
Abstract
The vitamin E forms γ- and δ-tocopherols (T) inhibit carcinogenesis in animal models; nevertheless, their cancer preventive activities in humans are uncertain. As an initial step to address this issue, we conducted a pilot phase 0 trial to determine the levels of tocopherols and their metabolites in prostate cancer patients undergoing radical prostatectomy. The patients were randomized to no supplementation or two capsules of a γ-T-rich vitamin E mixture daily for 7 or 14 day prior to prostatectomy. Blood and urine samples were collected before supplementation and on the day of surgery, along with prostate tissue, for analysis of tocopherols and their metabolites. Estimated blood loss during surgery was not significantly different across treatment arms and there were no reported adverse events. Prostate tissue levels of γ-T and δ-T were increased after 14 day of supplementation. Their side-chain degradation metabolites (CEHCs and CMBHCs) were significantly elevated in plasma, prostate and urine samples after supplementation for 7 or 14 day. In conclusion, supplementation with γ-T-rich vitamin E increased the prostate levels of γ-T and δ-T. The use of pure γ-T, δ-T or tocopherol mixtures with higher ratio of γ-T or δ-T to α-T is recommended for future studies.
Introduction
Numerous epidemiological studies have investigated the relationship between vitamin E (VE) nutrition and cancer (reviewed in (Citation1–4)). Many studies showed a negative association between dietary VE intake or blood levels of VE (mostly α-tocopherol (T)) and cancer risk, suggesting a cancer preventive effect. However, other studies, including some recent large-scale intervention studies, did not support the hypothesis that VE is cancer preventive (Citation1,Citation2,Citation5–19). In a large chemoprevention trial, the Alpha-Tocopherol, Beta-Carotene Cancer Prevention (ATBC) study, supplementation with VE (given as all rac-α-tocopheryl acetate 50 IU daily) was associated with a 32% decrease in prostate cancer incidence and a 41% decrease in mortality of prostate cancer, when prostate cancer analyzed as a secondary endpoint (Citation11). Results of this trial and the Nutritional Prevention of Cancer (NPC) study with selenium (Citation20) were the basis for the large-scale Selenium and Vitamin E Cancer Prevention Trial (SELECT). In SELECT, high doses of VE (provided as 400 IU of all rac-α-tocopheryl acetate) taken daily for an average of five years, alone or in combination with selenium, did not prevent prostate cancer in a population of relatively healthy men (Citation21). This high dose of α-T was shown to lower plasma levels of γ-T (Citation21). Furthermore, the 7 to 12 year follow-up of the SELECT showed that subjects receiving the α-T supplementation had a hazard ratio of 1.17 for developing prostate cancer (Citation22). While this was a negative result for the primary endpoint of prostate cancer prevention, it is important to note the VE used in this trial was high doses of α-T. Based on data from our laboratory and a review of published studies, we hypothesized that γ-T and δ-T have cancer preventive activity, while α-T does not, in animals and humans with adequate VE nutrition (Citation2). The present study is the first step in testing this hypothesis by conducting a pilot phase 0 trial in prostate cancer patients.
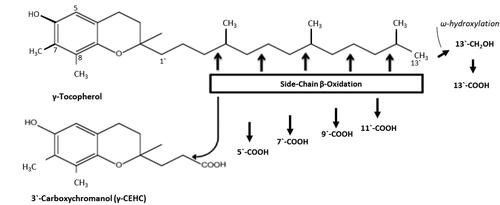
Tocopherols (including the α, β, γ, and δ forms) are major forms of VE and important dietary antioxidants, mainly from vegetable oils and nuts (Citation23). Tocotrienols are the other family of VE, but their dietary intake is generally low, except in individuals consuming large quantities of palm oil. Using γ-T as a base, the structures of these VE forms are shown in . Although γ-T is more abundant in the human diet than α-T, the latter is the major tocopherol found in human and animal tissues. This is because the transfer of VE from the liver to blood is mediated by α-T transfer protein (α-TTP) (Citation24,Citation25), even though all VE forms can be transported to the liver after absorption. α-TTP in the liver preferentially transfers α-T to very low-density lipoproteins, and therefore α-T is preferentially secreted into the circulation and then transferred to nonhepatic tissues. Due to their low affinities for α-TTP, hepatic γ-T, δ-T and tocotrienols are much less efficiently transferred out from the liver, and most of these VE forms undergo side-chain degradation in the liver and other organs. This pathway is initiated with hydroxylation of the ω-methyl group of the side chain by cytochromes P450 4 F or 3 A, and followed by five cycles of β-oxidation in the mitochondria, mainly in the liver, to cut off two-carbon units from the side-chain in each cycle (Citation26). This degradation pathway is illustrated in using γ-T as an example. Larger percentages of γ-T, δ-T and tocotrienols than α-T are degraded through this pathway (Citation27). The γ- and δ-forms of short chain metabolites – carboxyethyl hydroxychromans (CEHCs) and carboxymethylbutyl hydroxychromans (CMBHCs) – are readily measurable in tissue, urine and fecal samples, and they may be used as biomarkers for exposure and metabolism of these VE forms.
Figure 1. Structures of γ-tocopherol and the side-chain degradation pathway. The structures of different vitamin E forms are illustrated with γ-tocopherol as a base, which is dimethylated at the 7- and 8-positions. The chromanol ring is trimethylated at the 5-, 7- and 8-positions in α-tocopherol, dimethylated at the 5- and 8- positions in β-tocopherol and methylated at the 8-position in δ-tocopherol. The corresponding α, β, γ and δ-forms of tocotrienols each have double bonds at the 3`, 7` and 11` positions. The side‐chain degradation pathway is initiated by ω‐oxidation and followed by five cycles of β‐oxidation, each reducing the chain length by two carbons. The metabolites are named following reference Jiang (Citation4); for example, 13′‐COOH is the metabolite of VE with the carboxylic group at the 13′‐position. The degradation of other forms of tocopherols follows the same pathway (from reference Yang et al. (Citation1)).
α-T has been traditionally considered as “the vitamin E” because of its higher tissue abundance and superior activity over other VE forms in the classical fertility-restoration assay (Citation23). Many studies suggest that γ-T, which is not methylated at the 5-position of the chroman ring, can effectively trap reactive nitrogen species and inhibit cyclooxygenase, and is a stronger anti-inflammatory and anticancer agent than α-T (Citation1–4). Because of these properties, γ-T may be more effective than α-T in the prevention of cancer and other diseases. The robust cancer preventive activity of γ-T has been demonstrated in many studies in animal models (reviewed in (Citation1,Citation2)). For example, γ-T has been shown to inhibit prostate carcinogenesis dose-dependently in the transgenic rat for adenocarcinoma of prostate (TRAP) model (Citation28). Dietary administration of a γ-T-rich mixture of tocopherols (γ-TmT) or purified γ-T (or δ-T) inhibited the prostate carcinogenesis in the transgenic adenocarcinoma of the mouse prostate (TRAMP) model as well as models for colon, lung, and mammary carcinogenesis, while α-T was not effective (Citation1,Citation2,Citation29–36). Our recent study also demonstrated that dietary γ-TmT prevented a dietary carcinogen, 2-amino-1-methyl-6-phenylmidazo[4,5-b]pyridine (PhIP)-induced prostate carcinogenesis in CYP1A-humanized mice, and purified δ-T had higher inhibitory activities than γ-T and α-T (Citation33). The preventive effect was associated with the inhibition of many PhIP-induced events: elevation of 8-oxo-deoxyguanosine, nitrotyrosine and the proinflammatory enzyme COX2, as well as the pro-growth Ki-67 and p-AKT signaling. The treatment also prevented the loss of PTEN and Nrf2 expression. In a recent study, δ-T was shown to inhibit prostate carcinogenesis in prostate-specific PTEN-/- (PTENp-/-) mice, in which the activation of AKT is the major driving force for tumorigenesis. However, α-T (2 g/kg diet) was not effective (Citation34).
In contrast to the clear effect of γ-T in prostate cancer prevention in animal studies, results from human studies are inconsistent. For example, in a prospective study by Huang et al., serum γ-T level, but not α-T, was inversely correlated with the risk of developing prostate cancer (Citation5). Similarly, Bauer et al. reported an inverse association between serum γ-T levels and high-grade prostate cancer in an observational study (Citation13). Cheng et al. observed that both γ-T and α-T were inversely associated with the risk of aggressive prostate cancer among current smokers (Citation12) in a nested case-control study of the Carotene and Retinol Efficiency Trial (CARET). Conversely, in the case-cohort study of the SELECT, no significant association between serum levels of γ-T or α-T were observed (Citation37). In a pooled analysis of 15 studies by Key et al., blood levels of α-T were negatively associated with prostate cancer risk, while blood levels of γ-T were not (Citation38).
Based on the strong evidence for the prostate cancer prevention activity of γ-T and γ-TmT in animal models and the inconsistent results in humans, it would be important to test the hypothesis that these agents are cancer preventive in humans. As the first step, we conducted a pilot phase 0 study in prostate cancer patients to evaluate plasma and tissue levels of tocopherols and their metabolites using a commercially available γ-T-rich VE dietary supplement (High Gamma Vitamin E from Nature’s Bounty, Inc) to obtain preliminary information prior to pursuing the development of a pure γ-T or a desired γ-T-rich tocopherol mixture for testing in humans. Prostate cancer patients were treated with this γ-T-rich VE supplement for one or two weeks before prostate surgery; blood, urine and prostate tissue levels of α-, γ-, and δ-tocopherols and their metabolites were determined. As high doses of tocopherols have previously been reported to inhibit blood clotting in animal models (Citation39), we also evaluated blood loss during surgery with the neoadjuvant administration of tocopherol.
Materials and Methods
Eligible patients for this pilot trial included men diagnosed with localized prostate cancer undergoing radical prostatectomy as their primary treatment. Patients were excluded if they had a personal or family history of a bleeding disorder (including individuals taking warfarin or dicumarol); they were taking extra supplementation of VE (a multivitamin containing 60 IU or less of VE was allowable); they had a known history of problems absorbing dietary fats (eg., Crohn’s disease, cystic fibrosis), including patients taking colestipol or orlistat; or they were taking nonsteroidal anti-inflammatory drugs (NSAIDs) or corticosteroids at any time in the two weeks prior to study entry. Patients taking a baby aspirin (81 mg) and those using inhaled or topical steroids were eligible. The trial was approved by the Institutional Review Board of Rutgers, The State University of New Jersey, and the Robert Wood Johnson Medical School (NCT 00895115).
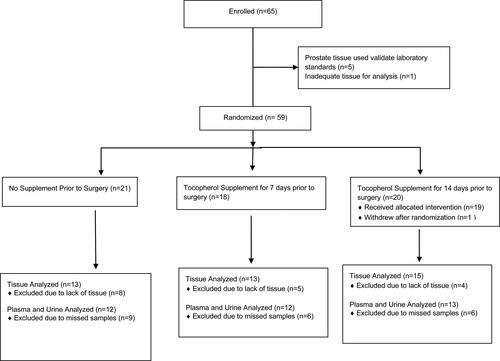
A total of 82 men were screened, 65 were enrolled, 59 were randomized, and one patient withdrew after randomization. Consort standards of clinical trials were followed (see ). The first five patients enrolled received no supplement and their tissue was used to establish laboratory standards and their data is not included in the analyses of the treatment groups. The subsequent patients were randomized by the Rutgers Cancer Institute of New Jersey Biometrics Department and Research Pharmacy to either: 1) no supplementation; 2) daily oral high γ-T-rich VE supplementation for 1 week prior to prostatectomy; or 3) daily oral high γ-T-rich VE supplementation for 2 weeks prior to prostatectomy. Patients randomized to treatment were given a commercially available γ-T-rich VE supplement (High Gamma Vitamin E from Nature's Bounty, Inc., Bohemia, NY 01716) in two soft gel capsules per day, with the last dose taken the evening before prostatectomy.
Figure 2. A consort flowchart of the study.
The supplement was from the same lot number and was stored in a continuously monitored pharmacy at room temperature (20° to 25° C) per USP Storage Conditions. For quality control of the supplement, the composition of different forms of tocopherols were analyzed by HPLC according to the procedure described in the following section. Each capsule contained 128 mg α-T, 200 mg γ-T and 71 mg δ-T, similar to values provided by the supplier. Patient compliance was assessed by pill count, medication diary, and patients were contacted through one to three random telephone calls by the clinical research coordinator or nurse.
Blood and urine samples were collected at baseline and on the day of surgery. Each patient underwent radical prostatectomy with or without bilateral pelvic lymph node dissection (PLND) in the standard manner. All surgeries were robotic prostatectomy. Decisions for carrying out PLND at the time of radical prostatectomy was based on National Comprehensive Cancer Network (NCCN) guidelines. Estimated blood loss during surgery was documented. Once the prostate tissue was removed from the patient, the tissue was immediately delivered to the pathologist who immediately sectioned the tissues to assess the adequacy for pathologic diagnosis. The remaining tissues were immediately snap frozen and stored in the Biospecimen Repository until analysis.
Analysis of Tocopherols and Metabolites
For the analysis, our previous method (Citation30) was used. In brief, both plasma and urine samples were mixed with deionized water and ethanol. Prostate tissues were homogenized in water and ethanol. The lipid-soluble materials in the supernatants were extracted with hexane. For analysis of short-chain metabolites of tocopherols (CEHC & CMBHC), the aqueous phase was extracted with ethyl acetate after incubation with glucuronidase and sulfatase to hydrolyze the conjugated CEHC and CMBHC metabolites. After drying in the speed vacuum concentrator, the residues were re-dissolved in methanol and then injected onto HPLC. The recovery of tocopherols and their metabolites was 70-85%. This was corrected by running samples with known quantities of tocopherols parallelly in the analysis.
Levels of α-T, γ-T and δ-T and the metabolites were quantified using an HPLC system with an electrochemical detector. The HPLC system consists of an ESA Model 542 refrigerated autosampler, two ESA Model 582 dual-piston pumps, and an ESA 5600 coulochem electrode array system (CEAS) with potentials set at 400, −1000, 200, 400, 500, 600, 700 and 800 mV. A SupelcosilTM LC18 reversed-phase column (150 mm x 4.6 mm in diameter; particle size, 5 µm) was used. Solvent A: acetonitrile, 28% (V:V); methanol, 4% (V:V); lithium acetate dihydrate, 30 mM; acetic acid, 2 ml/L; Solvent B: acetonitrile, 84% (V:V); methanol, 13% (V:V); lithium acetate dihydrate, 30 mM; acetic acid, 2 ml/L. The gradient cycle consisted of an initial 84% A and 16% B at a flow rate of 0.6 ml/min, the linear gradient was changed progressively by increasing to 85% B at a flow rate of 0.7 ml/min at 13 min, to 88% B at 0.8 ml/min at 20 min, to 94% B at 1.0 ml/min at 22 min, and to 100% B at 1.2 ml/min at 32 min. The gradient was run at 100% B at 1.2 ml/min until 56 min. Then B was reduced to 16% B at 1.1 ml/min at 56.1 min and stayed at 16% B at 1.1 ml/min until 65 min, then the flow rate was changed to 0.6 ml/min for the next run. The column and CEAS detector were housed in a temperature-regulated compartment maintained at 35 ± 0.1 °C, and the auto sampler was maintained at 6 °C. System control, data acquisition and analysis were performed with the CEAS software.
Statistical Considerations
For each of the tocopherols and their metabolites in plasma and urine samples, pre (before supplementation) and post (after supplementation) values are presented (as mean ± SD). The pre-post differences were compared between treatment groups. The least-squares means of the post-pre differences and the pair-wise comparisons were performed. For the tocopherols and their metabolites in prostate tissue samples, the post supplementation values (tissue obtained after prostatectomy) were compared and pair-wise comparisons were performed. The main analyses were parametric t-tests; normality assumptions were checked and verified the conclusions with non-parametric analyses (Wilcoxon signed-rank test) when the normality assumption was violated. The parametric and non-parametric analyses agreed on their significance/non-significance conclusions in almost all parameters. Results of non-parametric analysis was indicated (see footnote in and ) when the normality assumption did not hold for parametric t-test. The tests were not adjusted for multiple comparisons for this phase 0 trial.
Table 3. Plasma concentrations of tocopherols and their metabolites (µM) before and after supplementation.
Table 4. Prostate tissue levels of tocopherols and their metabolites (µmol/kg) at the time of prostatectomy.
Results
Patients
Between October 2009 and February 2012, a total of 59 patients were randomized in this pilot trial. Characteristics of the patient population are outlined in . There were no significant differences between the treatment groups. Overall, the median age of patients enrolled was 59 years (range 42-77 years) and the median Gleason score was 6. The median baseline PSA was 5.16 ng/mL (range 1-53.7 ng/mL) and the median PSA on the day of surgery was 4.8 ng/mL (range 0.8 − 47.25 ng/mL). Although there was a trend for decreasing PSA with supplementation, this change in PSA was not significant.
Table 1. Characteristics of patients enrolled.
Compliance and Adverse Effects
Compliance was high when assessed by either pill count or patient diary and this is supported by the urine tocopherol metabolite data. There were no adverse effects reported by patients in any of the treatment groups. The median estimated blood loss was 175 mls, 262.5 mls, and 200 mls, in the control, 7-day, and 14-day treatment groups, respectively (). The seemingly higher blood loss in the 7-day group was probably due to a few cases with high blood loss associated with more extensive surgery with lymph node dissection. The blood loss may vary but not one patient received blood transfusion. The difference in blood loss among the three treatment groups was not statistically different.
Table 2. Pathologic Gleason Score and estimated blood loss during surgery by treatment group.
Plasma Levels of Tocopherols and Metabolites
As shown in , mean plasma levels of α-T at baseline and after supplementation were consistently higher than the concentrations of γ-T and δ-T. Supplementation did not significantly increase mean plasma levels of α-T, γ-T, or δ-T, except in the subjects randomized to 1-week supplementation when the pre-post difference of γ-T or δ-T level was compared to the control group by pair-wise comparisons (P < 0.05). However, the data are difficult to interpret because the post-levels were lower than the pre-levels in the control group. A comparison of the effect on tocopherol levels between 1-week and 2-week supplementation groups did not demonstrate a statistically significant difference.
Although α-T levels were higher than γ-T and δ-T levels, higher metabolite levels of γ-CEHC and δ-CEHC than α-CEHC were detected in the plasma, both before and after supplementation. γ-CEHC had the highest mean values after supplementation in both 1-week and 2-week treatment groups. The γ- and δ-CEHC levels of the 1-week or 2-week treatment groups were significantly higher than the pre-supplementation levels (P < 0.05). Although the mean γ-CEHC and δ-CEHC levels for the 1-week supplementation group appeared higher than the 2-week group, the difference was not significant (P = 0.25).
Supplementation significantly increased γ- and δ-CMBHC levels, but had no effect on α-CMBHC levels in the plasma. γ-CMBHC levels were the highest in plasma, both before and after supplementation. While the 1-week treatment group had higher γ- and δ-CMBHC concentrations than the 2-week group, this difference was not statistically significant.
Prostate Tissue Levels of Tocopherols and Metabolites
Overall, the α-T levels were the highest in the prostate tissues, followed by γ-T, and then δ-T (). In patients randomized to the 1-week supplementation group, mean tissue levels of γ-T and δ-T were higher than, but not statistically significant from, those in the control group. In the 2-week supplementation group, the higher values of γ-T and δ-T reached statistical significance (P < 0.05 for γ-T; P < 0.005 for δ-T). However, there were no differences in tissue levels for any form of tocopherol when comparing the 1-week and 2-week supplementations.
There were no differences in the levels of α-CEHC in the prostate tissue between the groups receiving the supplement and the control group (). For γ-CEHC, the levels in the 1-week group were 6-fold higher than the pre-supplementation levels (P < 0.005). The levels of the 2-week group were also higher; however the difference was not statistically significant because of the large standard deviations, possibly due to the individual differences in the side-chain degradation of tocopherols. The tissue levels of δ-CEHC in the 2-week supplementation group were the highest and were significantly higher than the control group (P < 0.005).
Overall, α-CMBHC concentrations in the prostate tissue did not change significantly after supplementation (P = 0.50 for 1-week; P = 0.77 for 2-week). However, both γ-CMBHC and δ-CMBHC levels significantly increased in the prostate tissue with supplementation. The changes in γ-CMBHC and δ-CMBHC levels after supplementation were statistically significant for both supplementation groups when compared to the pre-supplementation levels (P < 0.05). Additionally, the mean values of γ-CMBHC and δ-CMBHC levels in the 2-week supplementation group were higher than their levels in the 1-week group; however, no statistically significant difference was detected between the two treatment groups.
Urine Levels of Tocopherol Metabolites
Tocopherols as fat-soluble vitamins are not excreted in the urine. We assessed their metabolites, CEHC and CMBHC. Urinary levels of γ-CEHC and δ-CEHC were much higher than those of α-CEHC (). While all forms of CEHC increased with both the 1-week and 2-week supplementation, changes in γ-CEHC and δ-CEHC levels were several-fold higher and statistically significant (P < 0.05) when compared to the pre-supplementation levels and the control group. All forms of CMBHC, including the α-CMBHC, were elevated after supplementation, and the increases were statistically significant in the 2-week supplementation group (P < 0.05 for α-T and γ-T; P < 0.005 for δ-T). In the 1-week supplementation group, however, only δ-CMBHC was significantly elevated (P < 0.005).
Table 5. Urinary concentrations of tocopherol metabolites (µM) before and after supplementation.
Discussion
We successfully conducted a pilot phase 0 trial with a high dose (800 mg daily) of a γ-T-rich VE supplement in prostate cancer patients before radical prostatectomy. This product was selected because it was commercially available for human use, allowing us to obtain preliminary information on plasma, tissues, and urine levels of tocopherols and their metabolites. High doses of tocopherols have previously been reported to inhibit blood clotting in animal models (Citation39). This was a major concern for the use of neoadjuvant tocopherol in this trial; therefore, patients with a bleeding history were excluded. In this trial, no adverse effects, including prolonged or excessive bleeding, were observed. The effect of the treatment on PSA levels was not statistically significant.
An important finding from our trial is that two weeks of supplementation significantly increased the levels of γ-T and δ-T levels in prostate tissue, even though their levels in the plasma were not increased. However, the levels of side-chain degradation metabolites (CEHCs and CHBMCs) of γ-T and δ-T were all increased in the plasma, prostate tissue and urine samples. To our knowledge, this is the first report of the level of different forms of tocopherols and their metabolites in human blood, prostate tissue and urine after supplementation with a mixture of tocopherols.
Upon examining the results from our pilot study, the study design with 18-21 subjects in each arm appears under-powered. With more subjects some of the differences in tocopherol levels may reach statistical significance. A limitation of this study is that the biological effects of supplementation with γ-T-rich VE were not determined. In future studies this information may be obtained by measuring cell proliferation (Ki67), apoptosis (cleaved-caspace-3) and other markers in prostate tissues with immunohistochemistry.
The lack of a robust increase of γ-T levels in the plasma after the γ-T-rich VE supplementation was unexpected but worth discussing. This is probably due to the rather low γ-T to α-T ratio in the tocopherol mixture used (α-T: γ-T: δ-T at ratios of 1: 1.56: 0.55). In our previous studies in animal models, dietary supplementation with γ-TmT, which contained 13% α-T, 57% γ-T, 24% δ-T and 0.5% β-T (at ratios of 1: 4.38: 1.85), consistently increased levels of γ-T in the plasma and different tissues (Citation30–33,Citation35,Citation36,Citation40–42). Tocopherols ingested from the diet are known to be absorbed in the intestinal mucosa and are transported to the liver via the lymphatic system. In the liver, α-TTP selectively mediates the transfer of α-T into very low-density lipoprotein and then into the blood (Citation23). γ-T, with lower affinity to α-TTP, is less efficiently transported to the blood through this mechanism. δ-T, with an even lower affinity for α-TTP, is not efficiently transferred. There are suggestions that δ-T may be transferred to the blood through mechanisms independent of α-TTP. As a consequence, α-T is found in much higher levels in the blood and non-hepatic tissues than γ-T and δ-T. Competition between α-T and γ-T for α-TTP has been observed both in animal models and in humans, that is, when high concentrations of α-T were given, the blood levels of γ-T decreased (Citation21). This may be the basis for the lack of increase in plasma levels of γ-T, even though the VE supplement contained more γ-T than α-T. For future studies, in order to raise the blood and non-hepatic tissue levels of γ-T, pure γ-T or a preparation with a much higher γ-T to α-T ratio should be used.
The γ-T and δ-T in the liver are metabolized through the side-chain degradation pathway (), with CEHCs and CMBHCs as the readily measurable short-chain metabolites (Citation30–33,Citation43). This is the biochemical basis for the observed high concentrations of γ- and δ- forms of CEHCs and CMBHCs in different samples in the supplemented groups. When high doses of α-T were given, α-CEHC and α-CMBHC were also observed in biological samples. These side-chain degradation metabolites of different tocopherols, excreted as glucuronide- and sulfate-conjugates in the urine, together with plasma levels of α-T, γ-T, δ-T and their metabolites, could be used to assess the dietary exposure and metabolism of tocopherols in human subjects. Since the urinary levels of γ- and δ- forms of CEHCs and CMBHCs are higher than those in the blood (as well as γ-T and δ-T in the blood), these urinary metabolites may be convenient exposure markers for γ-T and δ-T.
Whether VE prevents or promotes cancer is an intriguing topic. Based on our review of the results from animal and human studies, we propose that at nutritional levels, α-T, γ-T and δ-T are all cancer preventive; however, at supra-nutritional levels, only γ-T and δ-T are effective (Citation1). Possible mechanisms of cancer prevention by tocopherols include antioxidant action, which is shared by all forms of tocopherols (Citation1). In addition, because of unmethylated carbon at the 5-position of the chromanol ring, γ-T and δ-T can more effectively quench reactive nitrogen species than α-T (Citation2). Recent results from animal and cell line studies also suggest that δ-T can more effectively decrease the phosphorylation of AKT than other tocopherols (Citation33,Citation44). An interesting possibility for the higher cancer preventive activities of γ-T and δ-T is the possible cancer preventive activities of their side-chain metabolites. Longer-chain metabolites, such as 13′-carboxychromanols (13′-COOHs) and 11′-COOHs, have been shown to lower prostaglandin levels and to have anti-inflammatory, anti-proliferative and apoptosis promoting activities (Citation4). All these activities may result in cancer prevention. Short-chain metabolites – CEHCs and CMBHCs – retaining the intact chromanol ring structure, may also contribute to cancer preventive activity.
Based on the above results and discussions, we believe additional human studies with pure γ-T or δ-T or tocopherol mixtures with higher γ-T or δ-T to α-T ratio are needed to assess their preventive activity against prostate and other cancers.
Author Contribution Statement
S. Goodin, I. Kim, C. Yang planned the study. I. Kim and M. Orlick conducted the clinical procedures and studies. C. Yang and M. Lee conducted the laboratory analysis of tocopherols and tocopherol metabolites. S. Goodin and X. Zheng coordinated the studies. W. Shih conducted the statistical analyses. C. Yang, S. Goodin, I. Kim and W. Shih wrote the manuscript.
Institutional Approval
The trial was approved by the Institutional Review Board of Rutgers, The State University of New Jersey, and the Robert Wood Johnson Medical School (NCT 00895115).
Disclosure Statement
The authors have no conflicts of interest including any financial, personal, or other relationships with people or organizations that could influence the present article.
Additional information
Funding
References
- Yang CS, Luo P, Zeng Z, Wang H, Malafa M, Suh N. Vitamin E and cancer prevention: Studies with different forms of tocopherols and tocotrienols. Mol Carcinog. 2020;59(4):365–89. ) 2020 Feb 3. doi:https://doi.org/10.1002/mc.23160
- Yang CS, Suh N, Kong AN. Does vitamin E prevent or promote cancer? Cancer Prev Res (Phila)). 2012;5(5):701–5. doi:https://doi.org/10.1158/1940-6207.CAPR-12-0045
- Ju J, Picinich SC, Yang Z, Zhao Y, Suh N, Kong AN, Yang CS. Cancer-preventive activities of tocopherols and tocotrienols. Carcinogenesis. 2010;31(4):533–42. doi:https://doi.org/10.1093/carcin/bgp205
- Jiang Q. Natural forms of vitamin E and metabolites-regulation of cancer cell death and underlying mechanisms. IUBMB Life. 2019;71(4):495–506. doi:https://doi.org/10.1002/iub.1978
- Huang HY, Alberg AJ, Norkus EP, Hoffman SC, Comstock GW, Helzlsouer KJ. Prospective study of antioxidant micronutrients in the blood and the risk of developing prostate cancer. Am J Epidemiol. 2003;157(4):335–44. doi:https://doi.org/10.1093/aje/kwf210
- Wright ME, Weinstein SJ, Lawson KA, Albanes D, Subar AF, Dixon LB, Mouw T, Schatzkin A, Leitzmann MF. Supplemental and dietary vitamin E intakes and risk of prostate cancer in a large prospective study. Cancer Epidemiol Biomarkers Prev. 2007;16(6):1128–35. doi:https://doi.org/10.1158/1055-9965.EPI-06-1071
- Weinstein SJ, Wright ME, Lawson KA, Snyder K, Mannisto S, Taylor PR, Virtamo J, Albanes D. Serum and dietary vitamin E in relation to prostate cancer risk. Cancer Epidemiol Biomarkers Prev. 2007;16(6):1253–9. doi:https://doi.org/10.1158/1055-9965.EPI-06-1084
- Mahabir S, Schendel K, Dong YQ, Barrera SL, Spitz MR, Forman MR. Dietary alpha-, beta-, gamma- and delta-tocopherols in lung cancer risk. Int J Cancer. 2008;123(5):1173–80. doi:https://doi.org/10.1002/ijc.23649
- Weinstein SJ, Peters U, Ahn J, Friesen MD, Riboli E, Hayes RB, Albanes D. Serum alpha-tocopherol and gamma-tocopherol concentrations and prostate cancer risk in the PLCO Screening Trial: a nested case-control study. PLoS One. 2012;7(7):e40204. doi:https://doi.org/10.1371/journal.pone.0040204
- Zhang W, Shu XO, Li H, Yang G, Cai H, Ji BT, Gao J, Gao YT, Zheng W, Xiang YB. Vitamin intake and liver cancer risk: a report from two cohort studies in China. J Natl Cancer Inst. 2012;104(15):1173–81. doi:https://doi.org/10.1093/jnci/djs277
- Heinonen OP, Albanes D, Virtamo J, Taylor PR, Huttunen JK, Hartman AM, Haapakoski J, Malila N, Rautalahti M, Ripatti S, et al. Prostate cancer and supplementation with alpha-tocopherol and beta-carotene: incidence and mortality in a controlled trial. J Natl Cancer Inst. 1998;90(6):440–6. doi:https://doi.org/10.1093/jnci/90.6.440
- Cheng TY, Barnett MJ, Kristal AR, Ambrosone CB, King IB, Thornquist MD, Goodman GE, Neuhouser ML . Genetic variation in myeloperoxidase modifies the association of serum α-tocopherol with aggressive prostate cancer among current smokers . J Nutr. 2011;141(9):1731–7. doi:https://doi.org/10.3945/jn.111.141713
- Bauer SR, Richman EL, Sosa E, Weinberg V, Song X, Witte JS, Carroll PR, Chan JM. Antioxidant and vitamin E transport genes and risk of high-grade prostate cancer and prostate cancer recurrence. Prostate. 2013;73(16):1786–95. doi:https://doi.org/10.1002/pros.22717
- Antwi SO, Steck SE, Su LJ, Hebert JR, Zhang H, Fontham ET, Smith GJ, Bensen JT, Mohler JL, Arab L. Dietary, supplement, and adipose tissue tocopherol levels in relation to prostate cancer aggressiveness among African and European Americans: The North Carolina-Louisiana Prostate Cancer Project (PCaP). Prostate. 2015;75(13):1419–35. doi:https://doi.org/10.1002/pros.23025
- Chen G, Wang J, Hong X, Chai Z, Li Q. Dietary vitamin E intake could reduce the risk of lung cancer: evidence from a meta-analysis. Int J Clin Exp Med. 2015;8(4):6631–7.
- Chen JM, Jiang WX, Shao LM, Zhong DD, Wu YH, Cai JT. Association between intake of antioxidants and pancreatic cancer risk: a meta-analysis. Int J Food Sci Nutr. 2016;67(7):744–53. doi:https://doi.org/10.1080/09637486.2016.1197892
- Wang YY, Wang XL, Yu ZJ. Vitamin C and E intake and risk of bladder cancer: a meta-analysis of observational studies. Int J Clin Exp Med. 2014;7:4154–64.
- Cui L, Li L, Tian Y, Xu F, Qiao T. Association between Dietary Vitamin E Intake and Esophageal Cancer Risk: An Updated Meta-Analysis. Nutrients. 2018;10(7):801. doi:https://doi.org/10.3390/nu10070801
- Hu X, Li S, Zhou L, Zhao M, Zhu X. Effect of vitamin E supplementation on uterine cervical neoplasm: A meta-analysis of case-control studies. PLoS One. 2017;12(8):e0183395 doi:https://doi.org/10.1371/journal.pone.0183395
- Clark LC, Combs GF, Jr., Turnbull BW, Slate EH, Chalker DK, Chow J, Davis LS, Glover RA, Graham GF, Gross EG, et al. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention of Cancer Study Group. Jama. 1996;276(24):1957–63. doi:https://doi.org/10.1001/jama.1996.03540240035027
- Lippman SM, Klein EA, Goodman PJ, Lucia MS, Thompson IM, Ford LG, Parnes HL, Minasian LM, Gaziano JM, Hartline JA, et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). Jama. 2009;301(1):39–51., Jr. doi:https://doi.org/10.1001/jama.2008.864
- Klein EA, Thompson IM, Jr., Tangen CM, Crowley JJ, Lucia MS, Goodman PJ, Minasian LM, Ford LG, Parnes HL, Gaziano JM, et al. Vitamin E and the risk of prostate cancer: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA. 2011;306(14):1549–56. doi:https://doi.org/10.1001/jama.2011.1437
- Traber MG, In Bowman BA, Russell RM, International Life Sciences Institute-Nutrition, F. Present knowledge in nutrition. Washington, D.C: ILSI Press, International Life Sciences Institute; 2006.
- Qian J, Morley S, Wilson K, Nava P, Atkinson J, Manor D. Intracellular trafficking of vitamin E in hepatocytes: the role of tocopherol transfer protein. J Lipid Res. 2005;46(10):2072–82. doi:https://doi.org/10.1194/jlr.M500143-JLR200
- Morley S, Cecchini M, Zhang W, Virgulti A, Noy N, Atkinson J, Manor D. Mechanisms of ligand transfer by the hepatic tocopherol transfer protein. J Biol Chem. 2008;283(26):17797–804. doi:https://doi.org/10.1074/jbc.M800121200
- Sontag TJ, Parker RS. Cytochrome P450 omega-hydroxylase pathway of tocopherol catabolism. Novel mechanism of regulation of vitamin E status. J Biol Chem. 2002;277(28):25290–6. doi:https://doi.org/10.1074/jbc.M201466200
- Sontag TJ, Parker RS. Influence of major structural features of tocopherols and tocotrienols on their omega-oxidation by tocopherol-omega-hydroxylase. J Lipid Res. 2007;48(5):1090–8. doi:https://doi.org/10.1194/jlr.M600514-JLR200
- Takahashi S, Takeshita K, Seeni A, Sugiura S, Tang M, Sato SY, Kuriyama H, Nakadate M, Abe K, Maeno Y, et al. Suppression of prostate cancer in a transgenic rat model via gamma-tocopherol activation of caspase signaling. Prostate. 2009;69(6):644–51. doi:https://doi.org/10.1002/pros.20915
- Barve A, Khor TO, Nair S, Reuhl K, Suh N, Reddy B, Newmark H, Kong AN. Gamma-tocopherol-enriched mixed tocopherol diet inhibits prostate carcinogenesis in TRAMP mice. Int J Cancer. 2009;124(7):1693–9. doi:https://doi.org/10.1002/ijc.24106
- Guan F, Li G, Liu AB, Lee MJ, Yang Z, Chen YK, Lin Y, Shih W, Yang CS . δ- and γ-tocopherols, but not α-tocopherol, inhibit colon carcinogenesis in azoxymethane-treated F344 rats . Cancer Prev Res (Phila)). 2012;5(4):644–54. doi:https://doi.org/10.1158/1940-6207.CAPR-11-0521
- Chen JX, Liu A, Lee MJ, Wang H, Yu S, Chi E, Reuhl K, Suh N, Yang CS . δ- and γ-tocopherols inhibit phIP/DSS-induced colon carcinogenesis by protection against early cellular and DNA damages. Mol Carcinog. 2017;56(1):172–83. doi:https://doi.org/10.1002/mc.22481
- Das Gupta S, So JY, Wall B, Wahler J, Smolarek AK, Sae-Tan S, Soewono KY, Yu H, Lee MJ, Thomas PE, et al. Tocopherols inhibit oxidative and nitrosative stress in estrogen-induced early mammary hyperplasia in ACI rats. Mol Carcinog. 2015;54(9):916–25. doi:https://doi.org/10.1002/mc.22164
- Chen JX, Li G, Wang H, Liu A, Lee MJ, Reuhl K, Suh N, Bosland MC, Yang CS. Dietary tocopherols inhibit PhIP-induced prostate carcinogenesis in CYP1A-humanized mice. Cancer Lett. 2016;371(1):71–8. doi:https://doi.org/10.1016/j.canlet.2015.11.010
- Wang H, Yang X, Liu A, Wang G, Bosland MC, Yang CS . δ-Tocopherol inhibits the development of prostate adenocarcinoma in prostate specific Pten-/- mice. Carcinogenesis. 2018;39(2):158–69. doi:https://doi.org/10.1093/carcin/bgx128
- Lambert JD, Lu G, Lee MJ, Hu J, Ju J, Yang CS. Inhibition of lung cancer growth in mice by dietary mixed tocopherols. Mol Nutr Food Res. 2009;53(8):1030–5. doi:https://doi.org/10.1002/mnfr.200800438
- Li GX, Lee MJ, Liu AB, Yang Z, Lin Y, Shih WJ, Yang CS . δ-tocopherol is more active than α - or γ -tocopherol in inhibiting lung tumorigenesis in vivo . Cancer Prev Res (Phila)). 2011;4(3):404–13. doi:https://doi.org/10.1158/1940-6207.CAPR-10-0130
- Albanes D, Till C, Klein EA, Goodman PJ, Mondul AM, Weinstein SJ, Taylor PR, Parnes HL, Gaziano JM, Song X, et al. Plasma tocopherols and risk of prostate cancer in the Selenium and Vitamin E Cancer Prevention Trial (SELECT). Cancer Prev Res (Phila)). 2014;7(9):886–95. doi:https://doi.org/10.1158/1940-6207.CAPR-14-0058
- Key TJ, Appleby PN, Travis RC, Albanes D, Alberg AJ, Barricarte A, Black A, Boeing H, Bueno-de-Mesquita HB, Chan JM, Endogenous Hormones Nutritional Biomarkers, et al. Carotenoids, retinol, tocopherols, and prostate cancer risk: pooled analysis of 15 studies. Am J Clin Nutr. 2015;102(5):1142–57. doi:https://doi.org/10.3945/ajcn.115.114306
- Takahashi O. Haemorrhagic toxicity of a large dose of alpha-, beta-, gamma- and delta-tocopherols, ubiquinone, beta-carotene, retinol acetate and L-ascorbic acid in the rat. Food Chem Toxicol. 1995;33(2):121–8. doi:https://doi.org/10.1016/0278-6915(94)00120-D
- Ju J, Hao X, Lee MJ, Lambert JD, Lu G, Xiao H, Newmark HL, Yang CS. A gamma-tocopherol-rich mixture of tocopherols inhibits colon inflammation and carcinogenesis in azoxymethane and dextran sulfate sodium-treated mice. Cancer Prev Res (Phila). 2009;2(2):143–52. doi:https://doi.org/10.1158/1940-6207.CAPR-08-0099
- Suh N, Paul S, Lee HJ, Ji Y, Lee MJ, Yang CS, Reddy BS, Newmark HL. Mixed tocopherols inhibit N-methyl-N-nitrosourea-induced mammary tumor growth in rats. Nutr Cancer. 2007;59(1):76–81. doi:https://doi.org/10.1080/01635580701419022
- Smolarek AK, So JY, Thomas PE, Lee HJ, Paul S, Dombrowski A, Wang CX, Saw CL, Khor TO, Kong AN, et al . Dietary tocopherols inhibit cell proliferation, regulate expression of ERα, PPARγ, and Nrf2, and decrease serum inflammatory markers during the development of mammary hyperplasia. Mol Carcinog. 2013;52(7):514–25. doi:https://doi.org/10.1002/mc.21886
- Lee MJ, Feng W, Yang L, Chen YK, Chi E, Liu A, Yang CS. Methods for efficient analysis of tocopherols, tocotrienols and their metabolites in animal samples with HPLC-EC. J Food Drug Anal. 2018;26(1):318–29. doi:https://doi.org/10.1016/j.jfda.2017.07.012
- Wang H, Hong J, Yang CS . δ-Tocopherol inhibits receptor tyrosine kinase-induced AKT activation in prostate cancer cells. Mol Carcinog. 2016;55(11):1728–38. doi:https://doi.org/10.1002/mc.22422