

Abstract
Background
Endometrial cancer is a kind of gynaecological cancer. S100A2 is a newfound biomarker to diagnose endometrial cancer. This study was to investigate the role of S100A2 on regulating migration and invasion of endometrial cancer.
Methods
The mRNA and protein levels of S100A2 were obtained by quantitative real-time polymerase chain reaction, immunohistochemistry and western blot methods. Cell viability was measured by the Cell Counting Kit-8 assay. Cell migration and invasion were quantified using transwell assays. Western blot assay was conducted to quantify protein expressions of epithelial to mesenchymal transition-related proteins (N-cadherin and E-cadherin). Furthermore, in vivo tumour formation experiments were performed to evaluate the role of S100A2 on tumour xenografts.
Results
S100A2 was significantly up-regulated in endometrial cancer tissues. Knockdown of S100A2 inhibited cell viability, migration and invasion of endometrial cancer cells. Meanwhile, STING pathway was activated by the inhibited S100A2. STING inhibitor C-176 significantly reversed the effects of S100A2 knockdown on aggressive behaviours of endometrial cancer cells. Inhibition of S100A2 dramatically suppresses the tumour growth in vivo.
Conclusions
S100A2 functions as an oncogene in endometrial cancer. Targeting S100A2 may be a promising therapeutic method to treat endometrial carcinoma.
Plain Language Summary
This study was to investigate the role of S100A2 on regulating migration and invasion of endometrial cancer. S100A2 was significantly up-regulated in endometrial cancer tissues. Knockdown of S100A2 inhibited cell viability, migration and invasion of endometrial cancer cells. Meanwhile, STING pathway was activated by the inhibited S100A2. STING inhibitor C-176 significantly reversed the effects of S100A2 knockdown on aggressive behaviours of endometrial cancer cells. Inhibition of S100A2 dramatically suppresses the tumour growth in vivo. S100A2 functions as an oncogene in endometrial cancer. Targeting S100A2 may be a promising therapeutic method to treat endometrial carcinoma.
Keywords:
Introduction
Endometrial cancer is the most common malignancy of the female reproductive tract. Women diagnosed with advanced or recurrent endometrial cancer have a poorer prognosis, with a low 5-year survival rate (Urick and Bell Citation2019). Currently, the efficacy of radiation therapy for advanced endometrial cancer remains controversial, and there is no standard treatment when the disease progresses or recurs after initial treatment (de Boer et al. Citation2019). The recurrence and progression of endometrial cancer are also associated with chemotherapy resistance (Tossetta and Marzioni Citation2023). Therefore, a better understanding of the molecular characteristics of endometrial cancer is needed to improve patient outcomes.
As previously reported, aberrant expressed genes in tissues of endometrial cancer compared with that in the normal tissues were found to be linked to the diagnosis, prognosis, and the migration and invasion in vitro. For instance, CUL4A expression was highly expressed in tissues of endometrial cancer, and CUL4A promoted proliferation, invasion and migration of endometrial cancer cells (Wang and Chen Citation2021). Highly expressed SF3B1 also functioned as an oncogene in the tumorigenesis of endometrial cancer (Popli et al. Citation2020). Additionally, inhibition of UBE2C suppressed proliferation, migration, invasion and epithelial–mesenchymal transition (EMT) of endometrial cancer cells (Liu et al. Citation2020). Recently, S100A2 has been reported to act as an oncogene in pancreatic cancer (Chen et al. Citation2021), colorectal cancer (Li et al. Citation2020) and non-small cell lung cancer (Bulk et al. Citation2009), but function as a tumour suppressor gene in gastric cancer (ZHAO et al. Citation2013), renal cell carcinoma (Sugiyama et al. Citation2020) and breast cancer (Zheng et al. Citation2021, Sugino and Sawada Citation2022). Zhang et al. suggested that S100A2 was highly expressed in endometrial cancer, and was linked to poor prognosis (Zhang et al. Citation2022). However, whether S100A2 regulates the migration and invasion of endometrial cancer cells remains unclear.
In the present study, high S100A2 expression in tissues of endometrial cancer was verified in the collected clinical samples, and was linked to sensitive diagnosis and poor prognosis. Moreover, the regulatory effects of S100A2 on cell viability, migration and invasion were investigated, and the potential pathway was studied as well. Our study aimed to provide a new therapeutic method for endometrial cancer treatment.
Methods
Antibodies and plasmids
Antibodies of S100A2, E-cadherin, N-cadherin, cGAS, STING, phospho-STING (p-STING), Notch1, HES1, PTEN, AKT, p-AKT, β-catenin, GAPDH and horseradish peroxidase conjugated goat anti-rabbit IgG polyclonal secondary antibody were all purchased from Thermo Fisher (Waltham, MA). The detail of antibodies used in this study is listed in . The oligonucleotides targeting S100A2 (sh-S100A2) and the negative control (sh-NC) were synthesised, annealed and cloned into the shRNA expression vector pRNAT-H1.1/Neo (GenScript, Nanjing, China). The sequences of all plasmids are listed in .
Table 1. Information of the antibodies.
Table 2. Information of the plasmids.
Endometrial cancer tissue collection
Forty-six pairs of endometrial tumour tissues and adjacent normal tissues were gained from the Second Affiliated Hospital of Xuzhou Medical University after the study was approved by the Ethics Committee of the Second Affiliated Hospital of Xuzhou Medical University ([2022]052105). All collected samples were frozen in liquid nitrogen for subsequent studies. The correlation between mRNA levels of S100A2 in tissues and clinicopathological parameters of all patients is summarised in .
Table 3. Correlation between S100A2 expression and EC clinicopathological parameters.
RNA isolation and quantitative real-time polymerase chain reaction (qRT-PCR)
First, RNA was extracted from tissues and cells using the Trizol reagent (Solarbio, Beijing, China). The cDNA was synthesised using a cDNA Reverse Transcription kit (Yeasen, Shanghai, China). Quantitative PCR was performed using PowerUp™ SYBR™ Green Master Mix (A25742, Applied Biosystems, Waltham, MA) in StepOnePlus™ (Applied Biosystems, Waltham, MA) according to the manufacturer’s protocols. GAPDH acted as the endogenous control, and the 2−ΔΔCT method was for transcript expression level measurement. The primer sequences are listed in .
Immunohistochemistry
The analysis of S100A2 protein expression in tissues was performed by immunohistochemistry. Tissues were deparaffinised and rehydrated. The sections were stained with rabbit anti-S100A2 antibody. Goat anti-rabbit IgG conjugated with horseradish peroxidase was used as the secondary antibody. The representative images were acquired using the Olympus model BX51 microscope (Tokyo, Japan) and were analysed digitally using Image J software (Bethesda, MD).
Western blot
Total protein was isolated lysis buffer (pH 7.4) (Dixit et al. Citation2023). Soluble proteins were collected after centrifugation at 13,000 rpm for 5 min. Protein was quantitatively determined with Bradford reagent (B6916, Sigma, St. Louis, MO), and electrophoretically transferred onto polyvinylidene difluoride membranes (Thermo Fisher, Waltham, MA). The membranes were sealed in 5% skim milk and crossbred with primary antibodies, and incubated overnight at 4 °C. The protein levels were evaluated using ECL fluorescence reagent (RPN2106, GE HealthCare, Chicago, IL). The assay was repeated three times independently.
Cell culture, treatment and transfection
The human endometrial cancer cell lines HEC-1 and RL95-2 were both purchased from the ATCC (Manassas, VA). Both of the two cell lines were cultured in DMEM/F12 (Sigma, St. Louis, MO) supplemented with 10% foetal bovine serum (FBS, Invitrogen, Carlsbad, CA).
To establish HEC-1 and RL95-2 cells with stably silenced S100A2, lentivirus vectors carrying the interference sequence were added to 2 mL complete medium supplemented with a final concentration of 5 μg/mL polybrene. Twenty-four hours later, the medium was replaced with fresh complete medium without lentivirus and polybrene for another 48 h culture. Total RNA was collected to confirm the transfection efficiency using qRT-PCR.
In some experiments, cells were treated with the STING inhibitor C-176 (1 μM, S6575, Selleck, Houston, TX) for 48 h.
Assessment of cell viability
Cell viability was evaluated by Cell Counting Kit-8 (CCK-8) assay. HEC-1 and RL95-2 cells were seeded into 96-well plates (5000 cells/well). The 10 μL CCK-8 reagent was added to each well and incubated with cells for 1–2 h. The absorbance was measured at 450 nm by a spectrometer (Shimadzu, Kyoto, Japan).
Assessment of cell migration and invasion
The Matrigel was diluted with serum-free medium 1:8 to the final concentration (5 mg/mL; 356234, BD Biosciences, Franklin Lakes, NJ) in the Transwell chamber for the invasion experiment. 5 × 105 cells (200 mL) cultured in serum-free medium were placed in the upper chamber at 37 °C for 2 h, 500 mL medium with 10% FBS was added in the lower chamber. After incubating for 24 h, the cells in the upper chamber were wiped, while those invading to the bottom were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. For each group, five random sights were imaged and then counted using Image J software (Bethesda, MD). The assay to detect cell migration followed the same procedure as the assay to detect cell invasion, except that the chamber was coated with Matrigel.
In vivo tumour xenografts
Female BALB/c nude mice (6-week-old) were purchased from Vital River Laboratories (Beijing, China). All operations of mice were approved by the Animal Ethics Committee of the Second Affiliated Hospital of Xuzhou Medical University. HEC-1 cells with stably transfected with sh-S100A2 (4 × 106 cells) were injected subcutaneously into the back of mice, and then the mice were fed for 28 days. The tumour size was measured weekly and mice were sacrificed. Afterwards, tumour weight was accessed. The tumour volume was calculated by the formula: a × b2 (a: the longest diameter of the tumour, b: the length of the diameter perpendicular to a).
Statistical analysis
The experimental data were analysed by GraphPad Prism software version 8.3 (GraphPad, La Jolla, CA), and the data operation between the two groups was represented by mean ± SD and compared with t-test. One-way ANOVA was used to compare the mean of multiple groups. Tukey’s post hoc test was used to compare pairwise comparisons between groups. The diagnostic value was determined by receiver operating characteristic (ROC) curve analysis. The Kaplan–Meier curve was plotted for the overall survival, and the log-rank test was conducted accordingly. p < .05 means the difference is statistically significant.
Results
High expression of S100A2 is associated with poor prognosis of endometrial cancer
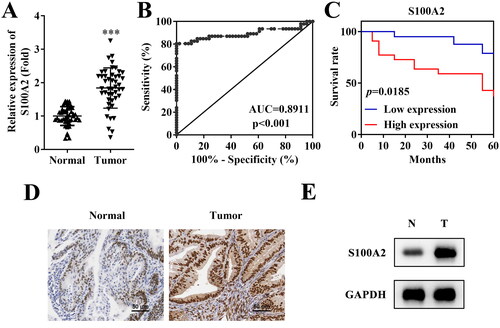
According to the PCR results, S100A2 mRNA levels were significantly upregulated in endometrial cancer tissues compared with the adjacent normal tissues (). The ROC analysis of S100A2 was performed, and the results suggested that the area under the ROC curve (AUC) of S100A2 in tumour tissues was 0.8911 (p < .001, ), indicating the potential diagnostic value of S100A2 in endometrial cancer. High expression of S100A2 was associated with the poor prognosis of endometrial cancer (). Moreover, the protein level of S100A2 was also found to be elevated in tissues of endometrial cancer according to the immunohistochemistry () and western blot () methods.
Figure 1. High expression of S100A2 is associated with poor prognosis of endometrial cancer. (A) PCR analysis of S100A2 mRNA expression in tissues of endometrial cancer and adjacent normal tissues, ***p < .001, n = 46. (B) The receiver operating characteristic (ROC) curve of S100A2 in endometrial cancer, AUC = 0.8911, p < .001. (C) Kaplan–Meier’s survival curve for endometrial cancer patients according to S100A2 expression in tumour lesions, p = .0185. The protein expression of S100A2 accessed by (D) immunohistochemistry method and (E) western blot, n = 3.
Inhibition of S100A2 suppresses aggressive behaviours of endometrial cancer cells
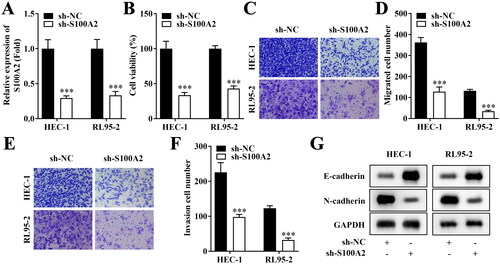
Then, the role of S100A2 on cellular functions of HEC-1 and RL95-2 cells was investigated. S100A2 was significantly down-regulated on mRNA level (). Inhibition of S100A2 prominently suppressed cell viability (), migration () and invasion (). Meanwhile, protein level of E-cadherin was upregulated and N-cadherin was down-regulated by silenced S100A2 ().
Figure 2. Inhibition of S100A2 suppresses aggressive behaviours of endometrial cancer cells. (A) mRNA levels of S100A2 after transfection in HEC-1 and RL95-2 cells, n = 3, ***p < .001. (B) Cell viability evaluated by CCK-8 assay, n = 3, ***p < .001. (C–F) Migrate and invasive HEC-1 and RL95-2 accessed by Transwell method, n = 3, ***p < .001. (G) Western blot analysis of E-cadherin and N-cadherin protein expression in HEC-1 and RL95-2, n = 3.
S100A2-mediated cGAS/STING pathway promoted the progress of endometrial cancer in vitro
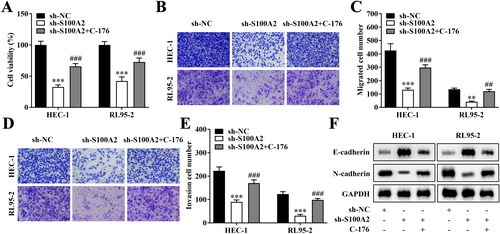
Subsequently, key protein levels of several pathways that have been reported to regulate endometrial cancer progress were evaluated under the S100A2 interference treatment. As indicated in , after S100A2 was down-regulated, cGAS/STING pathway was significantly activated in both HEC-1 and RL95-2 cells (). However, the activation of Notch1/HES1, PTEN/AKT and beta-catenin pathways was not significant in both HEC-1 and RL95-2 cells (). Afterwards, the cell viability, migration and invasion that were significantly promoted by the S100A2 knockdown were prominently inhibited by the addition of the STING inhibitor C-176 ().
Figure 3. The representative photos of the key protein of cGAS/STING, Notch1/HES1, PTEN/AKT and beta-catenin pathways obtained by western blot, n = 3.
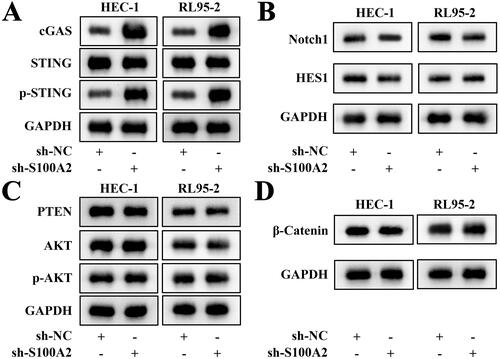
Figure 4. Inactivation of cGAS/STING pathway reversed the effects of S100A2 inhibition on endometrial cancer cells. (A) Cell viability evaluated by CCK-8 assay, n = 3, ***p < .001 (vs. sh-NC), ###p < .001 (vs. sh-S100A2). (B–E) Migrate and invasive HEC-1 and RL95-2 accessed by Transwell method, n = 3, **p < .01, ***p < .001 (vs. sh-NC), ##p < .01, ###p < .001 (vs. sh-S100A2). (F) Western Blot analysis of E-cadherin and N-cadherin protein expression in HEC-1 and RL95-2, n = 3.
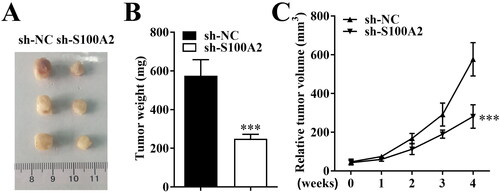
Knockdown of S100A2 suppressed tumour growth in vivo
In vivo tumour formation experiments demonstrated that knockdown of S100A2 inhibited the growth of HEC-1-derived tumours in nude mice compared with that in the negative control group. The tumour size and tumour weight of sh-S100A2 group were significantly lower than that in sh-NC group ().
Figure 5. Knockdown of S100A2 suppressed tumour growth in vivo. (A) The photographs, (B) tumour weight and (C) tumour volume of xenograft tumours from sh-NC and sh-S100A2 groups, n = 3, ***p < .001.
Discussion
Endometrial cancer is a gynaecological malignancy which usually occurs in perimenopausal and postmenopausal women, and lack of biological markers for detection (Zhou et al. Citation2021). S100A2 is a small molecule acidic protein with EF-hand calcium-binding structure, which is abnormally expressed during the occurrence and development of various tumours (Wang et al. Citation2021, Sugino and Sawada Citation2022). It has been reported that S100A2 participated in various physiological and pathological effects on human diseases (Sugino and Sawada Citation2022). Abnormal expression of S100A2 has been found in various tumours, and its abnormal expression is related to cell differentiation, protein phosphorylation, enzyme activity and other functions (Lapi et al. Citation2006, Naz et al. Citation2014). For instance, high expression level of S100A2 predicts poor outcomes in brain cancers (Yin et al. Citation2021), lung cancers (Wang et al. Citation2017) and liver cancers (Zhang et al. Citation2021). However, in kidney cancer (Dixit et al. Citation2023) and breast cancer (Wang et al. Citation2021), low expression level of S100A2 may be expected to be advantageous for tumour development. Therefore, S100A2 may be a double-edged sword in regulating tumour progression.
Moreover, studies have shown that the release of S100A2 is related to the prognosis of endometrial cancer (Zhang et al. Citation2022). Therefore, we wondered whether S100A2 regulates the aggressiveness of endometrial cancer cells. In this study, knockdown of S100A2 inhibited the tumour progress of endometrial cancer by suppressing the cell viability, migration, invasion and EMT. Previous studies have pointed out that S100A2 has an inhibitory effect on tumours mainly because S100A2 can bind Ca2+, resulting in a decrease in Ca2+ concentration, which inhibits cell proliferation and plays a role in inhibiting tumour occurrence (Kyriazanos et al. Citation2002, Doi et al. Citation2021). cGAS/STING pathway has been found to be activated in endometrial cancer and promotes Ca2+ release (Zeng et al. Citation2022). Notch1/HES1 pathway is suppressed by miR-134 to inhibit tumour progress of endometrial cancer (Gao et al. Citation2015), and inhibition of Notch1 pathway affects Ca2+ release in hepatocellular carcinoma (Si et al. Citation2023). Activated PTEN/AKT pathway in endometrial cancer (Kong et al. Citation2021), can be suppressed by elevated intracellular Ca2+ in human glioblastoma cells (Shin et al. Citation2006). Moreover, inhibition of beta-catenin pathway in endometrial cancer contributes to suppress tumour growth (Conza et al. Citation2021), and the non-canonical Wnt/Ca2+ pathway interacts with Notch signalling (Krishnamurthy and Kurzrock Citation2018). Therefore, we hypothesised that S100A2 may regulate the endometrial cancer cells through the four pathways. Our data suggested that cGAS/STING pathway was significantly activated in endometrial cancer tissues. Therefore, S100A2-mediated STING pathway prominently promoted the process of endometrial cancer.
Conclusions
In conclusion, this study demonstrated that down-regulated S100A2 might inhibit the cell viability, migration, invasion and EMT of endometrial cancer cells via cGAS/STING signalling pathway. This axis provides novel insights into the pathogenesis of endometrial cancer and S100A2 may serve as a new promising therapeutic target.
Ethical approval
The study was approved by the Ethics Committee of the Second Affiliated Hospital of Xuzhou Medical University.
Consent form
Written informed consent was obtained from all patients. All experiments were performed in accordance with relevant guidelines and regulations.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Additional information
Funding
References
- Bulk, E., et al., 2009. S100A2 induces metastasis in non-small cell lung cancer. Clinical Cancer Research, 15 (1), 1–8.
- Chen, Y., et al., 2021. S100A2 is a prognostic biomarker involved in immune infiltration and predict immunotherapy response in pancreatic cancer. Frontiers in Immunology, 12, 758004.
- Conza, D., et al., 2021. Metformin dysregulates the unfolded protein response and the WNT/beta-catenin pathway in endometrial cancer cells through an AMPK-independent mechanism. Cells, 10 (5), 1067.
- de Boer, S.M., et al., 2019. Adjuvant chemoradiotherapy versus radiotherapy alone in women with high-risk endometrial cancer (PORTEC-3): patterns of recurrence and post-hoc survival analysis of a randomised phase 3 trial. The Lancet. Oncology, 20 (9), 1273–1285.
- Dixit, G., et al., 2023. FGFR2 mutations promote endometrial cancer progression through dual engagement of EGFR and Notch signalling pathways. Clinical and Translational Medicine, 13 (5), e1223.
- Doi, S., et al., 2021. Regulation of the tubulin polymerization-promoting protein by Ca2+/S100 proteins. Cell Calcium, 96, 102404.
- Gao, Y., Liu, T., and Huang, Y., 2015. MicroRNA-134 suppresses endometrial cancer stem cells by targeting POGLUT1 and Notch pathway proteins. FEBS Letters, 589 (2), 207–214.
- Kong, C., et al., 2021. Downregulation of HOXA11 enhances endometrial cancer malignancy and cisplatin resistance via activating PTEN/AKT signaling pathway. Clinical & Translational Oncology, 23 (7), 1334–1341.
- Krishnamurthy, N. and Kurzrock, R., 2018. Targeting the Wnt/beta-catenin pathway in cancer: update on effectors and inhibitors. Cancer Treatment Reviews, 62, 50–60.
- Kyriazanos, I.D., et al., 2002. Expression and prognostic significance of S100A2 protein in squamous cell carcinoma of the esophagus. Oncology Reports, 9 (3), 503–510.
- Lapi, E., et al., 2006. S100A2 gene is a direct transcriptional target of p53 homologues during keratinocyte differentiation. Oncogene, 25 (26), 3628–3637.
- Li, C., et al., 2020. S100A2 promotes glycolysis and proliferation via GLUT1 regulation in colorectal cancer. FASEB Journal, 34 (10), 13333–13344.
- Liu, Y., et al., 2020. UBE2C is upregulated by estrogen and promotes epithelial–mesenchymal transition via p53 in endometrial cancer. Molecular Cancer Research, 18 (2), 204–215.
- Naz, S., et al., 2014. Protumorigenic actions of S100A2 involve regulation of PI3/Akt signaling and functional interaction with Smad3. Carcinogenesis, 35 (1), 14–23.
- Popli, P., et al., 2020. Splicing factor SF3B1 promotes endometrial cancer progression via regulating KSR2 RNA maturation. Cell Death & Disease, 11 (10), 842.
- Shin, S.Y., et al., 2006. Clozapine, a neuroleptic agent, inhibits Akt by counteracting Ca2+/calmodulin in PTEN-negative U-87MG human glioblastoma cells. Cellular Signalling, 18 (11), 1876–1886.
- Si, Y., et al., 2023. Phellodendronoside A exerts anticancer effects depending on inducing apoptosis through ROS/Nrf2/Notch pathway and modulating metabolite profiles in hepatocellular carcinoma. Journal of Hepatocellular Carcinoma, 10, 935–948.
- Sugino, H. and Sawada, Y., 2022. Influence of S100A2 in human diseases. Diagnostics, 12 (7), 1756.
- Sugiyama, T., Ozono, S., and Miyake, H., 2020. Expression profile of S100A2 and its clinicopathological significance in renal cell carcinoma. Anticancer Research, 40 (11), 6337–6343.
- Tossetta, G. and Marzioni, D., 2023. Targeting the NRF2/KEAP1 pathway in cervical and endometrial cancers. European Journal of Pharmacology, 941, 175503.
- Urick, M.E. and Bell, D.W., 2019. Clinical actionability of molecular targets in endometrial cancer. Nature Reviews Cancer, 19 (9), 510–521.
- Wang, H., et al., 2021. miR-325-3p promotes the proliferation, invasion, and EMT of breast cancer cells by directly targeting S100A2. Oncology Research, 28 (7), 731–744.
- Wang, T., Du, G., and Wang, D., 2021. The S100 protein family in lung cancer. Clinica Chimica Acta, 520, 67–70.
- Wang, T., et al., 2017. Expression and clinicopathological significance of S100 calcium binding protein A2 in lung cancer patients of Chinese Han ethnicity. Clinica Chimica Acta, 464, 118–122.
- Wang, X. and Chen, T., 2021. CUL4A regulates endometrial cancer cell proliferation, invasion and migration by interacting with CSN6. Molecular Medicine Reports, 23 (1), 23.
- Yin, A., et al., 2021. Integrative analysis identifies an immune-relevant epigenetic signature for prognostication of non-G-CIMP glioblastomas. Oncoimmunology, 10 (1), 1902071.
- Zeng, X., et al., 2022. IL6 induces mtDNA leakage to affect the immune escape of endometrial carcinoma via cGAS-STING. Journal of Immunology Research, 2022, 3815853.
- Zhang, C., et al., 2021. S100 family members: potential therapeutic target in patients with hepatocellular carcinoma: a STROBE study. Medicine, 100 (3), e24135.
- Zhang, Q., et al., 2022. High expression of S100A2 predicts poor prognosis in patients with endometrial carcinoma. BMC Cancer, 22 (1), 77.
- Zhao, Y., Zhang, T., and Wang, Q., 2013. Clinical significance of altered S100A2 expression in gastric cancer. Oncology Reports, 29 (4), 1556–1562.
- Zheng, S., et al., 2021. Comprehensive analysis of the prognosis and correlations with immune infiltration of S100 protein family members in hepatocellular carcinoma. Frontiers in Genetics, 12, 648156.
- Zhou, L., et al., 2021. Plasma-derived exosomal miR-15a-5p as a promising diagnostic biomarker for early detection of endometrial carcinoma. Molecular Cancer, 20 (1), 57.