

Abstract
Background
In order to contribute new insights for future prevention and treatment of intrahepatic cholestasis of pregnancy (ICP), and to promote positive pregnancy outcomes, we evaluated serum Ca2+ levels and inositol 1,4,5-trisphosphate receptor (InsP3R) expression in the liver tissue of a rat ICP model.
Methods
After establishing the model by injection of oestradiol benzoate and progesterone into pregnant rats, animals were divided into normal control (n = 5) and ICP model groups (n = 5). The expression of InsP3R protein in the liver, and serum levels of Ca2+, glycocholic acid and bile acid were detected.
Results
InsP3R mRNA and protein were significantly lower in the ICP model group compared to the normal group, as determined by qPCR and immunohistochemistry, respectively. Serum enzyme-linked immunosorbent assay results revealed significantly higher levels of glycocholic acid and bile acid in the ICP model group compared to the normal group, while Ca2+ levels were significantly lower. The levers of Ca2+ were significantly and negatively correlated with the levels of glycocholic acid. The observed decrease in Ca2+ was associated with an increase in total bile acids, but there was no significant correlation.
Conclusions
Our results revealed that the expression of InsP3R and serum Ca2+ levels was significantly decreased in the liver tissue of ICP model rats. Additionally, Ca2+ levels were found to be negatively correlated with the level of glycocholic acid.
PLAIN LANGUAGE SUMMARY
This study investigated the relationship between serum Ca2+ levels, inositol 1,4,5-trisphosphate receptor (InsP3R) expression and intrahepatic cholestasis of pregnancy (ICP) in a rat model. The results indicated a significant decrease in InsP3R expression and Ca2+ in the disease group compared to the control group, alongside elevated levels of glycocholic acid and bile acid. The levels of Ca2+ exhibited a negative correlation with the levels of glycocholic acid. These findings indicated that the decrease of InsP3R expression and Ca2+ levels may be related to the pathogenesis of ICP. The study provides further insight into the treatment of this disease.
Introduction
Intrahepatic cholestasis of pregnancy (ICP), also known as obstetric cholestasis syndrome, is a disorder that occurs specifically in the second and third trimesters of pregnancy. Total bile acids and cholylglycine are sensitive biochemical markers for the diagnosis of ICP patients. The overexpression of serum glycocholic acid and high bile acid levels is involved in the occurrence of adverse pregnancy outcomes in patients with ICP (Vasavan et al. Citation2021). ICP has serious adverse effects on perinatal infants, including faecal staining of amniotic fluid, foetal growth restriction and neonatal asphyxia. Moreover, ICP increases the rate of caesarean sections, and total bile acids exceeding 100 μmol/L significantly increase the risk of stillbirth (Çelik et al. Citation2019). Although the symptoms of ICP, such as jaundice and itchy skin, disappear rapidly after delivery, they frequently recur in a second pregnancy or in patients treated with oral oestrogen contraceptives, and the incidence was increased with age and number of pregnancies (Gao et al. Citation2020, Mangla et al. Citation2020). Therefore, close monitoring of serum glycocholic acid and bile acid levels is essential for the diagnosis and management of patients with ICP. Unfortunately, the current drug treatment options for ICP are limited.
In recent years, transporter genes involved in bile formation and secretion have been identified and cloned (Garrido et al. Citation2022). Enzyme 7 α-hydroxylase (CYP7A1) is specific to the liver and is the rate limiting step in bile acid production. As a sensor, FXR participates in numerous regulatory processes involving bile acid, cholesterol and triglyceride metabolism. Bile acids activate FXR with high affinity, thereby inhibiting CYP7A1, and negative feedback reduces the production of bile acids. Exogenous oestrogen downregulates the expression of bile acid transport protein, while progesterone metabolites inhibit FXR function, resulting in increased bile acid production (Chen et al. Citation2015). Studies have confirmed that there are gene mutations or abnormal expression of genes responsible for bile secretion and transport in patients with cholestasis, including bile salt export pump (BSEP), multidrug resistance associated protein 2 (MRP2), multidrug resistance transport protein (MDR1) and multidrug resistance transport protein 3 (MDR3). BSEP and MDR3 play a key role in bile acid secretion. About 16% of patients with ICP have mutations in MDR3 that affect the secretion of phospholipids in bile. In ICP patients, elevated levels of oestrogen and progesterone decrease the protein levels of BSEP and multidrug resistance protein 2 (MRP2), both of which play a role in bile acid homeostasis (Xiao et al. Citation2021). These advances have greatly promoted research on the mechanism of cholestasis and ICP. Intracellular calcium signals, whether free or bound to calmodulin, play a crucial role in mediating fundamental cellular processes in bile duct cells, including transepithelial transport, oocyte fertilisation, gene transcription and muscle contraction. Inositol 1,4,5-trisphosphate (InsP3) serves as the primary intracellular Ca2+ mobilisation messenger, while the inositol 1,4,5-trisphosphate receptor (InsP3R) functions as the primary Ca2+ release channel in epithelial cells, and the combination of the two regulates Ca2+ channels (Rodrigues et al. Citation2018, Franca et al. Citation2019). In a study of cholestatic-type liver injury, the absence of InsP3R in cholangiocytes is linked to impaired Ca2+ signalling and Ca2+-mediated bicarbonate secretion. However, the upstream components of the Ca2+ signalling pathway remained unchanged. This finding suggests that the defect in Ca2+ signalling results from a selective reduction in InsP3R expression (Herta et al. Citation2022). Although cholestasis is the result of a combination of factors, the combined loss of Ca2+ signalling and InsP3R may be the final common pathway leading to cholestasis (Amaya and Nathanson Citation2014, Fanyin et al. Citation2018). Ursodeoxycholic acid, a hydrophilic bile acid and the only effective treatment for certain types of chronic cholestatic liver disease, enhances liver function by stimulating bile Ca2+-dependent exocytosis (Lei et al. Citation2016, Mayo et al. Citation2018, Zhou et al. Citation2019, Fu and Xu Citation2021). Therefore, stimulation of intracellular Ca2+ signalling may be an effective strategy for the treatment of cholestatic diseases.
Recent research has found that the inhibition of IP3R3/p-IP3R3 levels in patients with ICP might affect the transportation of Ca2+ from the endoplasmic reticulum to the mitochondria, weakening the stress adaptation associated with mitochondria-associated membranes, which lead to adverse foetal outcomes in ICP (Li et al. Citation2023). Based on many similarities between ICP and the pathogenesis of cholestasis, it is speculated that Ca2+ signalling may also be impaired in hepatocytes of patients with ICP. Indeed, the disruption of Ca2+ signalling leads to abnormal functioning of intrahepatic bile, ultimately causing cholestasis. Abnormal Ca2+ signalling and InsP3R expression or function may represent the final common pathway for the pathogenesis of ICP. Using a rat ICP model, our aim was to clarify abnormalities of Ca2+ signalling and InsP3R in ICP, and to confirm that the alteration of this signalling is the final pathway for ICP pathogenesis. Our findings provide new insights for the effective treatment of ICP in the future, which should lead to a reduction in perinatal mortality and the incidence of caesarean sections.
Materials and methods
Experimental animals
Female and male Sprague Dawley rats (specific pathogen-free grade), aged 10 weeks old were purchased from Hunan Slaccas Jingda Laboratory Animal (Changsha, China) under licence number SCXK (Xiang) 2019-0004. No randomisation or blinding was done.
Ethics statement
The experimental procedures were performed in accordance with the principles of the Guide for the Ethical Use of Animals in Applied Etiology Research and the licences for experimental animals (licence number SCXK (Xiang) 2019-0004), and the experimental procedures were approved by the Ethics Committee of Taizhou Municipal Hospital of China (protocol number LWPJ106) (Li et al. Citation2017). Furthermore, we confirmed that all methods were reported in accordance with the ARRIVE guidelines (Kilkenny et al. Citation2010).
Reagents and instruments
Oestradiol benzoate (50-50-0) was purchased from Beijing Solarbio Technology (Beijing, China). Progesterone (57-83-0) was purchased from Shanghai Ye Yuan Biotechnology (Shanghai, China). Corn oil (GB/T19111) was provided by Qinglong Hi-Tech Oils & Fats Industry (Yichun, China). The Trizon reagent (cw0580s), Ultrapure RNA Ultrapure RNA Extraction Kit (CW0581M) and DAB colour reagent kit (CW0125) were purchased from Kangwei Century (Dongtai, China). The HiScript II Q RT SuperMix for qPCR (+gDNA wiper) (R223-01) was purchased from Vazyme Biotech (Nanjing, China). 2 × SYBR Green PCR Master Mix (A4004M) was purchased from Lifeint Technology (Xiamen, China). Haematoxylin Stain Solution (ZLI-9610) and horseradish-enzyme-labelled goat anti-rabbit IgG (H + L) (ZB-2301) were purchased from Zhongshan Jinqiao Biotechnology Group (Beijing, China). InsP3R (19962-1-AP) was purchased from BioLion Technology (Singapore). Centrifugation was accomplished with a D1008E microcentrifuge (SciLogex, Rocky Hill, CT). The dry electric heater (GL-150) was purchased from Haimen City Qilin Bell Instrument Manufacturing (Nantong, China). The low-temperature high-speed centrifuge (5424R) was a product of Eppendorf (Hamburg, Germany). The fluorescence PCR instrument (CFX Connect™) was a product of Bó Lè Life Medical Products (Shanghai, China). The automatic sample rapid grinder (Tiss-12) was a product of Shanghai Jingxin Industrial Development (Shanghai, China). The electric heating constant temperature incubator (DHP-9054) was a product of Shandong Broke Biological Industry (Shandong, China). A CX41 microscope (Olympus, Tokyo, Japan) and slicer (2235, Leica, Wetzlar, Germany) were used. The electric-heating blast-drying oven (DHG-9070A) was a product of Heng Science Instrument (Beijing, China).
Animal model construction and drug administration
Experimental animals were divided into normal control and ICP model groups, with five pairs of animals, respectively. Oestradiol benzoate solution was prepared by dissolving 12.5 mg of oestradiol benzoate in 100 mL of corn oil to make a 0.125 mg/mL solution. Progesterone solution was prepared by dissolving 750 mg of progesterone in 100 mL of corn oil to make a 7.5 mg/mL solution. Each pair of female and male rats was caged at a ratio of 1:1, and female rats exhibiting a vaginal plug on the same day were recorded as pregnancy 1 d. On the 15th day of pregnancy, each ICP model group was given a 10 mL/kg dose of benzoic acid. Glycol solution and progesterone solution were continuously administered by intragastric administration for 5 d, and then rats were sacrificed with isoflurane.
Real-time fluorescent quantitative PCR
Rat liver samples were collected for mRNA extraction. cDNA was synthesised with a HiScript II Q RT SuperMix for qPCR (+gDNA wiper) reverse transcription kit according to the manufacturer’s instructions. Briefly, cDNA was used as the template, the sample was loaded with fluorescent dyes, and the program was set on the fluorescence quantitative PCR instrument to carry out the amplification reaction. According to the relative quantitative 2−△△Ct method, the relative mRNA expression of VEGF5 and sflt-1 was calculated with β-actin as the internal reference. The primers used were: InsP3R-F, CTCTCCGGCACCCAAAGAAGAG, InsP3R-R, AGCACATCTCCTACTCCGCC; β-actin-F, GCCATGTACGTAGCCATCCA, β-actin-R, GAACCGCTCATTGCCGATAG.
Immunohistochemical detection
Liver tissue was collected from the rats. Tissue sections were placed in a 65 °C oven for 2 h, followed by xylene for 10 min, which was replaced with xylene for another 10 min, Subsequently, sections were placed for 5 min each in 100% ethanol, 95% ethanol and 80% ethanol in purified water. For antigen retrieval, sections were placed in a repair box with antigen retrieval solution, heated in a pressure cooker, and allowed to naturally cool for 2 min on the heat source. Then, they were moved into a wet box with freshly prepared 3% hydrogen peroxide to remove the endogenous peroxidase blocking solution for 10 min, and rinsed with PBS. Next, the slides were soaked three times in PBS (5 min each), the PBS around the tissue was blotted with absorbent paper, 5% bovine serum albumin was added dropwise onto the slides for blocking at 37 °C for 30 min. After absorbing the blocking solution around the tissue with absorbent paper, a sufficient amount of diluted primary TH antibody (1:200) was dropped onto each slide, which was put in a humid box and incubated at 4 °C overnight. For secondary antibody staining the following day, the wet box was allowed to stand at room temperature for 45 min, the slides were soaked in PBS three times (5 min each), and goat anti-rabbit antibody (1:100) was added for incubation at 37 °C for 30 min. After washing the slides with PBS, colour development and counterstaining were performed with DAB for 10 min. The degree of staining was checked under a microscope before rinsing slides with PBS for 1 min, counterstaining with haematoxylin for 3 min, differentiating with hydrochloric acid alcohol, and returning to blue. Finally, slides were rinsed with tap water for 1 min, dehydrated, cleared, covered and examined by microscopy.
Serum ELISA detection
Pregnant rats were sacrificed after anaesthesia with isoflurane, and 1.5 mL of whole blood was collected by encapsulating the eyes. After whole-blood samples were collected, they were allowed to stand at room temperature for 30 min, centrifuged at 3000 rpm for 15 min, and about 0.5 mL of serum was taken for ELISA detection.
Statistical analysis
SPSS 20.0 software (SPSS Inc., Chicago, IL) was used for statistical analysis. All experiments were repeated three times. Quantitative results are expressed as the mean and standard deviation (χ ± s). One-way ANOVA was used for comparison of the two groups, the Student–Newman–Keuls method was used for pairwise comparisons, and Pearson’s correlation analysis was used to assess for linear relationships between the two groups. The test level was α = 0.05 and p < .05 indicated a significant difference. No data were excluded from the analysis, and the investigators were not blinded in any animal experiments throughout the study.
Ethics statement
The experimental procedures were performed in accordance with the principles of the Guide for the Ethical Use of Animals in Applied Etiology Research and the licences for experimental animals (licence number SCXK (Xiang) 2019-0004), and the experimental procedures were approved by the Ethics Committee of Taizhou Municipal Hospital of China (protocol number LWPJ106) (Li et al. Citation2017). Furthermore, we confirmed that all methods were reported in accordance with the ARRIVE guidelines (Kilkenny et al. Citation2010).
Results
InsP3R mRNA and protein expression were reduced in ICP rats
We designed and carried out experiments according to the flowchart (). The qPCR curve of InsP3R mRNA in the normal group was basically consistent, and the same was the model group. The relative expression level of InsP3R mRNA in the normal group was about twice that in the ICP model group. It was found that InsP3R mRNA expression in the model group was significantly lower than that in the normal group (, p < .05).
Figure 1. Flowchart for the study (Established the ICP and the normal model. Collected liver and serum. Analysed the expression of InsP3R in liver of the model. Analysed the levels of serum Ca2+, glycocholic acid and bile acid in serum of the model. Analysed the correlation between the level of Ca2+ and the levels of glycocholic acid and bile acid.).
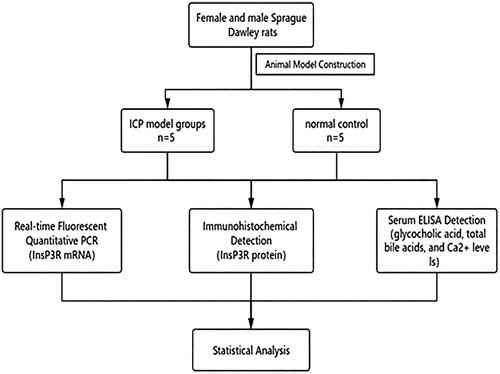
Figure 2. qPCR analysis of InsP3R mRNA expression. (A) Results of PCR amplification. (B) InsP3R mRNA relative expression of control group and model group. *InsP3R mRNA expression in the model group was significantly lower compared with the control/normal group (p < .05).
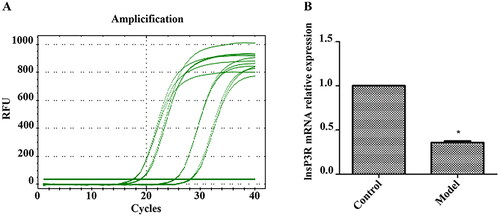
There was more positive staining for InsP3R protein in the rat liver tissues from the normal group compared to the ICP model group. According to statistics, the integrated optical density/area (IOD/area) level of the InsP3R protein in the normal group was significantly higher than that in the model group. These results demonstrated that the expression of InsP3R protein in the model group was significantly lower than that in the normal group (, p < .05).
Figure 3. Immunohistochemistry analysis of InsP3R protein expression. (A) Images of immunohistochemistry. (B) Relative expression of lnsP3R protein in each group. *Expression of InsP3R in the model group was significantly lower compared with the control/normal group (p < .05).
Correlations between glycocholic acid, total bile acids and Ca2+ levels
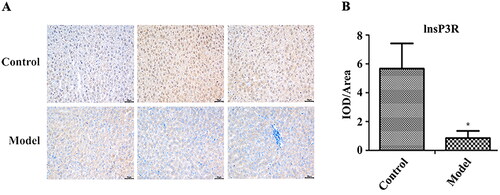
The serum Ca2+, glycyrrhetinic acid and total bile acids levels of pregnant rats were detected using a fully automated biochemical analyser. Levels of total bile acids and glycocholic acid in the model group were significantly higher compared to the normal group (, p < .05). Ca2+ levels were negatively correlated with glycocholic acid levels (p < .05). The decrease of Ca2+ was related to the increase of total bile acids, but there was no significant correlation ().
Figure 4. Correlation between glycocholic acid, total bile acids and calcium ion. CG: glycocholic acid; TBA: total bile acids.
Table 1. Comparison of ELISA indexes (x ± s).
Discussion
ICP is a common complication during pregnancy whose pathogenesis is mainly caused by cholestasis in central capillaries of the liver (Fu and Xu Citation2021). A meta-analysis found that both total bile acids and glycocholic acids were effective markers for the diagnosis of ICP, and glycocholic acids had a higher diagnostic power than total bile acids. Compared to healthy pregnant women, levels of alanine aminotransferase, aspartate aminotransferase, total bile acids and glycocholic acid in pregnant women with ICP were abnormally increased (p < .05), indicating that ICP was closely related to bile acids and positively correlated with glycolic acid levels. This study found that levels of glycocholic acid and bile acids in the ICP model were higher than those in the normal group, consistent with previous results (Piechota and Jelski Citation2020). The total bile acids are closely related to perinatal outcomes. Severe ICP is defined as total bile acids ≥40 μmol/L, which may lead to steatorrhoea and reduce the absorption of fat-soluble vitamin K, resulting in postpartum haemorrhage. In addition, it may cause intrauterine foetal death, foetal cardiac dysfunction (Vasavan et al. Citation2021), neonatal intracranial haemorrhage and other risks.
There are three types of InsP3R: type 1 (InsP3R1), type 2 (InsP3R2) and type 3 (InsP3R3) (Amroudie and Ataei Citation2016, Iyer and Franzini-Armstrong Citation2021, Yang et al. Citation2021). Previous studies have found that the loss of InsP3R2 in hepatocytes and loss of InsP3R3 in cholangiocytes are both important for the pathogenesis of cholestasis. InsP3R3 expression was decreased in cholangiocytes of patients with biliary diseases such as cirrhotic cholangitis (Franca et al. Citation2019, Rodrigues et al. Citation2022). The major epithelial cells of the liver are hepatocytes and cholangiocytes, and InsP3R is the only intracellular Ca2+ release channel for these cells (Zhang et al. Citation2021). To open Ca2+ channels, four InsP3 molecules must bind to the InsP3R (Alzayady et al. Citation2016). Furthermore, Ca2+ directly modulates the open probability of this channel. InsP3R1 releases Ca2+ at lower Ca2+ concentrations, while higher Ca2+ concentrations inhibit the channel. In contrast, the open probability of InsP3R3 increases with increasing Ca2+ concentration (Hagar and Ehrlich Citation2000). Hepatocytes express InsP3R1 and InsP3R2, while all three isoforms are expressed in cholangiocytes (Zhang et al. Citation2021).
Ursodeoxycholic acid elevated the concentration of cytoplasmic free calcium ions in bile duct cells and induced membrane binding of calcium-dependent conventional protein kinase Cα (Alpini et al. Citation2002), a mechanism similar to that observed in hepatocytes (Beuers et al. Citation1998). Ursodeoxycholic acid may stimulate the secretion of bicarbonate anions in bile duct cells through calcium-dependent mechanisms, activate calcium-dependent chloride ion channels, and stimulate chloride/bicarbonate exchange through AE2. In addition, ursodeoxycholic acid conjugate may indirectly increase the concentration of cytoplasmic free calcium ions in bile duct cells by stimulating ATP secretion into bile, which may then induce calcium-dependent chloride anion secretion via apical purinergic ATP receptors (Dranoff et al. Citation2001, Nathanson et al. Citation2001). The levels of calcium and magnesium ATPase on the erythrocyte membrane in ICP patients increased with strong activity (Zhu et al. Citation2019). Meanwhile, Kremer et al. found that patients with ICP and primary biliary cirrhosis (PBC) had an increased concentration of cytoplasmic free calcium in nerve cells (Kremer et al. Citation2010). These studies have demonstrated that the accumulation of cholic acid can enhance the uptake of Ca2+ by cells in the body, thereby reversing the decrease in plasma calcium ion concentration.
Calcium signalling in hepatocytes begins with agonist binding to G protein-coupled receptors or receptor tyrosine kinases. After agonist receptor binding, phospholipase C was activated, leading to the breakdown of the membrane phosphatidylinositol 4,5-bisphosphate to generate diacylglycerol and InsP3. InsP3 subsequently diffuses into the cytoplasm, where it binds to InsP3Rs located in the endoplasmic reticulum membrane, nuclear membrane or nucleoplasmic reticulum. InsP3–InsP3R binding results in a conformational change in InsP3R, leading to the release of internal Ca2+ stores (Hagar and Ehrlich Citation2000, Hodeify et al. Citation2018). InsP3 is inactivated upon conversion to inositol 1,2-bisphosphate by type I inositol polyphosphate phosphatase, or via InsP3-kinase- mediated phosphorylation to form inositol 1,3,4,5-tetraphosphate (Miranda et al. Citation2019, Marongiu et al. Citation2021). InsP3R2 led to impaired Ca2+ signalling and inhibited the insertion of multidrug resistance-related protein 2 into the apical plasma membrane of hepatocytes, affecting the protein’s ability to convert organic anions for secretion into bile.
In cholangiocytes, InsP3R3 accounts for approximately 80% of all InsP3Rs. InsP3R1 and InsP3R2 are responsible for the release of bicarbonate following activation of muscarinic receptor type 3 by acetylcholine. Ca2+ acts directly on apical Ca2+-dependent chloride channels, recently identified as TMEM16A channels in both small and large cholangiocytes, allowing the efflux of chloride, which is then similarly exchanged for bicarbonate via the chloride–bicarbonate exchanger (Dutta et al. Citation2020). InsP3R3 is mainly localised in the apical region of cholangiocytes (Rodrigues et al. Citation2018, Zhang et al. Citation2021). Activation of InsP3R3 leads to bicarbonate secretion through a cAMP-dependent cascade, in which activation of secretin receptors indirectly stimulates InsP3 production and Ca2+ release through InsP3R3 (Roos et al. Citation2021). cAMP production leads to apical, cystic fibrosis transmembrane conductance regulator (CFTR)-dependent release of ATP into the bile, which activates apical P2Y receptors to induce InsP3-mediated Ca2+ release through apical type III InsP3Rs. Ca2+ then drives apical Ca2+-dependent chloride channel activation and subsequent bicarbonate secretion (Minagawa et al. Citation2007). Downregulation of InsP3R3 selectively interferes with cAMP-induced bicarbonate secretion; in this model, calcium signalling was impaired or absent (Alzayady et al. Citation2016). The results of the present study show that InsP3R mRNA and protein expression levels in the ICP model group were significantly lower than those in the normal group. Therefore, we speculated that the mechanism of cholestasis in patients with ICP is similar to that of intrahepatic cholestasis caused by other diseases. The Ca2+ levels in the ICP model group were lower compared to the normal group, and significantly and negatively correlated with the glycocholic acid level (p < .05). This obvious correlation was considered to be the result of an insufficient sample size. Thus, the correlation between Ca2+ and bile acids needs further confirmation in a larger number of samples. Bile duct cells contain oestrogen receptors, and the oestrogen level in ICP patients is significantly increased, which could lead to the activation of the nuclear transcription factor erythroid 2-like 2 (NRF2) in bile duct cells. A previous study identified a musculo-aponeurotic fibrosarcoma recognition element in the InsP3R3 promoter that directly binds to NRF2 in cholangiocytes (Ueasilamongkol et al. Citation2020). Increased NRF2 binding at this site led to chromatin remodelling, which reduced promoter activity to inhibit InsP3R3 expression in cholangiocytes, resulting in reduced Ca2+ signalling and bile duct secretion.
Normal bile acid flux can stimulate the influx of Ca2+ through store-operated calcium channels (SOC) (Bird and Putney Citation2018, Gupta et al. Citation2020). Some scholars believe that cholestatic bile acids inhibit the influx of Ca2+, leading to the disruption of intracellular Ca2+ homeostasis and signalling in the downstream region of SOC (Bird and Putney Citation2018, Bonus et al. Citation2021). Other studies have shown that the administration of ursodeoxycholic acid during the early stages of ICP or other liver diseases could significantly reduce bilirubin, alanine aminotransferase and alkaline phosphatase contents to treat cholestasis (Krupa et al. Citation2021). Collectively, these studies show that choleretic bile acids can increase the concentration of free Ca2+ ions in the cytoplasm by depleting Ca2+ in the intracellular pool, which induces the translocation of matrix interacting molecules (such as STIM1) to generate SOC in the plasma membrane. Opening of SOC produces inflow of extracellular Ca2+, restoring Ca2+ in the intracellular pool. This subsequently constricts the bile ducts, enhances the excretion of bile acids, reduces the accumulation of bile acids, and thus reduces the effect of accumulated bile acids on bile ducts and hepatocyte damage. Therefore, by increasing the concentration of serum Ca2+ or artificially controlling the influx of Ca2+, choleretic drugs might promote Ca2+ influx through SOC to increase the concentration of intracytoplasmic Ca2+, thereby enhancing the excretion of bile acids and achieving treatment of patients with cholestasis (including ICP). However, this speculation still needs further experiments to confirm.
Conclusions
Serum Ca2+ levels and InsP3R expression in liver tissue were significantly decreased in the rat ICP model, and the Ca2+ level was found to be negatively correlated with the level of glycocholic acid. These findings solve a fundamental problem of current ICP treatment and provide new insights for future treatment of ICP. Indeed, our results have the potential to markedly reduce both perinatal mortality and caesarean sections, which is of great significance and has a far-reaching impact.
Serum Ca2+ levels and InsP3R expression in liver tissue were significantly decreased in the rat ICP model, and the Ca2+ level was found to be negatively correlated with the level of glycocholic acid. These findings provide new insight for future treatment of ICP.
Author contributions
D.P. and M.J. made substantial contributions to acquisition, analysis and interpretation of the data, and wrote the first draft of the manuscript; M.J., G.T., J.S., Z.S., R.C. and D.W. made substantial contributions to acquisition of data. D.P. and D.W. contributed to experimental conception and design, and analysis and interpretation of data, and drafted the manuscript and revised it critically. All authors read and approved the final manuscript.
Acknowledgements
We thank Liwen Bianji (Edanz) (www.liwenbianji.cn/) for basic language editing of a draft of this manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
All generated raw data and/or analysed data from the current study are available from the corresponding author on reasonable request.
Additional information
Funding
References
- Alpini, G., et al., 2002. Ursodeoxycholate and tauroursodeoxycholate inhibit cholangiocyte growth and secretion of BDL rats through activation of PKC alpha. Hepatology, 35 (5), 1–9.
- Alzayady, K.J., et al., 2016. Defining the stoichiometry of inositol 1,4,5-trisphosphate binding required to initiate Ca2+ release. Science Signaling, 9 (422), ra35.
- Amaya, M.J. and Nathanson, M.H., 2014. Calcium signaling and the secretory activity of bile duct epithelia. Cell Calcium, 55 (6), 317–324.
- Amroudie, M.N. and Ataei, F., 2016. Experimental and theoretical study of IBC domain from human IP3R2; molecular cloning, bacterial expression and protein purification. International Journal of Biological Macromolecules, 124, 1321–1327.
- Beuers, U., Boyer, J.L., and Paumgartner, G., 1998. Ursodeoxycholic acid in cholestasis: potential mechanisms of action and therapeutic applications. Hepatology, 28 (6), 1449–1453.
- Bird, G.S. and Putney, J.W., Jr. 2018. Pharmacology of store-operated calcium entry channels. In: Calcium entry channels in non-excitable cells. CRC Press/Taylor & Francis, Chapter 16.
- Bonus, M., Häussinger, D., and Gohlke, H., 2021. Liver cell hydration and integrin signaling. Biological Chemistry, 402 (9), 1033–1045.
- Çelik, S., et al., 2019. Predictors of adverse perinatal outcomes in intrahepatic cholestasis of pregnancy. Ginekologia Polska, 90 (4), 217–222.
- Chen, Y., et al., 2015. Estrogen and estrogen receptor-α-mediated transrepression of bile salt export pump. Molecular Endocrinology, 29 (4), 613–626.
- Dranoff, J.A., et al., 2001. Polarized expression and function of P2Y ATP receptors in rat bile duct epithelia. American Journal of Physiology. Gastrointestinal and Liver Physiology, 281 (4), G1059–G1067.
- Dutta, A.K., et al., 2020. Signaling through the interleukin-4 and interleukin-13 receptor complexes regulates cholangiocyte TMEM16A expression and biliary secretion. American Journal of Physiology. Gastrointestinal and Liver Physiology, 318 (4), G763–G771.
- Fanyin, M., et al., 2018. Ursodeoxycholate inhibits mast cell activation and reverses biliary injury and fibrosis in Mdr2−/− mice and human primary sclerosing cholangitis. Laboratory Investigation, 98, 1465–1477.
- Franca, A., et al., 2019. Effects of endotoxin on type 3 inositol 1,4,5-trisphosphate receptor in human cholangiocytes. Hepatology, 69 (2), 817–830.
- Fu, C. and Xu, Y., 2021. Value of serum glycocholic acid and total bile acids in predicting maternal and perinatal outcomes in intrahepatic cholestasis of pregnancy. Journal of Healthcare Engineering, 2021, 3911998.
- Gao, X.X., et al., 2020. Prevalence and risk factors of intrahepatic cholestasis of pregnancy in a Chinese population. Scientific Reports, 10 (1), 16307.
- Garrido, A., et al., 2022. Histone acetylation of bile acid transporter genes plays a critical role in cirrhosis. Journal of Hepatology, 76 (4), 850–861.
- Gupta, K., et al., 2020. Bile canaliculi contract autonomously by releasing calcium into hepatocytes via mechanosensitive calcium channel. Biomaterials, 259, 120283.
- Hagar, R.E. and Ehrlich, B.E., 2000. Regulation of the type III InsP(3) receptor by InsP(3) and ATP. Biophysical Journal, 79 (1), 271–278.
- Herta, T., et al., 2022. Role of the IgG4-related cholangitis autoantigen annexin A11 in cholangiocyte protection. Journal of Hepatology, 76 (2), 319–331.
- Hodeify, R., et al., 2018. Regulation and role of store-operated Ca2+ entry in cellular proliferation. In: J.A. Kozak and J.W. Putney, Jr., eds. Calcium entry channels in non-excitable cells. Boca Raton (FL): CRC Press/Taylor & Francis, 215–240.
- Iyer, R. and Franzini-Armstrong, C., 2021. The location of InsP3 receptors in Purkinje cells of murine cerebellum does not supports a direct interaction in the transfer of calcium ions between ER and mitochondria. European Journal of Translational Myology, 31 (3), 9935.
- Kilkenny, C., et al., 2010. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLoS Biology, 8 (6), e1000412.
- Kremer, A.E., et al., 2010. Lysophosphatidic acid is a potential mediator of cholestatic pruritus. Gastroenterology, 139 (3), 1008–1018, 1018.e1.
- Krupa, Ł., et al., 2021. Importance of bile composition for diagnosis of biliary obstructions. Molecules, 26 (23), 7279.
- Lei, Z., et al., 2016. Effects of taurochenodeoxycholic acid on Ca2+/CaM signalling mediated by the TGR5 signalling pathway. Die Pharmazie, 71 (7), 390–393.
- Li, Y., et al., 2017. Bile acids and intestinal microbiota in autoimmune cholestatic liver diseases. Autoimmunity Reviews, 16 (9), 885–896.
- Li, Y., et al., 2023. The spatial expression of mTORC2–AKT–IP3R signal pathway in mitochondrial combination of endoplasmic reticulum of maternal fetal interface trophoblast in intrahepatic cholestasis of pregnancy. Journal of Perinatal Medicine, 51 (8), 1032–1039.
- Mangla, A., Guerra, M.T., and Nathanson, M.H., 2020. Type 3 inositol 1,4,5-trisphosphate receptor: a calcium channel for all seasons. Cell Calcium, 85, 102132.
- Marongiu, L., et al., 2021. Inositol 1,4,5-trisphosphate 3-kinase B promotes Ca2+ mobilization and the inflammatory activity of dendritic cells. Science Signaling, 14 (676), eaaz2120.
- Mayo, M.J., et al., 2018. NGM282 for treatment of patients with primary biliary cholangitis: a multicenter, randomized, double-blind, placebo-controlled trial. Hepatology Communications, 2 (9), 1037–1050.
- Minagawa, N., et al., 2007. Cyclic AMP regulates bicarbonate secretion in cholangiocytes through release of ATP into bile. Gastroenterology, 133 (5), 1592–1602.
- Miranda, M.C.D., et al., 2019. Epidermal growth factor (EGF) triggers nuclear calcium signaling through the intranuclear phospholipase C delta-4 (PLCδ4). Journal of Biological Chemistry, 294 (45), 16650–16662.
- Nathanson, M.H., et al., 2001. Stimulation of ATP secretion in the liver by therapeutic bile acids. Biochemical Journal, 358 (Pt 1), 1–5.
- Piechota, J. and Jelski, W., 2020. Intrahepatic cholestasis in pregnancy: review of the literature. Journal of Clinical Medicine, 9 (5), 1361.
- Rodrigues, M.A., et al., 2022. Inositol 1,4,5-trisphosphate receptor type 3 (ITPR3) is overexpressed in cholangiocarcinoma and its expression correlates with S100 calcium-binding protein A4 (S100A4). Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie, 145, 112403.
- Rodrigues, M.A., Gomes, D.A., and Nathanson, M.H., 2018. Calcium signaling in cholangiocytes: methods, mechanisms, and effects. International Journal of Molecular Sciences, 19 (12), 3913.
- Roos, F.J.M., et al., 2021. Impact of hypoxia and AMPK on CFTR-mediated bicarbonate secretion in human cholangiocyte organoids. American Journal of Physiology. Gastrointestinal and Liver Physiology, 320 (5), G741–G752.
- Ueasilamongkol, P., et al., 2020. Type 3 inositol 1,4,5-trisphosphate receptor is increased and enhances malignant properties in cholangiocarcinoma. Hepatology, 71 (2), 583–599.
- Vasavan, T., et al., 2021. Fetal cardiac dysfunction in intrahepatic cholestasis of pregnancy is associated with elevated serum bile acid concentrations. Journal of Hepatology, 74 (5), 1087–1096.
- Xiao, J., et al., 2021. Molecular pathogenesis of intrahepatic cholestasis of pregnancy. Canadian Journal of Gastroenterology & Hepatology, 2021, 6679322.
- Yang, F., et al., 2021. Preservation of endoplasmic reticulum (ER) Ca2+ stores by deletion of inositol-1,4,5-trisphosphate receptor type 1 promotes ER retrotranslocation, proteostasis, and protein outer segment localization in cyclic nucleotide-gated channel-deficient cone photoreceptors. FASEB Journal, 35 (6), e21579.
- Zhang, L., et al., 2021. Upregulated SOCC and IP3R calcium channels and subsequent elevated cytoplasmic calcium signaling promote nonalcoholic fatty liver disease by inhibiting autophagy. Molecular and Cellular Biochemistry, 476 (8), 3163–3175.
- Zhou, Q., et al., 2019. Effect of 4-phenylbutyric acid and tauroursodeoxycholic acid on magnesium and calcium metabolism in streptozocin-induced type 1 diabetic mice. Biological Trace Element Research, 189 (2), 501–510.
- Zhu, P., et al., 2019. Correlation of lipid peroxidation and ATP enzyme on erythrocyte membrane with fetal distress in the uterus in patients with intrahepatic cholestasis of pregnancy. European Review for Medical and Pharmacological Sciences, 23, 2318–2324.