Archit Heritage, 20, pp. 15–31. doi: 10.3366/E135075240900017X
- Winchester, A., 2000. The Harvest of the Hills – rural life in northern England and the Scottish Borders, 1400–1700 (Edinburgh).
- Zlateva, B., Dumanov, B., & Rangelov, M., 2018. ‘Applications of soil phosphate analyses for identifi- cation of activity areas at Doschkere, SE Bulgaria’, J Hist Archaeol & Anthropol Sci, 3, pp. 57–60.