

Abstract
This study investigated the role of LncRNA HOTAIR knockdown in the biological impacts on cervical cancer cells. The HOTAIR gene in two human cervical cancer cell lines was silenced with small interfering (si) RNA siHOTAIR. Proliferation, apoptosis, migration and invasion of cells were assessed following the knockdown. The expressions of Notch1, EpCAM, E-cadherin, vimentin and STAT3 were assessed using qRT-PCR and Western blotting analysis. Compared with controls, HOTAIR levels were reduced significantly, the OD values of cells were significantly decreased in proliferation assays, cell apoptosis was significantly increased, cell migration and invasion were significantly reduced after HOTAIR knockdown. Molecular analysis showed that Notch1, EpCAM, vimentin and STAT3 expressions were decreased significantly, while the expression of E-cadherin was significantly increased after HOTAIR knockdown. Rescue experiments further confirmed that Notch1 and STAT3 were involved in siHOTAIR-mediated reduction of migration and invasion of cervical cancer cells.
What is already known on this subject? Long non-coding RNAs including HOTAIR, is implicated in occurrence and development of cancer and have been explored to develop new therapeutic options for cancer.
What do the results of this study add? HOTAIR silencing significantly reduces the viability and migration ability of cells and induces cell apoptosis, adding experimental data supporting the potential use of HOTAIR specific-siRNA as a therapeutic avenue for the cancer.
What are the implications of these findings for clinical practice and/or further research? The finding from this study would help develop clinically applicable therapeutic avenues for the cancer and identify new treatment targets in the relevant pathways leading to new drugs or treatments.
IMPACT STATEMENT
Introduction
Cervical cancer is the second most frequent female malignant tumour and accounts for about 527,000 new cases of cancers and over 265,000 deaths every year worldwide (Torre et al. Citation2015, Cancer Genome Atlas Research et al. Citation2017). Despite significant advance in diagnostic techniques and therapeutic strategies for the cancer, the overall outcome of patients with cervical cancer is still poor (Kodama et al. Citation2010, Vu et al. Citation2018) and the survival time following various treatment strategies is unsatisfactory (Kodama et al. Citation2010). Therefore, developing new therapeutic strategies is essential to significantly improve therapeutic outcome for the cancer (Menderes et al. Citation2016).
Long non-coding RNA (LncRNA) is a group of endogenous RNAs having a length of more than 200 nt that do not encode proteins. Increasing evidence indicates that LncRNAs are implicated in various biological and pathological processes by up- or down-regulating the expression of target genes (Caley et al. Citation2010, Zhu and Xu, Citation2013, Cai et al. Citation2017). Therefore, LncRNAs have been investigated as potential therapeutic targets and agents in various tumours, including cervical cancer (Garcia-Gimenez et al. Citation2017, Wang et al. Citation2017). HOTAIR is a LncRNA produced based on the antisense strand of the homeobox C gene in the chromosome 12 (Rinn et al. Citation2007). This RNA is highly expressed in different cancers such as breast and colorectal cancers and is regarded as an oncogene in different tumours, including cervical cancer (Berteaux et al. Citation2005, Hajjari and Salavaty, Citation2015, Tang and Hann, Citation2018). The abnormal HOTAIR expression is associated with progression, migration, proliferation and invasion of cancers, including cervical cancer (Liu et al. Citation2016). For example, it may promote tumour progression by absorbing miR-17-5p and miR-326 in cervical cancer (Ji et al. Citation2018). Conversely, knockdown of HOTAIR suppresses the invasiveness of cancer, especially in cells with abundant PRC2 activity (Gupta et al. Citation2010).
However, whether HOTAIR can reduce the malignancy of cervical cancer cells via other pathways is largely unclear. In the present study, we studied the impact of HOTAIR knockdown on the biological behaviours of cervical cancer cells with a focus on its regulation of genes that are related to invasion and metastasis of tumour.
Materials and methods
Cell lines
Cervical cancer cell lines DoTc2 (ATCC CRL-7920) and C-33-A (ATCC HTB-31) were purchased from the American Type Collection Centre (ATTC, Rockefeller, MD, USA). Cells were cultured in RPMI-1640 medium (M0159, Creative Bioarray, Shirley, NY, USA) containing 10% foetal bovine serum (FBS) at 37 C° in a humidified air containing 5% CO2.
Reagents and instruments
FBS (FBS001-HI) was obtained from Neuromics, Edina, MN, USA; siHOTAIR for HOTAIR knockdown and scramble control (si-NC) were purchased from Beyotime Biotech, Shanghai, China; Trizon Reagent, Lipofectamine 3000 Transfection Reagent. RiboPure RNA Purification Kit, RevertAid First Strand cDNA Synthesis Kit, Dead Cell Apoptosis Kit with Annexin V FITC and PI, TaqMan Pre-Amp Master Mix, RIPA buffer, Pierce BCA protein assay kit and goat anti-rabbit IgG secondary antibody and WesternBreeze Chemiluminescent Kit were purchased from Thermo Scientific, Beijing, China; CCK-8 reagent was purchased from Sigma, St. Luis, USA; rabbit antibodies against Notch 1, EpCAM, E-cadherin, vimentin and STAT3 were from Abcam, Cambridge, UK. Inverted phase contrast microscopy (IX51) was purchased from Olympus, Kyoto, Japan.
Transfection
For HOTAIR knockdown, cells were transfected using Lipofectamine 3000 Transfection Reagents based on the supplier’s instructions. Briefly, cells were cultured to a confluency of 70-80% and transfected with siHOTAIR or si-NC (0.5 ng/mL) diluted in Opti-MEM medium using Lipofectamine 3000 diluted with MEM medium. Non-transfected cells were employed as negative control. The sequences of siHOTAIRs and si-NC are listed in . For rescue experiments, pcDNA3.1-STAT3 and pcDNA3.1-Notch1 overexpression vectors were synthesised at GenePharma (GenScript, China) and transfected into cells in various combinations with siHOTAIR using Lipofectamine 3000 Transfection Reagents based on the supplier’s instructions.
Table 1. List of small interfering RNA (siRNA) sequences used in the present study.
Cell viability assessment
The viability of cell was assessed using a CCK8 cell viability assay kit based on the supplier’s protocols. Briefly, cells were precipitated by centrifugation at 500 g at 37 °C for 10 min, resuspended in RPMI-1640 medium and cultured in the wells of 96-well plates. After cultured for 48 h, 10 µl CCK8 assay solution was added to each well and the cells were cultured at 37 °C for another 4 h. Optical density (OD) was then read at an absorbance wavelength of 450 nm with a microplate reader. The experiments were run in triplicate and the average values of readings were calculated for each treatment.
Flow cytometry
Apoptosis was assessed using the Dead Cell Apoptosis Kit according to the supplier’ instructions. Briefly, cells were seeded into the wells of 12-well plates, cultured to a confluency between 70 and 80% and pelleted by centrifugation at 500 g at 25 °C for 5 min. Cell pellets were resuspended and washed with PBS three times, digested with RNase A (100 μg/ml) at 37 °C for 30 min. 100 μl cells were labelled with Annexin V (3 μl) and propidium iodide (PI) (7 μl) at 25 °C for 30 min in the dark and loaded onto a FACScalibur flow cytometer (Becton Dickinson) for analysis.
Transwell assay
To evaluate the migration and invasion ability after siHOTAIR knockdown, Transwell assays were performed essentially as described previously (Pijuan et al. Citation2019). Briefly, 48 hours after transfection, cells were harvested as described above, washed with 1 X PBS and treated with 0.25% trypsin containing EDTA at 37 °C for 2 min. Digested cells were precipitated by centrifugation at 300 g at 25 °C for 10 min and resuspended in RPMI-1640 medium. 104 cells (100 μl) were added to the top chambers of the Transwell membrane inserts (8 μm pore diameter) and incubated at 37 °C and 5% CO2 for 24 h in serum-free medium. For invasion analysis, 10% FBS was used as a chemoattractant in the medium. After incubation, cells migrated to the lower chambers were fixed in 4% paraformaldehyde solution at 25 °C for 20 min. The cells were then dyed with 0.1% crystal violet solution at 25 °C for 15 min and photographed under a microscope. The numbers of invaded and migrated cells in each treatment were counted in five randomly selected fields for analysis.
qRT-PCR
qRT-PCR was performed to quantify the level of RNA transcripts. TRIzol reagents were used to extract total RNA from cells and the extracted RNA was reversely transcribed into cDNA using RevertAid First Strand cDNA Synthesis Kit according to the supplier’s protocols. A total of 1.5 μl of the cDNA was used in pre-amplification PCR reactions using TaqMan Pre-Amp Master Mix in a total reaction volume of 10 μl. The cycling parameters were 10 min at 95 °C followed by 18 cycles, each consisting of 15 s at 94 °C and 4 min at 62 °C. qRT-qPCR was performed on a CFX Connect PCR machine using TaqMan Gene Expression Assay mixture. GADPH was employed as an internal reference. The primers used for qRT-qPCR are presented in . Relative transcript level was determined using the 2−ΔΔCt protocol as previously described (Livak and Schmittgen, Citation2001).
Table 2. List of primer sequences used in the present study.
Western blot analysis
Cells were pelleted by centrifugation at 500 g at 37 °C for 10 min, washed twice with pre-chilled PBS and lysed in the radioimmunoprecipitation assay (RIPA) buffer to extract proteins. Protein contents in the lysates were measured with BCA protein assay kit according to the supplier’s instructions. After denatured by boiling at 100 °C for 5 min, 60 µg proteins were used in 12% polyacrylamide gel electrophoresis (SDS-PAGE), transferred to a polyvinylidene fluoride (PVDF) membranes. The PVDF membranes were then blocked by soaking in 5% non-fat milk solution prepared with 1x tris-buffered saline (TBS) containing 0.1% Tween 20 (TBST) for 4 h at 25 °C on a shaker operating at 30 rpm. After blocking, the membranes were reacted to the antibodies at 4 °C overnight. The filters were then rinsed six times at 25 °C with TBS buffer, then incubated with goat horseradish peroxidase (HRP) conjugated anti-rabbit IgG at 25 °C for 2 h. Immunoreactive bands on the blots were visualised using WesternBreeze Chemiluminescent kit in the dark according to the manufacturer’s instructions.
Statistical analysis
Statistical analyses of experimental data were performed using SPSS software (v. 19.0, SPSS Inc., Chicago, IL, USA). Data were shown as the means ± SD (standard derivation) of at least three repeats. One-way ANOVA was used to compare the difference among groups. A p-value <0.05 was considered statistically significant.
Results
siHOTAIR reduces HOTAIR expression
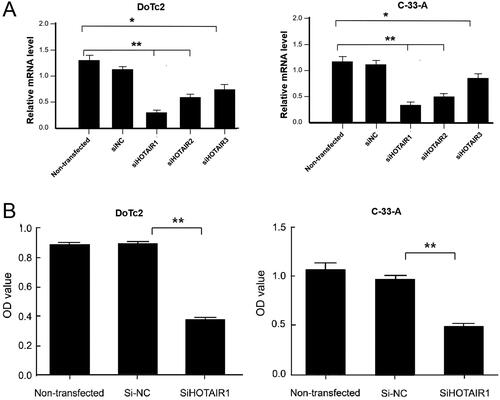
To achieve sufficient knockdown, cells from two cervical cancer cell lines DoTc2 and C-33-A were transfected with three siHOTAIRs. 48 hours after the transfection, the levels of HOTAIR transcript were determined using RT-PCR and were found significantly down-regulated as compared with the controls (non-transfected cells and cells transfected with si-NC, ). Among the three siHOTAIRs tested, shHOTAIR1 generated the greatest reduction in the HOTAIR transcript level, and was hence used in subsequent experiments. On the other hand, si-NC did not change HOTAIR1 level significantly, indicating that HOTAIR could be knockdown by siHOTAIRs specifically.
Figure 1. HOTAIR levels and cell viability determined by OD value after DoTc2 and C-33-A cells were transfected with siHOTAIRs. (A) relative mRNA levels, (B) Cell viability. Experiments were repeated three times. Comparisons were made using one-way ANOVA. * and ** p < 0.05 and < 0.01.
siHOTAIR knockdown reduces cell viability
We then determined the viability of DoTc2 and C-33-A cells 48 hours after siHOTAIR was knockdown using CCK8 assay. As shown in , compared with the controls (non-transfected cells and cells transfected with si-NC), the OD values of siHOTAIR1-transfected cells were only about 40% of the controls and the reduction was statistically significant. On other hand, the scramble control (si-NC) did not change the viability significantly () as compared with non-transfected cells, indicating that HOTAIR knockdown specifically reduces the viability and proliferation ability of DoTc2 and C-33-A cells.
siHOTAIR knockdown increases apoptosis
To ascertain the mechanisms underlying the reduced viability, we analysed cell apoptosis after siHOTAIR knockdown using flow cytometry. The data showed that there were significantly more apoptotic cells (both early and late apoptosis) in both cell lines after HOTAIR was knockdown in the cells as compared with the controls (p < 0.01, ). On the other hand, apoptotic rates were similar between si-NC and non-transfected cells.
Figure 2. Apoptosis, migration and invasion of DoTc2 and C-33-A cells after transfection using siHOTAIR1. (A) upper panel, flow cytometry results, lower panel: % of apoptosis cells. (Ba) left side: transwell assay results, right side: cell count. (Bb) migration assessment. Left side: transwell assay results, right side: cell count. Experiments were triplicated. Comparisons were made using one-way ANOVA. * and ** p < 0.05 and < 0.01.
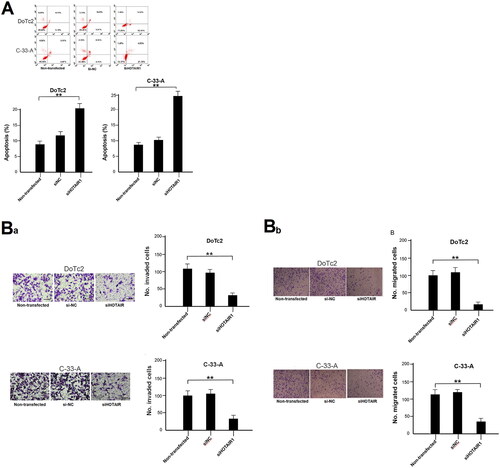
siHOTAIR knockdown reduces migration and invasion ability
To assess the changes in migration and invasion ability after HOTAIR knockdown, cells were assessed using the Transwell assays. It was found that both migration and invasion of the two cell lines were significantly reduced after HOTAIR was knockdown as compared with controls (p < 0.01, ). On the other hand, no statistically significant difference in the migration and invasion ability was observed between non-transfected and si-NC-transfected cells ().
siHOTAIR knockdown changes the expressions of genes related to invasion and metastasis
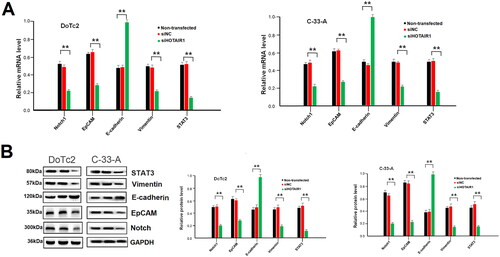
Since the invasion and metastasis ability of the cells was reduced after siHOTAIR knockdown, we analysed the expression of a few selected genes that are related to invasion and metastasis pathways. The expressions of Notch1, EpCAM, E-cadherin, vimentin, and STAT3 were detected at both mRNA transcript and protein levels. The data showed that the transcript and protein levels of these genes were significantly reduced except E-cadherin, which was significantly increased after HOTAIR was knockdown (P < 0.01, ). The expression of these genes was, however, not altered if the cells were not transfected or transfected with si-NC (.
Figure 3. Expression of invasions and metastasis-related genes in DoTc2 and C-33-A cells after HOTAIR knockdown. (A) relative mRNA levels, (B) left side: typical Western blot, right side; relative protein content. Experiments were triplicated. Comparisons were made using one-way ANOVA. * and ** p < 0.05 and < 0.01.
Notch1 and STAT3 overexpression restores migration and invasion ability
To further verify that the downregulated genes are involved in the regulation of migration and invasion, we overexpressed Notch1 and STAT3 alone or together (Supplementary Figure 1(A and B)) in the presence of siHOTAIR. Migration and invasion assay showed that either Notch1 or STAT3 alone was not able to change the reduction of migration and invasion after HOTAIR knockdown, but overexpression of both Notch1 and STAT3 resulted in partial restoration of reduced migration and invasion due to siHOTAIR (Supplementary Figure 1(C)).
Discussion
Using two cervical cancer cell lines, we found that HOTAIR silencing resulted in reduced viability, metastatic ability and increased apoptosis. The knockdown also changed the expressions of genes in metastasis pathways. These data indicate that siRNA technology may be applied to cervical cancer cells as a potential therapeutic tool and HOTAIR could be a possible target for gene therapy.
Previously, HOTAIR level was demonstrated elevated in a broad spectrum of cancers such as lung tumour (Loewen et al. Citation2014), breast tumour (Zhao et al. Citation2018), cervical cancer (Liu et al. Citation2018). HOTAIR also plays a role in the metastasis of squamous cell carcinoma in the oesophagus (Gupta et al. Citation2010, Yang et al. Citation2011, Chen et al. Citation2013). After HOTAIR knockdown with interfering RNA, the viability of DoTc2 and C-33-A cells was reduced significantly and apoptosis was increased, demonstrating that the viability of cervical cancer cells is suppressed by downregulating HOTAIR. This is consistent with results from a recent study showing that knockdown of HOTAIR reduces cell viability and enhances the apoptosis of cervical cancer cells by upregulating BCL2 expression (Liu et al. Citation2018). HOTAIR knockdown also inhibits the propagation and metastasis of breast cancer cell, because HOTAIR reduces the expression of a target miR-20a-5p (Zhao et al. Citation2018).
To investigate HOTAIR knockdown-mediated attenuation of the malignant behaviours of the cervical cancer cells, we analysed the expression level of several genes that are involved in invasion and metastasis pathways. Notch signalling pathway has been proposed as a new target for cancer therapy because Notch is aberrantly activated in many different solid tumours (Li et al. Citation2017). Notch1 was found decreased after HOTAIR knockdown in the two cell lines, suggesting that HOTAIR may affect cell viability, apoptosis, invasion and migration by downregulating Notch1. On the other hand, activating Notch was found to promote the progression of ovarian cancer (Liu et al. Citation2020).
Epithelial cell adhesion molecule (EpCAM) is known to be strongly associated with poor clinical outcomes in cancer patients (Park et al. Citation2020). It is regarded as a pro-metastatic gene and the presence of circulating cancer cells expressing EpCAM in the blood of cancer patients is strongly associated with poor prognosis (de Wit et al. Citation2018). This gene is upregulated in many tumours, such as squamous cell carcinoma (Stoecklein et al. Citation2006) and breast cancer (Osta et al. Citation2004). The expression of EpCAM also was downregulated in the cervical cell lines after HOTAIR knockdown. Previously, downregulation of EpCAM was shown to suppress the proliferation of gastric cancer and arrest cell cycle of AGS and SGC7901 cells (Wenqi et al. Citation2009). In addition, silencing of EpCAM with siRNA was found to efficiently inhibits the proliferation of MCF-7 and WERI-Rb1 cells (Subramanian et al. Citation2015). These data are consistent with our results.
E-cadherin plays pivotal role in maintaining the integrity of cell and downregulation of E-cadherin could lead to the occurrence of EMT events (Guilford, Citation1999, Wells et al. Citation2008). Our analysis showed that E-cadherin expression was upregulated in HOTAIR knockdown-cells with reduced invasion and migration ability. It is likely that the upregulation of E-cadherin may lead to mesenchymal to epithelial reverting transitions (MErT) (Chao et al. Citation2010). When E-cadherin is upregulated, the growth and metastasis of bladder cancer cells are suppressed through downregulating the β-catenin/TCF target genes (Li et al. Citation2018). Therefore, upregulation of E-cadherin is considered to be a therapeutic goal in cancer treatment (Howard et al. Citation2008).
In addition, the expression of vimentin was also assessed and was found downregulated after HOTAIR was knockdown. Vimentin protein promotes migration of cell by inputting mechanical signals into the cells from the environment and regulating the dynamic changes in the microtubules and actomyosin network according to environment stimuli (Battaglia et al. Citation2018). It is overexpressed in various epithelial cancers, including prostate cancer, gastrointestinal cancer and cervical cancer (Satelli and Li, Citation2011, Lin et al. Citation2017). Upregulation of vimentin increases the migration and invasion of human cervical cancer cells, while silencing vimentin by siRNA attenuates cell migration and invasion (Xu et al. Citation2017).
IL6/STAT3 signalling has been shown to drive metastasis in cancers by inducing EMT (Zeng et al. Citation2019). Our results showed that after HOTAIR knockdown, the expression of signal transducer and activator of transcription 3 (STAT3) is significantly downregulated. This might contribute to reduced viability, reduced invasion and migration ability of the cancer cells. Previously, it was shown that miR-411 overexpression leads to reduced STAT3 expression and inhibition of cell proliferation and invasion in cervical cancer (Shan et al. Citation2019). STAT3 is broadly upregulated both in cancer and non-cancerous cells. It inhibits the expression of key immune activators and promotes the synthesis of immunosuppressive factors. Increased STAT3 expression and activation are observed in cervical disease and may play role in cervical carcinogenesis (Morgan et al. Citation2018, Morgan and Macdonald, Citation2019a). Therefore, whether the downregulation of STAT3 expression after HOTAIR knockdown is associated with the activation of immunosuppressive factors would be an interesting topic for further study.
To further verify the involvement of genes downregulated by HOTAIR knockdown, we performed rescue experiments for Notch1 and STAT3 as examples. Our data showed that co-overexpression of Notch1 and STAT3 could partially restore the reduced migration and invasion ability of DoTc2 cells by siHOTAIR, suggesting that these two genes are likely linked to migration and invasion. Therefore, HOTAIR at least partially acts via Notch1 and STAT3 to impact the progression of cervical cancer. Although targeting STAT3 may have therapeutic potential for cancers, particularly for HPV-driven cervical cancers, the development of direct STAT3 inhibitors has been problematic. Our works suggest that HOTAIR may be a potential agent to target STAT3, in additional the protein Janus kinase 2 that has been demonstrated to effective to reduces cell proliferation and induces apoptosis in HPV containing cervical cancer cells (Morgan and Macdonald, Citation2019b).
There are limitations in our study. As a cell-based investigation, the results need to be validated with in vivo and human study. Relationship between HOTAIR and all individual genes for EpCAM, E-cadherin, and vimentin and E-cadherin has not been validated using rescue experiments or other approaches, and should be further verified. The molecular mechanisms underlying HOTAIR-mediated regulation of expression of the metastatic genes were not explored.
In conclusion, knockdown of HOTAIR by siRNA reduces cell growth, invasion, migration and induces apoptosis in cervical cancer cells. Invasion- and metastasis-related genes are downregulated except E-cadherin after HOTAIR knockdown, indicating that HOTAIR has broad impact on metastasis signalling pathways (Supplementery ). These findings enhance our understanding of the metastasis of cervical cancer and would help develop potential therapeutic approach for the cancer.
Ethical approval
Since this was a cell-based study, no ethical approval was needed based on local regulations on medical science research.
Supplemental Material
Download Zip (3.6 MB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
References
- Battaglia, R.A., et al. 2018. Vimentin on the move: new developments in cell migration. F1000Research, 7, 1796.
- Berteaux, N., et al. 2005. H19 mRNA-like noncoding RNA promotes breast cancer cell proliferation through positive control by E2F1. The Journal of Biological Chemistry, 280 (33), 29625–29636.
- Cai, L., et al. 2017. The lncRNA HNF1A-AS1 is a negative prognostic factor and promotes tumorigenesis in osteosarcoma. Journal of Cellular and Molecular Medicine, 21 (11), 2654–2662.
- Caley, D.P., et al. 2010. Long noncoding RNAs, chromatin, and development. The Scientific World Journal, 10, 90–102.
- Cancer Genome Atlas Research, et al. 2017. Integrated genomic and molecular characterization of cervical cancer. Nature, 543, 378–384.
- Chao, Y.L., Shepard, C.R. and Wells, A., 2010. Breast carcinoma cells re-express E-cadherin during mesenchymal to epithelial reverting transition. Molecular Cancer, 9 (1), 179.
- Chen, F.J., et al. 2013. Upregulation of the long non-coding RNA HOTAIR promotes esophageal squamous cell carcinoma metastasis and poor prognosis. Molecular Carcinogenesis, 52 (11), 908–915.
- de Wit, S., et al. 2018. EpCAM(high) and EpCAM(low) circulating tumor cells in metastatic prostate and breast cancer patients. Oncotarget, 9 (86), 35705–35716.
- Garcia-Gimenez, J.L., et al. 2017. Role of glutathione in the regulation of epigenetic mechanisms in disease. Free Radical Biology and Medicine, 112, 36–48.
- Guilford, P., 1999. E-cadherin downregulation in cancer: fuel on the fire? Molecular Medicine Today, 5 (4), 172–177.
- Gupta, R.A., et al. 2010. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature, 464 (7291), 1071–1076.
- Hajjari, M. and Salavaty, A., 2015. HOTAIR: an oncogenic long non-coding RNA in different cancers. Cancer Biology & Medicine, 12 (1), 1–9.
- Howard, E.W., et al. 2008. E-cadherin upregulation as a therapeutic goal in cancer treatment. Mini Reviews in Medicinal Chemistry, 8 (5), 496–518.
- Ji, F., et al. 2018. Long noncoding RNA HOTAIR: an oncogene in human cervical cancer interacting with microRNA-17-5p. Oncology Research Featuring Preclinical and Clinical Cancer Therapeutics, 26 (3), 353–361.
- Kodama, J., et al. 2010. Prognostic factors in stage IB-IIB cervical adenocarcinoma patients treated with radical hysterectomy and pelvic lymphadenectomy. Journal of Surgical Oncology, 101 (5), 413–417.
- Li, C., et al. 2018. Upregulation of Ecadherin expression mediated by a novel dsRNA suppresses the growth and metastasis of bladder cancer cells by inhibiting beta-catenin/TCF target genes. International Journal of Oncology., 52, 1815–1826.
- Li, L., et al. 2017. Notch signaling pathway networks in cancer metastasis: a new target for cancer therapy. Medical Oncology, 34 (10), 180.
- Lin, J., et al. 2017. The prognostic values of the expression of Vimentin, TP53, and Podoplanin in patients with cervical cancer. Cancer Cell International, 17 (1), 80.
- Liu, M., et al. 2018. Long non-coding RNA HOTAIR promotes cervical cancer progression through regulating BCL2 via targeting miR-143-3p. Cancer Biology & Therapy, 19 (5), 391–399.
- Liu, S., Zhang, M. and Qu, P., 2016. Expression level and clinical significance of HOX transcript antisense intergenic RNA in cervical cancer: a meta-analysis. Scientific Reports, 6, 38047.
- Liu, Z., et al. 2020. GATA1-regulated JAG1 promotes ovarian cancer progression by activating Notch signal pathway. Protoplasma, 257 (3), 901–910.
- Livak, K.J. and Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods, 25 (4), 402–408.
- Loewen, G., et al. 2014. Functions of lncRNA HOTAIR in lung cancer. Journal of Hematology & Oncology, 7, 90.
- Menderes, G., et al. 2016. Immunotherapy and targeted therapy for cervical cancer: an update. Expert Review of Anticancer Therapy, 16 (1), 83–98.
- Morgan, E.L. and Macdonald, A., 2019a. Autocrine STAT3 activation in HPV positive cervical cancer through a virus-driven Rac1-NFkappaB-IL-6 signalling axis. PLOS Pathogens, 15 (6), e1007835.
- Morgan, E.L. and Macdonald, A., 2019b. JAK2 inhibition impairs proliferation and sensitises cervical cancer cells to cisplatin-induced cell death. Cancers, 11 (12), 1934.
- Morgan, E.L., et al. 2018. STAT3 activation by E6 is essential for the differentiation-dependent HPV18 life cycle. PLoS Pathogens, 14 (4), e1006975.
- Osta, W.A., et al. 2004. EpCAM is overexpressed in breast cancer and is a potential target for breast cancer gene therapy. Cancer Research, 64 (16), 5818–5824.
- Park, D.J., et al. 2020. EpCAM-high liver cancer stem cells resist natural killer cell-mediated cytotoxicity by upregulating CEACAM1. Journal for ImmunoTherapy of Cancer, 8 (1), e000301.
- Pijuan, J., et al. 2019. In vitro cell migration, invasion, and adhesion assays: from cell imaging to data analysis. Frontiers in Cell and Developmental Biology, 7, 107.
- Rinn, J.L., et al. 2007. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell, 129 (7), 1311–1323.
- Satelli, A. and Li, S., 2011. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cellular and Molecular Life Sciences, 68 (18), 3033–3046.
- Shan, D., Shang, Y. and Hu, T., 2019. MicroRNA-411 inhibits cervical cancer progression by directly targeting STAT3. Oncology Research, 27 (3), 349–358.
- Stoecklein, N.H., et al. 2006. Ep-CAM expression in squamous cell carcinoma of the esophagus: a potential therapeutic target and prognostic marker. BMC Cancer, 6, 165.
- Subramanian, N., et al. 2015. EpCAM aptamer mediated cancer cell specific delivery of EpCAM siRNA using polymeric nanocomplex. Journal of Biomedical Science, 22, 4.
- Tang, Q. and Hann, S.S., 2018. HOTAIR: an oncogenic long non-coding rna in human cancer. Cellular Physiology and Biochemistry, 47 (3), 893–913.
- Torre, L.A., et al. 2015. Global cancer statistics, 2012. CA: a Cancer Journal for Clinicians, 65 (2), 87–108.
- Vu, M., et al. 2018. Cervical cancer worldwide. Current Problems in Cancer, 42 (5), 457–465.
- Wang, X., Yang, Y. and Huycke, M.M., 2017. Microbiome-driven carcinogenesis in colorectal cancer: Models and mechanisms. Free Radical Biology and Medicine, 105, 3–15.
- Wells, A., Yates, C. and Shepard, C.R., 2008. E-cadherin as an indicator of mesenchymal to epithelial reverting transitions during the metastatic seeding of disseminated carcinomas. Clinical & Experimental Metastasis, 25 (6), 621–628.
- Wenqi, D., et al. 2009. EpCAM is overexpressed in gastric cancer and its downregulation suppresses proliferation of gastric cancer. Journal of Cancer Research and Clinical Oncology, 135 (9), 1277–1285.
- Xu, Z., et al. 2017. URI promotes the migration and invasion of human cervical cancer cells potentially via upregulation of vimentin expression. Am J Transl Res, 9, 3037–3047.
- Yang, Z., et al. 2011. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Annals of Surgical Oncology, 18 (5), 1243–1250.
- Zeng, Y.T., et al. 2019. REX1 promotes EMT-induced cell metastasis by activating the JAK2/STAT3-signaling pathway by targeting SOCS1 in cervical cancer. Oncogene, 38 (43), 6940–6957.
- Zhao, W., et al. 2018. LncRNA HOTAIR influences cell growth, migration, invasion, and apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer Medicine, 7 (3), 842–855.
- Zhu, L. and Xu, P.C., 2013. Downregulated LncRNA-ANCR promotes osteoblast differentiation by targeting EZH2 and regulating Runx2 expression. Biochemical and Biophysical Research Communications, 432 (4), 612–617.