

Abstract
Backgroud
Neurological disorders are common in preterm (PT) born individuals. Diffusion tensor imaging (DTI) studies using tract-based spatial statistics (TBSS) effectively detect microstructural white matter (WM) abnormalities in the brain. We conducted this systematic review to integrate the findings of TBSS studies to determine the most consistent WM alterations in PT born individuals.
Methods
PubMed, Embase, Web of Science and Science Direct were searched. DTI studies using TBSS in PT born individuals were screened up to October 2022. The systematic review included studies reporting alterations in FA values for the entire brain in a stereotactic space, with three coordinates (x, y, z), according to the seed-based d mapping method.
Results
The search strategy identified seventeen studies that fulfilled our inclusion criteria, with a total of 911 PT-born individuals and 563 matched controls were analysed. Of the seventeen studies, eight were dedicated to 650 adults, five to 411 children and four to 413 infants. Ten studies recruited 812 individuals born very prematurely (GA <29 weeks), six studies recruited 386 moderately premature individuals (GA = 29–32 weeks) and one study recruited 276 individuals born late prematurely (GA >32 weeks). This meta-analysis of six studies including 388 individuals highlighted four brain regions in which fractional anisotropy (FA) was lower in PT group than in people born at term. The quantitative meta-analysis found that the most robust WM alterations were located in the corpus callosum (CC), the bilateral thalamus and the left superior longitudinal fasciculus (SLF) II. Significant changes in FA reflect WM abnormalities in PT born individuals from infant to young adulthood.
Conclusions
Significant changes in FA reflect WM abnormalities in individuals born PT from infancy to young adulthood. The abnormal development of the CC, bilateral thalamus and left SLF may play a vital role in the neurodevelopment of PT individuals.
PLAIN LANGUAGE SUMMARY
Neurological disorders are prevalent in preterm (PT) born individuals. The use of tract-based spatial statistics (TBSS) in diffusion tensor imaging (DTI) studies has proven effective in detecting microstructural abnormalities of the white matter (WM) of the brain. In order to determine the most consistent alterations in WM among those born prematurely, we have screened DTI studies using TBSS in this PT born population up until October 2022. The meta-analysis identified four brain regions where fractional anisotropy (FA) was lower in the PT group than in those born at term. The quantitative meta-analysis identified the corpus callosum, the bilateral thalamus and the left superior longitudinal fasciculus II. As the most robust WM alterations. Various studies have demonstrated the links between PT birth, intelligence quotient, gestational age and subject age.
Introduction
About 15 million infants are born preterm (PT) (defined as < 37 weeks’ gestational age [GA]) each year, and this incidence is increasing). As survival rates increase, PT birth has become an important public health issue (Allotey et al. Citation2018). Most of previous studies on PT born individuals have highlighted the increased neurological risks associated with prematurity (Saigal and Doyle Citation2008). These individuals are at risk for adverse neurodevelopmental outcomes, including deficits in language (Aeby et al. Citation2013), attention (Bora et al. Citation2014), executive function, working memory (Menegaux et al. Citation2017) and poor educational and social outcomes (Anderson Citation2014). Neurological disorders such as attentional deficit/hyperactivity disorder (ADHD) (McNally et al. Citation2010) and autism spectrum disorders (ASD) (Solomon et al. Citation2011) are common in PT-born individuals.
Magnetic resonance imaging (MRI) has been widely used to study brain development in PT-born individuals. Compared with MRI to detect morphometric changes, diffusion tensor imaging (DTI) enables researchers to examine WM properties at a microstructural level (Basser Citation1995). One important measure of DTI is fractional anisotropy (FA), which is sensitive to the alignment of white matter fibres and their structural integrity (Schmithorst et al. Citation2002). Several original TBSS studies have observed altered FA values in PT-born individuals. They had a cluster of lower FA located within several major WM fibre tracts compared with the controls, including in the corpus callosum (CC) (Vangberg et al. Citation2006, Skranes et al. Citation2007, Allin et al. Citation2011, Eikenes et al. Citation2011), cingulum (Eikenes et al. Citation2011), external capsule (EC) (Skranes et al. Citation2007, Eikenes et al. Citation2011), internal capsule (IC) (Vangberg et al. Citation2006, Skranes et al. Citation2007), inferior fronto-occipital/longitudinal fasciculus (IFOF/ILF) (Skranes et al. Citation2007, Eikenes et al. Citation2011), superior longitudinal fasciculus (SLF) (Skranes et al. Citation2007, Allin et al. Citation2011) and occipital WM/optic radiation (Vangberg et al. Citation2006). Increased FA has also been found in various WM tracts, including the anterior corona radiata (CR) and the inferior fronto-occipital fasciculus (IFOF) (Allin et al. Citation2011, Padilla et al. Citation2014). However, some studies brought about inconsistent conclusions, reporting no FA group differences in abovementioned structures (Feldman et al. Citation2012a, Loe et al. Citation2013).
To resolve the inconsistency across different studies and figure out the most consistent WM alterations in PT born individuals, we conducted a coordinate-based meta-analysis of the whole-brain studies using seed-based d mapping (SDM) software, which has a technical advantage over other meta-analytical tools in that the same map including positive and negative findings from contributing studies.
Methods
Data source
Comprehensive online searches for literatures were performed in PubMed, Embase, Web of Science and Science Direct databases published before 31th October 2022. The keywords searched were ‘premature labor’ ‘preterm infant’ ‘premature birth’ ‘premature delivery’ or ‘preterm born’; ‘tract-based spatial statistical’ or ‘TBSS’; ‘diffusion tensor’ or ‘DTI’; and ‘fractional anisotropy’ or ‘FA’. Moreover, all the review articles and their references were manually searched to obtain additional papers and raw data. Authors were contacted for any inaccessible raw data of concern. PROSPERO registration is not applicable to this systematic review and meta-analysis.
Study selection and data extraction
Two authors with experience in systematic review independently searched the literature, examined the retrieved articles, and extracted and crosschecked data. The authors then assessed the eligibility of these studies for systematic review. Any discrepancy was discussed and resolved by consensus or a third author.
The included studies should meet the following criteria: (1) analysis of DTI images using TBSS; (2) data were acquired from human infants, adolescents, and adults in the original studies; (3) participants born before 37 weeks’ gestation were included in the preterm born group; (4) healthy age-matched term-born participants were included as a comparison group; and (5) a whole-brain analysis were performed.
The exclusion criteria for this study were as follows: (1) reporting only region of interest (ROI) findings were reported instead of whole-brain results; and (2) reporting only the results of correlation analysis without reporting the results of whole-brain analysis.
To be included in the meta-analysis, studies drawn from the systematic review were required to meet the additional criterion of reporting whole-brain results of FA value alteration in a stereotactic space in three coordinates (x, y, z) according to the SDM method (Radua et al. Citation2014).
Studies that used multiple independent patient samples were separately compared with matching HC groups, and the appropriate coordinates were regarded as separate datasets. The correspondence authors of articles without whole-brain coordinates but were otherwise suitable for meta-analysis were contacted for additional information. There was no language restriction, although all included articles were written in English.
The following data were extracted from eligible studies for meta-analysis: name of the first author, year of publication, sample size, gender, mean age of subjects, GA, birth weight, the scores of Wechsler intelligence scales (either the Wechsler Intelligence Scale for Children-Revised [WISC-R] or the Wechsler Adult Intelligence Scale [WAIS]-III, depending on the subjects’ age), strength of the magnetic field, number of directions, method of voxel-based analysis, threshold for significance and the method used to correct whole-brain results for multiple comparisons.
Quality assessment checklist
The qualities of the studies were assessed using a set of nine weighted criteria specifically for the review and similar to the previously published quality assessment. The checklist was developed from previous meta-analyses focusing on MRI variables (Kmet et al. Citation2004, Welton et al. Citation2015). Although it was not designed as an assessment tool, the checklist provided an objective indication of the rigour in the individual studies. Articles that scored less than a predefined threshold of 50% were excluded from the meta-analysis.
SDM meta-analysis
The meta-analysis was performed using the SDM software package (version 6.11, http://www.sdmproject.com), following a standardised process to compare the FA changes between PT and HC. SDM created effect size and variance maps based on the peak coordinates of each included study. Both negative and positive coordinates were reconstructed in the same map to prevent a particular voxel from appearing to be significant in opposite directions. The re-creation was based on converting the peak t value to Hedges’ effect size and then applying a non-normalised Gaussian kernel to the voxels close to the peak. We used an uncorrecte P < 0.005 as the main threshold (Radua et al. Citation2013). To reduce the possibility of false-positive results further, we used an additional peak-height threshold of Z > 1 and extent threshold of 10 voxels (Radua et al. Citation2013). In particular, a TBSS template based on the FMRIB58 FA skeleton (heep://www.fmrib.ox.ac.uk/fsl) was provided to restrict the analysis to the same WM tracts that were available in the TBSS studies.
Furthermore, to assess the robustness of findings, sensitivity and heterogeneity analyses were conducted. First, to test the replicability of the results, we used a systematic, whole-brain, voxel-based, jack-knife sensitivity analysis. We repeated the main analysis six times, discarding a different study each time. If a previously significant brain region remained significant in all or most of the combinations of the studies, it could be concluded that this finding was highly replicable (Radua et al. Citation2011). Second, between-study variance was analysed to assess the significant heterogeneity of brain abnormalities by using a random-effects model with Q statistics (χ2 distribution converted to z values) and tested with a permutation approach. The potential publication bias was formally assessed with Egger’s tests. We created funnel plots of the peaks of the main findings in order to discard gross abnormalities.
The meta-regressions were analysed with the mean age of subjects at testing, GA, birth weight, percentage of males and the scores of Wechsler intelligence scales (full IQ) in each study as independent variables. To minimise the detection of spurious relationships, we decreased the probability threshold to 0.0005 and required abnormalities to be detected both in the slope and in one of the extremes of the regression, and discarded findings in regions other than those detected in the main analyses. Furthermore, we inspected regression plots to discard fittings driven by too few studies (Radua et al. Citation2013).
Results
Characteristics of the studies included in the systematic review
The search strategy identified 1546 studies, 17 of which met our inclusion criteria. A total of 1065 PT-born individuals and 563 matched HCs in these studies were analysed. The clinical and demographic data was summarised in . The scanning methods and altered FA regions from all included studies were showed in . The flow chart of the search strategy was presented in . The studies by Vangberg et al. (Citation2006) and Skranes et al. (Citation2007), as well as part of Feldman et al. (Citation2012b) and Loe et al. (Citation2013) used overlapping patient samples.
Figure 1. Search strategy used. DTI, diffusion tensor; FA, fractional anisotropy, ROI, region of interest; TBSS, tract-based spatial statistical.
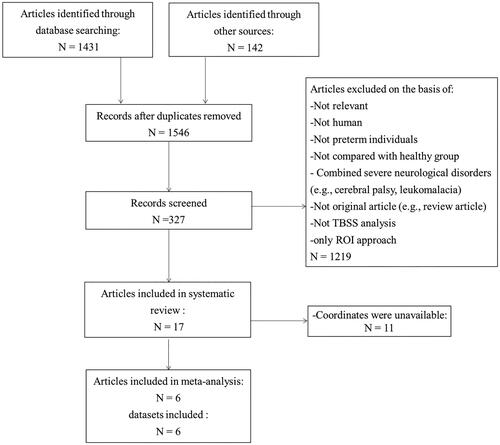
Table 1. Demographic and clinical characteristics of subjects in the 17 PT-TBSS studies included in the systematic review.
Table 2. The main research results in the 17 PT-TBSS studies included in the systematic review.
A total of eight studies focused on adults, five on children and three on infants. Ten studies recruited the very PT–born (GA <29 weeks) individuals, five studies the moderate PT (GA = 29–32 weeks) and one study late PT (GA >32 weeks).
Only six studies reported the three-dimensional coordinates in stereotactic space (Vangberg et al. Citation2006, Allin et al. Citation2011, Feldman et al. Citation2012a, Loe et al. Citation2013, Padilla et al. Citation2014, Salvan et al. Citation2014). A meta-analysis was performed using SDM software based on them.
Quality assessment
All the articles surpassed the minimum quality threshold (Supplementary Table 1). The item that deducted the most points was the acquisition parameters, such as magnet strength and number of diffusion gradient directions.
Narrative review
TBSS study of preterm-born adults
A total of eight studies recruited 650 adults. Vangberg et al. (Citation2006) and Skranes et al. (Citation2007) used overlapping samples. The main finding was that the PT group had significantly reduced FA values in several WM regions, including the CC, IC and superior fasciculus, compared to the control group. The authors speculated that this may be caused by reduced myelination. Skranes reported in detail that low FA values in these areas were associated with perceptual, cognitive, motor and mental health impairments.
Eikenes (Citation2011), Nagy (Citation2009) and Salvan (Citation2014) found that several major WM tracts had significantly reduced FA values in the PT group, for example in the hippocampal fornix and splenium of the CC, ILF and IFOF. Meanwhile, in the PT group, FA was found to positively correlate with birth weight and total IQ.
However, Allin’s study (2011) differed from others, suggesting FA values were increased in clusters included the bilateral IFOF and bilateral anterior CR. Meanwhile, two studies failed to find FA differences between PT-born adults and controls. The study by Feldman et al. (Citation2012a) consisted of convenience samples of children born at two sites: Stanford University (site 1) and University of Pittsburgh (site 2). The participants in the study by Loe et al. study (2013) overlapped with those of site 1 in the Feldman’s study. They found no significant group difference in FA value at either site. The authors believed this may due to many PT-born participants did not have significant WM lesions on conventional MRI.
TBSS study of preterm-born children
Three studies including 411 individuals were focused on school-age and pre-school-age children.
Saunavaara et al. (Citation2017) divided the PT individuals into two subgroups according to the history of foetal growth restriction (FGR). Compared to term-born controls, PT-born children with FGR were found to have lower FA values in the forceps major, forceps minor, bilaterally anterior thalamic radiation, CST, cingulum, IFOF, ILF, SLF and uncinate fasciculus. However, there were no significant differences in FA between non-FGR and HC groups.
Padilla et al. (Citation2014) conducted similar research. Compared to IUGR, PT-born children had reduced FA in the corpus callosum, and increased FA in the anterior corona radiata. No significant differences were identified in any other diffusivity parameter.
The study by Young et al. (Citation2018) of four-year-old children found that lower FA was mainly in the left hemisphere. Within the preterm group, no significant associations were found between FA and any cognitive ability. In contrast, Kelly’s study found that the reduced FA was concentrated in the right hemisphere (Kelly et al. Citation2016b). FA was significantly associated to neurodevelopmental outcomes in PT-born children.
Lee et al. (Citation2013) focused on neurodevelopmental trends in premature infants of different ages. A total of 18 PT-born children and 18 term-born children were divided into three subgroups according to the corrected age at the time of DTI scanning. In the preterm group, FA values of overall WM showed an increase with age. In the children older than two years old, no significant difference between groups was found, except in the CS and genu of CC regions, where the FA values in PT-born children were lower. In the full-term group, most WM structures had reached maturation at around one year of age. This trend indicated that WM maturation was a gradual occurrence during a child’s first two years.
TBSS study of preterm-born infants
Four studies included 413 infants born very preterm (born ≤28 weeks GA) (Anjari et al. Citation2007, Rose et al. Citation2008, Alexandrou et al. Citation2014). They found that the most immature infants displayed the most extensive reductions in FA, suggesting more severe WM microstructural abnormalities with increasing exposure to the ex-utero environment. The largest TBSS study in late-PT infants also found that they exhibited widespread brain WM microstructural alterations (Kelly et al. Citation2016a).
SDM meta-analysis results
Six studies with peak coordinates were included in the meta-analysis (Vangberg et al. Citation2006, Allin et al. Citation2011, Feldman et al. Citation2012a, Loe et al. Citation2013, Padilla et al. Citation2014, Salvan et al. Citation2014). They included a total of 213 PT-born individuals (107 men and 106 women with a mean age of 13.6 [range 1–20.85] years) and 169 matched HC participants (78 men and 91 women with a mean age of 13.5 [range 1–20] years). The mean GA of PT-born individuals was 29.1 weeks [range 28.2–30.0 weeks].
The results of the pooled meta-analysis were provided in and . Compared with controls, reduced FA was found in the CC, bilateral thalamus and left SLF II. None of the clusters showed significant publication bias based on Egger’s test (p > 0.05) (Egger et al. Citation1997).
Figure 2. Four clusters with decreased FA were identified in PT compared with HC: the corpus callosm, bilateral thalamus and the left superior longitudinal fasciculus II. FA, fractional anisotropy; PT, preterm; HC, healthy control. CC, corpus callosm; L, left; R, right; SLF, superior longitudinal fasciculus.
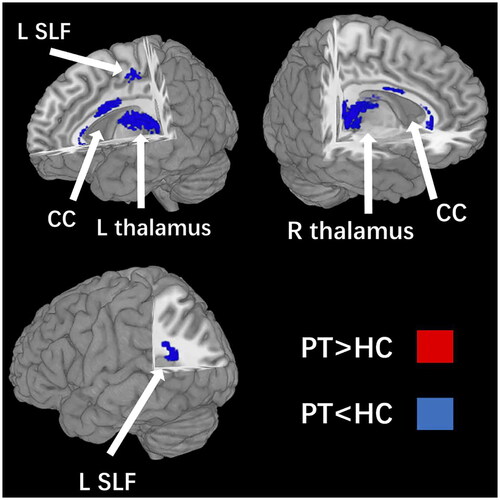
Table 3. Regional differences in FA values between PT and HC subjects identified by the present meta-analyses (voxelwise p < 0.005 and FWHM 20 mm).
As Supplementary Table 2 showed, a jack-knife sensitivity analysis revealed that these clusters were highly reliable, especially the clusters in the CC and the right thalamus, which were significant for all iterations.
To characterise the potential effects of key demographic and clinical variables on WM, we explored the following variables by means of simple linear regression: percentages of male individuals, mean age at testing, GA, birth weight, and the scores of full IQ (available in three studies (Allin et al. Citation2011, Loe et al. Citation2013, Salvan et al. Citation2014)). However, these variables were not significantly linearly associated with PT-related WM FA changes.
Discussion
From 32 weeks of gestation, WM experienced an abrupt increase about five-fold in volume, with the majority of this augmentation happening after week 36 (Hüppi et al. Citation1998). Prior to full term, the foetus resides in the uterus, and is comparatively well shielded from external stimuli. On the contrary, premature infants are subjected to distinct auditory and visual stimuli, and their extra-uterine environment may accelerate WM maturation. This rapid development of the WM particularly in premature infants put it at increased risk of damage (Volpe Citation1981). Additionally, these infants are vulnerable to brain insults, such as maternal infections, high vulnerability to inflammatory mechanisms (Volpe Citation2009), oxidative processes (Haynes et al. Citation2005), and other perinatal complications (Volpe Citation2001). As a result, WM injury is the most common form of brain injury in very premature infants (Volpe Citation1981).
The current study pooled TBSS studies on PT to conduct a quantitative meta-analysis and provided a narrative review of current findings. This is the first SDM meta-analysis of PT-TBSS studies. To resolve the inconsistency across different studies, we conducted a coordinate-based meta-analysis of six whole-brain studies. The DSM analysis combines FA values in children, adolescents, and young adults born PT.
Most robust WM alterations: the CC, the bilateral thalamus and the left SLF II
The meta-analysis highlighted four brain regions in which FA was lower in the PT than in the HC. The quantitative meta-analysis found that the most robust WM alterations be located in the CC, the bilateral thalamus and the left SLF II.
The CC is the largest fibre bundle of the human brain connecting the left and right cerebral hemispheres (McNally et al. Citation2010). The genu of the CC connects the prefrontal and orbitofrontal regions, and the body connects the precentral frontal regions and parietal lobes (Liao et al. Citation2013). CC injury may be due to the inherent vulnerability of immature oligodendrocytes, which makes the developing CC vulnerable to hypoxic–ischemic injury and haemorrhage (Glickstein and Berlucchi Citation2008). Reductions in CC volumes have been described in PT-born children and adolescents (Nosarti et al. Citation2008, Chao et al. Citation2009). Deficits in FA have also been detected in the CC in PT-born infants (Abe et al. Citation2004, Vangberg et al. Citation2006, Skranes et al. Citation2007, Rose et al. Citation2008). The study by Dibble et al. (2021) presented analogous findings. The research included eleven studies that utilised DTI data. The corpus callosum was identified as a region of interest in both the early and moderate-late preterm groups, with statistically significant diffusion measures reported in all eleven studies. FA reductions in the CC may indicate the functional impairment of frontal lobe information transfer between hemispheres and may be associated with impairment of several motor, perceptual, and cognitive functions (Glickstein and Berlucchi Citation2008, Yamada et al. Citation2015). Low FA values of CC are also found in attention-deficit hyperactivity disorder (ADHD) (Skranes et al. Citation2007), major depressive disorder (MDD) (Yamada et al. Citation2015) and bipolar disorder (Yamada et al. Citation2015).
Our study found that the thalamus showed significantly lower FA in the preterm group. Brain volumes in the thalamus are smaller in PT infants than in full-term infants (Counsell et al. Citation2007). The thalamus is known to myelinate earlier, at around 24–25 weeks, and is noted to be more vulnerable to injury in the preterm population (Huang and Castillo Citation2008). The thalamus is involved in language processing and its lesions produce verbal fluency impairment (Crosson Citation1999).
The present meta-analysis identified significantly decreased FA in the left SLF II. The SLF II occupies the central core of the white matter above the insula. It extends from the angular gyrus to the caudal-lateral prefrontal regions (Makris et al. Citation2005) contains long association fibres connecting all the lobes. To date, many DTI studies of PT-born patients have reported decreased FA in the SLF. For example, Hüppi et al. (Citation2001) reported fewer fibre bundles in the SLF in PT infants with perinatal brain damage. Saunavaara et al. (Citation2017) also reported that PT-born children had lower FA values in the SLF. The SLF tract is involved in inter-hemispheric communication (Fjell et al. Citation2010). Low FA values in these tracts may indicate fewer fibre bundles and disturbed fibre organisation that cause cognitive impairments and reduced functioning, for instance, of motor, language articulation and working memory (Makris et al. Citation2005, Skranes et al. Citation2007, Hatton et al. Citation2014).
Meta-regression analyses: no significant associations with age at testing, GA or IQ and PT-related WM FA changes
In our meta-analysis, there were no significant associations with age at testing, GA or IQ and PT-related WM FA changes, at least linearly, but many studies had different results, which may be related to the few studies included in the meta-analysis.
We did not find a linear relationship between FA values and age at testing in meta-analysis, but some recent studies have reported the difference in FA values between PTs and controls seems to increase between birth and young adulthood. At term-equivalent age, the differences in FA are mainly seen in the frontal and central part of the brain (Anjari et al. Citation2007, Rose et al. Citation2008). In childhood, association fibres also become affected (Anjari et al. Citation2007). In the studies of young adults, central WM were still affected, but other affected fibres were the IFOF and ILF (Vangberg et al. Citation2006).
The meta-analysis showed no correlation between different brain regions and GA, which may be because participants in the studies were concentrated between 28 and 30 weeks of gestation. Of all the studies we included in the systematic review, only one study of Kelly et al. (Citation2016a) was of late-PT infants (born at 32–36 GA).
We did not find a linear relationship between FA values and cognitive function in meta-analysis, but some recent studies have reported an association between WM injury and disabilities in children born prematurely. Impairment of WM maturation in PT infants could result in inadequate neurological maturation and clinical dysfunction related to the specific region. FA was found to correlate positively with total IQ in the young PT-born adults (Eikenes et al. Citation2011).
Limitations and conclusions
This meta-analysis has some limitations that must be considered. First, the SDM analyses were conducted using a limited number of studies, because the remaining studies were excluded due to the exclusion criterion, such as control groups born not full-term, studies not targeted at whole-brain research, and individuals with missing coordinate data. Second, a subgroup analysis was not presented and discussed in this study because some subgroups contained only one study, which led to a lack of statistical value. Third, a lot of studies provided limited information about clinical characteristics and social background. Fourth, we utilised the FMRIB58 adult template, which is not designed for neonatal MRI analyses, potentially resulting in analysis deviations. Additionally, we solely assessed alterations in white matter FA values in the brain, with varying results for other indicators which will be explored in upcoming studies.
Conclusion
To our knowledge, this is the first SDM meta-analysis on PT-TBSS studies. Our results indicate that the significant changes in FA in PT-born individuals reflect WM abnormalities from childhood to young adulthood. Abnormal maturation of the CC, bilateral thalamus and left SLF II may be a key target in the neurodevelopment of PT-born individuals. In the future, we will need more data to study the dynamic brain microstructure changes of individuals born with PT, as well as the effects of external and internal factors on brain development.
Contribution to authorship
LZ, XL, and CS conceived and designed the study. LZ and GH searched in databases and conducted the study selection. LZ and MC independently screened studies following the inclusion and exclusion criteria and extracted the data from eligible studies. LZ and SZ assessed the risks of bias of all included studies. CS resolved discrepancies in the aforementioned processes. LZ and XL analysed the data. LZ wrote the manuscript, and then XL and CS critically revised the manuscript. All authors reviewed and approved the final manuscript.
Supplemental Material
Download PNG Image (399.1 KB){kind=link}
Supplemental Material
Download PNG Image (805.6 KB){kind=link}
Supplemental Material
Download PDF (676.6 KB)Acknowledgements
We thank all the authors of the studies included and the subjects.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
Additional information
Funding
References
- Abe, O., et al., 2004. Topography of the human corpus callosum using diffusion tensor tractography. Journal of Computer Assisted Tomography, 28 (4), 533–539.
- Aeby, A., et al., 2013. Language development at 2 years is correlated to brain microstructure in the left superior temporal gyrus at term equivalent age: a diffusion tensor imaging study. NeuroImage, 78, 145–151.
- Alexandrou, G., et al., 2014. White matter microstructure is influenced by extremely preterm birth and neonatal respiratory factors. Acta Paediatrica (Oslo, Norway: 1992), 103 (1), 48–56.
- Allin, M.P., et al., 2011. White matter and cognition in adults who were born preterm. PloS One, 6 (10), e24525.
- Allotey, J., et al., 2018. Cognitive, motor, behavioural and academic performances of children born preterm: a meta-analysis and systematic review involving 64 061 children. BJOG: An International Journal of Obstetrics and Gynaecology, 125 (1), 16–25.
- Anderson, P.J., 2014. Neuropsychological outcomes of children born very pretermed. In: Seminars in Fetal and Neonatal Medicine. Elsevier, 90–96.
- Anjari, M., et al., 2007. Diffusion tensor imaging with tract-based spatial statistics reveals local white matter abnormalities in preterm infants. NeuroImage, 35 (3), 1021–1027.
- Basser, P.J., 1995. Inferring microstructural features and the physiological state of tissues from diffusion-weighted images. NMR in Biomedicine, 8 (7–8), 333–344.
- Bora, S., et al., 2014. Neonatal cerebral morphometry and later risk of persistent inattention/hyperactivity in children born very preterm. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 55 (7), 828–838.
- Chao, Y.P., et al., 2009. Probabilistic topography of human corpus callosum using cytoarchitectural parcellation and high angular resolution diffusion imaging tractography. Human Brain Mapping, 30 (10), 3172–3187.
- Counsell, S.J., et al., 2007. Thalamo-cortical connectivity in children born preterm mapped using probabilistic magnetic resonance tractography. NeuroImage, 34 (3), 896–904.
- Crosson, B., 1999. Subcortical mechanisms in language: Lexical–semantic mechanisms and the thalamus. Brain and Cognition, 40 (2), 414–438.
- Egger, M., Smith, G.D. and Phillips, A.N., 1997. Meta-analysis: principles and procedures. BMJ (Clinical Research ed.), 315 (7121), 1533–1537.
- Eikenes, L., et al., 2011. Young adults born preterm with very low birth weight demonstrate widespread white matter alterations on brain DTI. NeuroImage, 54 (3), 1774–1785.
- Feldman, H.M., et al., 2012a. White matter microstructure on diffusion tensor imaging is associated with conventional magnetic resonance imaging findings and cognitive function in adolescents born preterm. Developmental Medicine and Child Neurology, 54 (9), 809–814.
- Feldman, H.M., et al., 2012b. Language and reading skills in school-aged children and adolescents born preterm are associated with white matter properties on diffusion tensor imaging. Neuropsychologia, 50 (14), 3348–3362.
- Fjell, A.M., et al., 2010. When does brain aging accelerate? Dangers of quadratic fits in cross-sectional studies. NeuroImage, 50 (4), 1376–1383.
- Glickstein, M. and Berlucchi, G., 2008. Classical disconnection studies of the corpus callosum. Cortex; a Journal Devoted to the Study of the Nervous System and Behavior, 44 (8), 914–927.
- Hatton, S.N., et al., 2014. White matter tractography in early psychosis: clinical and neurocognitive associations. Journal of Psychiatry & Neuroscience: JPN, 39 (6), 417–427.
- Haynes, R., et al., 2005. Oxidative and nitrative injury in periventricular leukomalacia: a review. Brain Pathology (Zurich, Switzerland), 15 (3), 225–233.
- Huang, B.Y. and Castillo, M., 2008. Hypoxic-ischemic brain injury: imaging findings from birth to adulthood. Radiographics: A Review Publication of the Radiological Society of North America, Inc, 28 (2), 417–439; quiz 617.
- Hüppi, P. S., et al., 2001. Microstructural brain development after perinatal cerebral white matter injury assessed by diffusion tensor magnetic resonance imaging. Pediatrics, 107 (3), 455–460.,
- Hüppi, P.S., et al., 1998. Quantitative magnetic resonance imaging of brain development in premature and mature newborns. Annals of Neurology, 43 (2), 224–235.
- Kelly, C.E., et al., 2016a. Moderate and late preterm infants exhibit widespread brain white matter microstructure alterations at term-equivalent age relative to term-born controls. Brain Imaging and Behavior, 10 (1), 41–49.
- Kelly, C.E., et al., 2016b. Axon density and axon orientation dispersion in children born preterm. Human Brain Mapping, 37 (9), 3080–3102.
- Kmet, L.M., Cook, L.S. and Lee, R.C., 2004. Standard quality assessment criteria for evaluating primary research papers from a variety of fields.
- Lee, A.Y., et al., 2013. Radiologic differences in white matter maturation between preterm and full-term infants: TBSS study. Pediatric Radiology, 43 (5), 612–619.
- Liao, Y., et al., 2013. Is depression a disconnection syndrome? Meta-analysis of diffusion tensor imaging studies in patients with MDD. Journal of Psychiatry & Neuroscience: JPN, 38 (1), 49–56.
- Loe, I.M., Lee, E.S. and Feldman, H.M., 2013. Attention and internalizing behaviors in relation to white matter in children born preterm. Journal of Developmental and Behavioral Pediatrics: JDBP, 34 (3), 156–164.
- Makris, N., et al., 2005. Segmentation of subcomponents within the superior longitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study. Cerebral Cortex (New York, N.Y.: 1991), 15 (6), 854–869.
- Mcnally, M.A., et al., 2010. Corpus callosum segment circumference is associated with response control in children with attention-deficit hyperactivity disorder (ADHD). Journal of Child Neurology, 25 (4), 453–462.
- Menegaux, A., et al., 2017. Impaired visual short-term memory capacity is distinctively associated with structural connectivity of the posterior thalamic radiation and the splenium of the corpus callosum in preterm-born adults. NeuroImage, 150, 68–76.
- Nagy, Z., et al., 2009. Structural correlates of preterm birth in the adolescent brain. Pediatrics, 124 (5), e964–e972.
- Nosarti, C., et al., 2008. Grey and white matter distribution in very preterm adolescents mediates neurodevelopmental outcome. Brain: a Journal of Neurology, 131 (Pt 1), 205–217.
- Padilla, N., et al., 2014. Differential vulnerability of gray matter and white matter to intrauterine growth restriction in preterm infants at 12 months corrected age. Brain Research, 1545, 1–11.
- Radua, J., et al., 2011. Voxel-based meta-analysis of regional white-matter volume differences in autism spectrum disorder versus healthy controls. Psychological Medicine, 41 (7), 1539–1550.
- Radua, J., et al., 2013. A general approach for combining voxel-based meta-analyses conducted in different neuroimaging modalities. Current Medicinal Chemistry, 20 (3), 462–466.
- Radua, J., et al., 2014. Meta-analysis of functional neuroimaging studies indicates that an increase of cognitive difficulty during executive tasks engages brain regions associated with time perception. Neuropsychologia, 58, 14–22.
- Rose, S.E., et al., 2008. Altered white matter diffusion anisotropy in normal and preterm infants at term-equivalent age. Magnetic Resonance in Medicine, 60 (4), 761–767.
- Saigal, S. and Doyle, L.W., 2008. An overview of mortality and sequelae of preterm birth from infancy to adulthood. Lancet (London, England), 371 (9608), 261–269.
- Salvan, P., et al., 2014. Road work on memory lane—functional and structural alterations to the learning and memory circuit in adults born very preterm. NeuroImage, 102 (Pt 1), 152–161.
- Saunavaara, V., et al., 2017. Very preterm children with fetal growth restriction demonstrated altered white matter maturation at nine years of age. Acta Paediatrica (Oslo, Norway: 1992), 106 (10), 1600–1607.
- Schmithorst, V.J., et al., 2002. Correlation of white matter diffusivity and anisotropy with age during childhood and adolescence: a cross-sectional diffusion-tensor MR imaging study. Radiology, 222 (1), 212–218.
- Skranes, J., et al., 2007. Clinical findings and white matter abnormalities seen on diffusion tensor imaging in adolescents with very low birth weight. Brain: a Journal of Neurology, 130 (Pt 3), 654–666.
- Solomon, M., et al., 2011. From lumping to splitting and back again: atypical social and language development in individuals with clinical-high-risk for psychosis, first episode schizophrenia, and autism spectrum disorders. Schizophrenia Research, 131 (1–3), 146–151.
- Vangberg, T.R., et al., 2006. Changes in white matter diffusion anisotropy in adolescents born prematurely. NeuroImage, 32 (4), 1538–1548.
- Volpe, J.J., 1981. Neurology of the newborn. Major Problems in Clinical Pediatrics, 22, 1–648.
- Volpe, J.J., 2001. Perinatal brain injury: from pathogenesis to neuroprotection. Mental Retardation and Developmental Disabilities Research Reviews, 7 (1), 56–64.
- Volpe, J.J., 2009. Brain injury in premature infants: a complex amalgam of destructive and developmental disturbances. The Lancet Neurology, 8 (1), 110–124.
- Welton, T., et al., 2015. Functionally relevant white matter degradation in multiple sclerosis: a tract-based spatial meta-analysis. Radiology, 275 (1), 89–96.
- Yamada, S., et al., 2015. Microstructural abnormalities in anterior callosal fibers and their relationship with cognitive function in major depressive disorder and bipolar disorder: a tract-specific analysis study. Journal of Affective Disorders, 174, 542–548.
- Young, J.M., et al., 2018. Altered white matter development in children born very preterm. Brain Structure & Function, 223 (5), 2129–2141.