
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Kinetics and full body kinematics were measured in ten elite goalkeepers diving to save high and low balls at both sides of the goal, aiming to investigate their starting position, linear and angular momentum, and legs' contribution to end-performance. Our results showed that goalkeepers adopted a starting position with a stance width of 33 ± 1% of leg length, knee flexion angle of 62 ± 18° and hip flexion angle of 63 ± 18°. The contralateral leg contributed more than the ipsilateral leg to COM velocity (p < 0.01), both for the horizontal (2.7 ± 0.1 m·s−1 versus 1.2 ± 0.1 m·s−1) and for the vertical component (3.1 ± 0.3 m·s−1 versus 0.4 ± 0.2 m·s−1). Peak horizontal and peak angular momenta were significantly larger (p < 0.01) for low dives than for high dives with a mean difference of 55 kg·m·s−1 and 9 kg·m2·s−1, respectively. In addition, peak vertical momentum was significantly larger (p < 0.01) for high dives with a mean difference between dive heights of 113 kg·m·s−1. Coaches need to highlight horizontal lateral skills and exercises (e.g. sideward push-off, sideward jumps), with emphasis on pushing-off with the contralateral leg, when training and assessing goalkeeper’s physical performance.
Introduction
Optimization of performance in football (soccer), or any other team ball sports, relies on knowledge from disciplines as diverse as biomechanics, physiology and psychology (Barlett, Citation1999; Powers & Howley, Citation2007; Williams, Davids, & Williams, Citation1999). In football, the goalkeeper has a unique and critical role in the team, which requires timed and explosive adjustments of body speed, position and orientation in response to a stimulus. Research has mainly focused on the psychological aspects of this task, focusing on perceptual and cognitive skills used in the diving save during penalty kicks (Franks & Hanvey, Citation1997; Kuhn, Citation1988; Savelsbergh, Williams, Van der Kamp, & Ward, Citation2002, Citation2005; Williams, Citation2000). Biomechanical studies are scarce, possibly because of the role differentiation between legs and the high impact at landing, which make full-body kinematic and kinetic measurements of the diving save challenging to perform. To our knowledge, only two studies have investigated the biomechanics of goalkeeper’s diving saves (Spratford, Mellifont, & Burkett, Citation2009; Suzuki, Togari, Isokawa, Ohashi, & Ohgushi, Citation1987). Suzuki et al. (Citation1987) calculated the path of goalkeeper’s centre of mass (COM) in diving to save balls suspended at three different heights (0.30 m, 0.90 m and 1.50 m) at one side of the goal. The measurements were performed on four goalkeepers (two high-level and two novice goalkeepers) in two dimensions and at a relatively low sampling rate of 60 samples/s. The data showed that high-level goalkeepers dived more directly towards the ball, with greater velocity than novice goalkeepers. Spratford et al. (Citation2009) compared dives between goalkeeper’s preferred and non-preferred sides. They performed the measurements on 6 goalkeepers (under-20 Australian national team) at a sampling rate of 120 samples/s. They found limited asymmetry in the movement patterns between dive sides: a greater COM net projection angle and greater pelvic and torso rotations at initiation, when diving towards the non-preferred side.
Both studies used balls suspended directly to the side of the goalkeepers (under the high post), constraining them to perform a frontal plane dive. This is different from the diving save performed in game situations, where the goalkeepers prefer to dive diagonally (sideward and forward), so they can reach the ball at a better angle and effectively reduce the goal area that they need to cover. Moreover, balls were always suspended at one height at a time, allowing for height-specific adjustments and anticipation before the initiation of the dive.
The diving save is a complex motor task characterized by large forces exerted at push-off generated in a very short period of time, and in many cases in absence of prior knowledge of ball location. Therefore, the starting position and the push-off technique are most likely key factors in diving save performance. However, starting position, and dive side and height dependent leg contributions without prior knowledge of ball location, were not studied before. Therefore, the aims of this study were to document the characteristics of the starting position and to analyse how each leg contributes to the dive towards high and low balls at both sides of the goal, in the absence of prior knowledge of ball location. In addition, total body linear and angular momentum were compared between dive heights and sides. Based on goalkeeper coaches’ input, the ipsilateral leg push-off (the leg on the diving side) was hypothesized to contribute the most to the diving save. We also hypothesized that high dives would require greater linear momentum, while low dives may need greater angular momentum.
Methods
Ten elite football goalkeepers, mean ± standard deviation age 18.4 ± 2.6 years, mass 82.6 ± 9.1 kg, height 186 ± 2.4 cm, and dominant leg 9 right and 1 left, participated in this study. The participants’ level, at the time of the experiment, was as follows: two goalkeepers in the Dutch Eredevisie (the highest level of competition nationally), six goalkeepers in the Dutch Eerste Divisie (the second highest level of competition nationally), and two goalkeepers in the Dutch under-17 Eredevisie (the highest level of competition nationally for players under 17 years of age). Before performing the experiment participants or their parents signed an informed consent form. For each participant, anthropometric measurements, age and injury history were gathered. Participants had not suffered any injury that prevented them from performing the diving save at their maximum power or caused them to change their movement pattern at the time of the experiment. The experiments were conducted at the Adidas miCoach Performance Centre of Amsterdamsche Football Club Ajax (AFC Ajax). The Ethics Committee of the Faculty of Behavioral and Movement Sciences of the Vrije Universiteit Amsterdam had approved the research protocol.
Data collection and pre-processing
Before starting the measurement, the participants performed a goalkeeper specific warm-up routine with their coaches and around 8 diving saves to get familiar with the experimental set-up. Each participant was then instructed to dive as fast as possible in response to a visual stimulus, to the correct ball side and height. For each participant, two dives were measured for two heights at both sides of the goal, for a total of 8 dives per participant with 2 minutes recovery time between dives. The visual stimulus was produced by a light-emitting diode (LED) board placed at the penalty spot, consisting of four LED lamps indicating the side and height of the ball to save. The balls were suspended 1m in front of the goal line, at two heights from the force plates’ level (30 and 190 cm). They were attached to a thin rope by a magnet allowing the goalkeeper to hit or catch the ball. The Balls could easily be taken off the magnet, so the goalkeepers did not experience any significant resistance when saving a ball. A ball was suspended at low height on one side, and the other ball was suspended at high height on the other side, and they were re-attached to the ropes after each dive, to avoid any anticipation of height and side.
A passive marker motion analysis system (Vicon 612, Oxford United Kingdom) was used after calibration with an active wand comprising five markers. Trajectory data from 44 markers was captured using 10 infrared cameras at 200 samples/s (). Single markers were attached to different body segments in the form of clusters (feet, shanks, pelvis, thorax, head and forearms), the thighs were modelled between the shanks and pelvis, and the upper arms were modelled between the thorax and forearms, in order to obtain a full-body model without occlusion of the markers during the trials and limiting the risk of landing on markers. The markers were attached in a well recognizable pattern to facilitate the labelling with Vicon Nexus Software (version 1.8.5). Soft markers were used on areas that are prone to impact at landing, and 3 single markers were attached to each ball. Anatomical coordinate systems of the segments were marked with single markers and related to the corresponding marker clusters during a measurement in a reference position (T-pose). Details of the three-dimensional inverse dynamics model that was used in this study can be found elsewhere (Faber, Chang, Kingma, & Dennerlein, Citation2013; Faber, Kingma, & van Dieën, Citation2011; Ibrahim, Faber, Kingma, & van Dieën, Citation2016; Kingma, de Looze, Toussaint, Klijnsma, & Bruijnen, Citation1996).
Figure 1. Two schematic diagrams of the experimental set-up (front and top view).
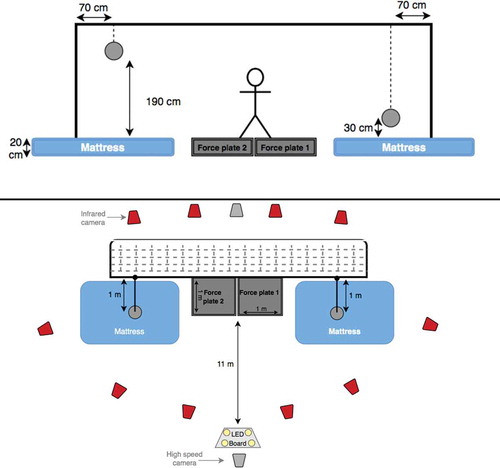
Two custom-made strain-gauge based, 1 × 1 m, force plates (Vrije Universiteit Amsterdam, Amsterdam, The Netherlands) were used to measure ground reaction forces (GRF) produced by each leg separately at a rate of 1000 samples/s. Each force plate was separately covered by artificial football grass, to prevent any force transfer between force plates. A mattress was placed beside each force plate, to avoid unevenness of the floor level, and to keep the goalkeeper and the marker set-up safe at each landing. We believe that the presence of mattresses at landing areas did not affect the performance of the dive, instead it made the goalkeeper more comfortable with the experimental set-up when diving with the markers attached. Two Basler high-speed video cameras (50 Hz) were used to record all trials in the frontal plane for visual checks.
Due to the fact that the diving save is an explosive movement with a large range of motion, gaps in the kinematic data occurred occasionally when markers were out of sight. These gaps were interpolated using a cubic spline function, for a maximum gap length of five samples. Subsequently, a bi-directional second order low-pass Butterworth filter with a cut-off frequency of 12 Hz was used, to smooth the kinematic and kinetic data. The optimal cut-off frequency was estimated on kinematic data using the equation developed by Yu, Gabriel, Noble, and An (Citation1999).
Data analysis
All kinematic and kinetic analysis were carried out using custom software in MATLAB (R2015b, MathWorks inc., United States of America). Six time instants were identified for the diving save: Light signal, dive onset, contralateral peak force (CPF), ipsilateral peak force (IPF), take-off and ball contact.
Detection of dive onset
For detecting the onset of the dive movement, we used an algorithm based on the Approximated Generalized Likelihood-ratio (AGLR) (Staude & Wolf, Citation1999). Neither this algorithm, nor any other algorithm, has been previously used for onset detection in the diving save. Therefore, a wide scope of data inputs and thresholds were tested to establish a method to estimate onset time as accurate as possible while preventing any bias. AGLR works by (1) detecting the alarm time (the time instant when the signal reaches the pre-set threshold) using a sliding test window, then (2) tracking back the signal to detect the initial change time using Maximum Likelihood techniques (Poor, Citation1988). Eventually, we used a threshold of 20% bodyweight and three different data inputs: (1) Total horizontal GRF, (2) Total vertical GRF and (3) Vertical GRF of the contralateral leg (the leg opposite to the diving side). Different data inputs had to be considered, because the goalkeeper can start the dive by choosing from or mixing these three different strategies: (1) Exerting horizontal forces for horizontal displacement towards the ball, (2) Exerting vertical forces for a pre-push off jump and (3) Exerting vertical forces with the contralateral leg for stepping sideward with the ipsilateral leg towards the ball. While it may seem better to choose the earliest onset from these inputs, sometimes the onset detected from one of the data inputs was slightly too early, due to small movements in the starting position before initiating the dive. Therefore, for each individual dive, the final dive onset was defined as the average of the two out of three onsets, having the smallest mutual difference. Subsequently, total time was split into reaction time (from light to dive onset) and dive time (from dive onset to ball contact).
Kinematics
Timing variables were defined relative to the onset of movement. The instant of contralateral peak force (CPF) was defined as the instant that the contralateral leg exerted its maximum resultant GRF. The instant of ipsilateral peak force (IPF) was defined as the instant that the ipsilateral leg exerted its maximum resultant GRF. Take-off was defined as the instant that the vertical component of GRF, summed over legs, dropped below 10% bodyweight threshold and ball contact as the instant that a shift in position of the ball’s markers was detected in any direction.
The characteristics of the starting position (the body posture at the instant of light signal) included stance width (SW), knee joint flexion angle, hip joint flexion angle and forward centre of pressure (CoP) position. The SW was calculated as the distance between the medial malleoli and was expressed as a percentage of the participant’s leg length. The leg length of each goalkeeper was measured from the palpated greater trochanter to the ground while the subject was standing bare feet. The knee and hip joint angles were defined as the Euler angles of the shank anatomical coordinate system (ACS) relative to the thigh ACS, and of the thigh ACS relative to the pelvis ACS, respectively. Their sequence of rotation was: flexion-extension, external-internal rotation and abduction-adduction (Wu et al., Citation2002). The forward CoP was calculated relative to the calcaneus and as a percentage of the participant’s foot length (calcaneus to CoP/calcaneus to tip-toe).
Total body linear momentum in the vertical and horizontal direction towards the ball, and frontal plane angular momentum time series were calculated (Yeadon, Citation1990) from the light signal to ball contact.
Kinetics
The body CoM velocity resulting from the push-off of each of the legs was calculated based on equation 1, which is derived from Newton’s second law of motion:
where is the velocity [m· s−1] and the subscript
represents the leg side (right or left leg).
and
indicate light and take-off instant respectively.
indicates GRF [N] of the leg concerned in the calculation,
is the bodyweight [kg],
is the gravitational acceleration [m· s−2] and
is the sample rate [s−1]. The acceleration resulting from each leg’s push-off was quantified as:
Statistical analysis
All time series were time-normalized from the light signal until ball contact. All data are presented as mean ± standard deviation between-subject. Peak magnitudes, and absolute values at CPF and IPF of the time series were compared between heights and sides, and between heights, and contra- and ipsilateral legs, with two-way repeated measures analysis of variance (ANOVA). The level of significance was set at p < 0.05 and the effect size measure partial eta-squared was reported (0.01 small, 0.06 medium, 0.14 large). All statistical analyses were carried out using IBM SPSS Statistics 20.
Results
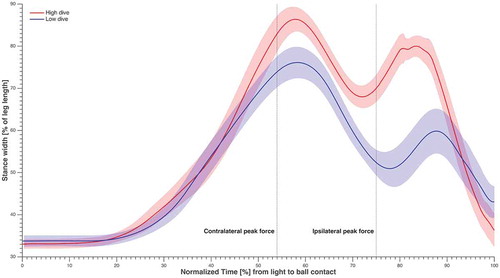
Our results showed that the starting position of the goalkeepers was characterized by a SW of 33 ± 1% of leg length, 62 ± 18° knee flexion and 63 ± 18° hip flexion. The small standard deviation for SW at the starting position, might be caused by similarities in the coaching systems at the goalkeepers’ clubs. The forward CoP was on average located at 75 ± 12% of foot length from the heel. After the light signal, the goalkeepers stepped sideways with their ipsilateral leg to increase the SW from 33 ± 1% to maximum values of 88 ± 8% and 77 ± 11%, for high and low dives respectively (). Repeated measures ANOVAs showed that there was a significant effect of ball height (p < 0.01) on all variables except reaction time, and all effect sizes were large (). Whereas there were no effects of side on any of the variables and no interaction effect. Therefore, the figures in the remainder of the paper show the data averaged over sides to allow for a clear comparison between heights.
Table 1. Mean values with standard deviations for total time, reaction time, dive time, and kinematic and kinetic data (** p < 0.01). Mean differences with effect sizes are also shown.
Figure 2. The time series of stance width averaged over subjects and sides, normalised over time from light to ball contact, and the standard error in shading. The time instants corresponding to maximum GRF for the contralateral and ipsilateral leg were highlighted.
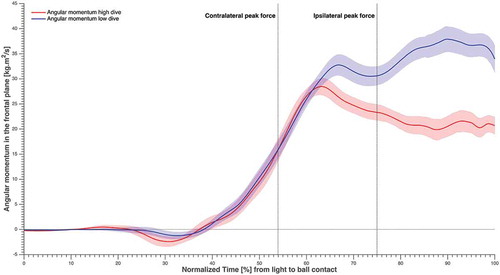
Peak resultant GRF was larger for the ipsilateral leg than for the contralateral leg in high dives, but the opposite was true in low dives (). Horizontal linear momentum was significantly greater (p < 0.01) for low dives than for high dives, at IPF and at the instant that the peak value was attained (; ). The vertical linear momentum was significantly greater (p < 0.01) for high dives than for low dives, at IPF and at the instant that the peak value was attained. In addition, the angular momentum in the frontal plane was significantly greater (p < 0.01) for low dives than for high dives, at IPF and at the instant that the peak value was attained (; ). The push-off angle formed by the contralateral leg GRF vector and CoP-ball vector at CPF was 34 ± 3° for high dives and 54 ± 4° for low dives, whereas at IPF the push-off angle formed by the ipsilateral leg GRF vector and CoP-ball vector was 41 ± 3° for high dives and 60 ± 4° for low dives.
Figure 3. The resultant GRF of each leg averaged over subjects and sides, in solid and dashed lines (refer to the figure legend), normalised over time from light to ball contact, and the standard error in shading. The time instants corresponding to maximum GRF for the contralateral and ipsilateral leg were highlighted.
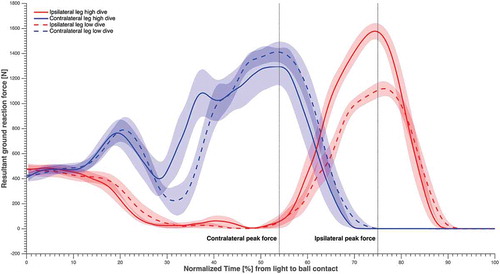
Figure 4. Total body linear momentum averaged over subjects and sides, in solid and dashed lines (refer to the figure legend), normalised over time from light to ball contact, and the standard error in shading. The time instants corresponding to maximum GRF for the contralateral and ipsilateral leg were highlighted.
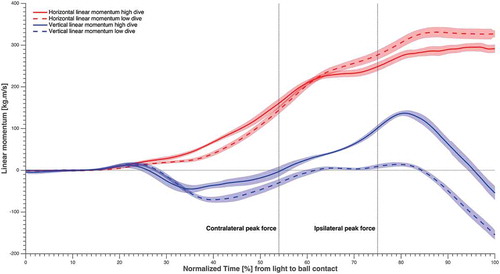
Figure 5. Total body angular momentum in the frontal plane averaged over subjects and sides, normalised over time from light to ball contact, and the standard error in shading. The time instants corresponding to maximum GRF for the contralateral and ipsilateral leg were highlighted.
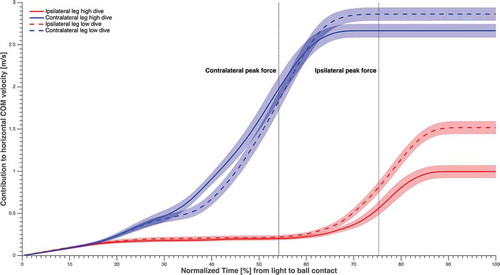
The maximum contribution to the COM horizontal as well as the vertical velocity towards the ball was larger (p < 0.01) for the contralateral leg (2.7 ± 0.1 m·s−1 and 3.1 ± 0.3 m·s−1, respectively) than for the ipsilateral leg (1.2 ± 0.1 m·s−1 and 0.4 ± 0.2 m·s−1, respectively), during all diving save conditions ( and ). Furthermore, maximum contralateral leg contribution to vertical COM velocity was significantly larger for high dives than low dives (p < 0.01), whereas the maximum contralateral leg contribution to the horizontal COM velocity was significantly larger for low dives than for high dives (p < 0.01).
Figure 6. The contribution of each leg to vertical COM velocity averaged over subjects and sides, in solid and dashed lines (refer to the figure legend), normalised over time from light to ball contact, and the standard error in shading. The time instants corresponding to maximum GRF for the contralateral and ipsilateral leg were highlighted.
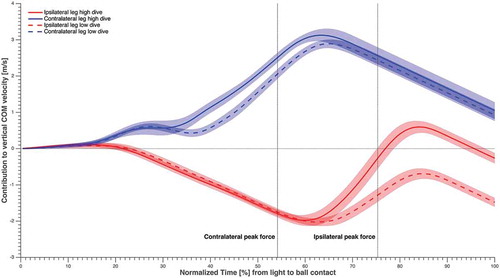
Figure 7. The contribution of each leg to horizontal COM velocity averaged over subjects and sides, in solid and dashed lines (refer to the figure legend), normalised over time from light to ball contact, and the standard error in shading. The time instants corresponding to maximum GRF for the contralateral and ipsilateral leg were highlighted.
Discussion
This study describes the mechanics of the diving save in elite goalkeepers towards high and low balls, at both sides of their body. Overall, findings were that subjects stepped sideward with the ipsilateral leg at initiation of the diving save. In accordance with our second hypothesis, angular momenta were larger in low dives. Peak linear momenta were larger in vertical direction for high dives and in horizontal direction for low dives. In contrast to our first hypothesis, the contralateral leg contributed more than the ipsilateral leg to COM velocity. There were no significant differences between diving towards the right versus left side on any of the outcome variables (). The latter contradicts the findings of Spratford et al. (Citation2009) on goalkeepers from the Australian youth team. The high level of the goalkeepers participating in the current study may explain the absence of an effect of diving side on any outcome variable. It is clear from the present study that the diving save is a bilateral task involving both lower limbs in explosive push-offs. In addition, placing the ball slightly in front of the goal and using two force plates instead of only one, may have given the goalkeeper more freedom to perform without constraints. The experimental set-up of the current study took into consideration many factors to realistically simulate a game-situation diving save, but one limitation was still that we had the balls hanging in a stationary position.
This study was the first to use automated detection of dive onset. The average calculated reaction time was 211 ± 25 ms, and this is in accordance with previous literature on perception and anticipation during penalty kicks (Savelsbergh et al., Citation2002, Citation2005). There was no significant effect of height or side on reaction times, in spite of the differences in force production between these conditions, indicating that the algorithm was not biased by differences in movement strategy.
and showed that contralateral push-off was essential, as the contralateral leg contributed significantly more than the ipsilateral leg to the COM velocity towards the ball. This was in contrast with our first hypothesis, which was based on training recommendations and the emphasis of goalkeeper’s technical, and strength and conditioning coaches. The reason behind this might be the fact that the contact time of contralateral push-off (480 ± 71 ms) was significantly longer (p < 0.01) than ipsilateral push-off (400 ± 40 ms). Furthermore, the ipsilateral leg was mainly used at the beginning of the dive to increase the SW of the goalkeepers (), and it could not start contributing until it was back on the ground and able to apply force (). The maximum SW values reached corresponded with the moment that the ipsilateral foot was back on the ground and ready to start contributing positively to the COM velocity.
The relatively wide SW reached before IPF might be beneficial for putting the contralateral leg in a better position for push-off and putting the body in a better position for generation of angular momentum. The push-off angles formed by GRF vectors of each leg with the CoP-ball vector at CPF and IPF separately, showed a better alignment for the contralateral leg when compared to the ipsilateral leg. The GRF vector of the ipsilateral leg was more upright and therefore more suitable for vertical linear momentum, which was indeed mainly generated near IPF (). However, using the ipsilateral leg to increase SW at the beginning of the dive implied a shorter contact time for the ipsilateral push-off and caused a negative contribution to COM vertical velocity () before the ipsilateral leg was back to the ground and ready to contribute positively again. So, despite a smaller overall contribution to vertical COM velocity compared to the contralateral leg, the large and rapid increase of the ipsilateral contribution to vertical COM velocity at IPF () indicates that the ipsilateral push-off was still essential to obtain vertical momentum in the high dives. Future research could analyse the diving save performance while attempting to change the SW at the starting position. Starting from a SW closer to the maximum values that we found after the sidestep, may reduce the length of the required side step and allow the ipsilateral leg to contribute positively for a longer time.
Low dives were characterised by greater horizontal linear momentum and frontal plane angular momentum, while high dives were characterised by greater vertical linear momentum. This is partially in accordance with our second hypothesis, as horizontal linear momentum appeared to be essential for both high and low dives. Overall, the horizontal linear momentum was significantly greater (p < 0.01) than the generated vertical linear momentum at all time events. This is evident given the fact that the horizontal space of the goal to be covered by the goalkeeper, is notably beyond the vertical one. Both dive heights required a substantial angular momentum in order to orient the body horizontally and therefore increasing the horizontal reach. Contralateral leg abduction while sidestepping with the ipsilateral leg, to increase the SW, may have been beneficial to generate the angular momentum, as the latter greatly increased around CPF ().
The current goalkeeping strength and conditioning programs focus mainly on vertical strength and power movement patterns (e.g. squat exercise, squat jumps, countermovement vertical jumps). Instead, it would be beneficial if strength and conditioning coaches focus on two main findings of this study: the strong contralateral leg contribution to total COM velocity, and the greater horizontal linear momentum. Thus, emphasizing lateral movement patterns in the frontal plane driven by the contralateral leg (e.g. side push-offs, lateral jumps, side lunge exercise) for physical performance testing and training, instead of the vertical movement patterns in the sagittal plane. This would be advantageous, as the two skills are relatively independent of one another and represent different leg strength/power abilities (Meylan et al., Citation2009). Future studies can give more insight into joint power and work to give comprehensive directions for coaches regarding isolated and coordinated joint movement training for goalkeepers.
Conclusion
In conclusion, goalkeepers start the dive by pushing-off with their contralateral leg, while increasing the SW by sidestepping with the ipsilateral leg towards the ball. Once the maximal SW is reached, the contralateral GRF tends to fall and the ipsilateral GRF starts to build-up and take over the dive. Strength and conditioning coaches need to highlight horizontal lateral skills, to both sides of the body, with emphasis on the push-off with the contralateral leg, for effective assessment and training.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Barlett, R. M. (1999). Sports biomechanics: Reducing injury and improving performance. London: E & FN Spon.
- Faber, G. S., Chang, C., Kingma, I., & Dennerlein, J. (2013). Lifting style and participant’s sex do not affect optimal inertial sensor location for ambulatory assessment of trunk inclination. Journal of Biomechanics, 46, 1027–1030.
- Faber, G. S., Kingma, I., & van Dieën, J. H. (2011). Effect of initial horizontal object position on peak L5/S1 moments in manual lifting is dependent on task type and familiarity with alternative lifting strategies. Ergonomics, 54, 72–81.
- Franks, I. M., & Hanvey, T. (1997). Cues for goalkeepers. High-tech methods used to measure penalty shot response. Soccer Journal, 42, 30–33.
- Ibrahim, R., Faber, G. S., Kingma, I., & van Dieën, J. H. (2016). Kinematic analysis of the drag flick in field hockey. Sports Biomechanics, 16(1), 45–57.
- Kingma, I., de Looze, P. M., Toussaint, M. H., Klijnsma, G. H., & Bruijnen, B. M. T. (1996). Validation of a full body 3-D dynamic linked segment model. Human Movement Science, 15, 833–860.
- Kuhn, W. (1988). Penalty kick strategies for shooters and goalkeepers. In T. Reilly, A. Lees, K. Davids, & W. Murphy (Eds.), Science and football (pp. 489–492). London: E & FN Spon.
- Meylan, C., McMaster, T., Cronin, J., Mohammad, N., Rogers, C., & deKlerk, M. (2009). Single-leg lateral, horizontal, and vertical jump assessment: Reliability, interrelationships, and ability to predict sprint and change-of-direction performance. Journal of Strength and Conditioning Research, 23(4), 1140–1147.
- Poor, H. V. (1988). An introduction to signal detection and estimation (pp. 173–185). New York, NY: Heidelberg, Springer.
- Powers, S. K., & Howley, E. T. (2007). Exercise physiology, theory and application to fitness and performance (6th ed.). New York, NY: McGraw Hill.
- Savelsbergh, G. J. P., Williams, A. M., Van der Kamp, J., & Ward, P. (2002). Visual search, anticipation and expertise in soccer goalkeepers. Journal of Sports Sciences, 20, 279–287.
- Savelsbergh, G. J. P., Williams, A. M., Van der Kamp, J., & Ward, P. (2005). Anticipation and visual search behavior in expert soccer goalkeepers. Ergonomics, 48, 11–14, 1686–1697.
- Spratford, W., Mellifont, R., & Burkett, B. (2009). The influence of dive direction on the movement characteristics for elite football goalkeepers. Sports Biomechanics, 8, 235–245.
- Staude, G., & Wolf, W. (1999). Objective motor response onset detection in surface myoelectric signals. Medical Engineering & Physics, 21, 449–467.
- Suzuki, S., Togari, H., Isokawa, M., Ohashi, J., & Ohgushi, T. (1987). Analysis of the goalkeeper’s diving motion. In T. Reilly (Ed.), Proceedings of the First World Congress of Science and Football (pp. 468–475). Liverpool: E&F Spon.
- Williams, A. M. (2000). Perceptual skill in soccer: Implications for talent identification and development. Journal of Sports Sciences, 18, 1–14.
- Williams, A. M., Davids, K., & Williams, J. G. (1999). Visual perception and action in sport. London: E & FN Spon.
- Wu, G., Siegler, S., Allard, P., Kirtley, C., Leardini, A., Rosenbaum, D., … Stokes, I. (2002). ISB recommendation on definitions of joint coordinate system of various joints for the reporting of human joint motion, Part I: Ankle, hip and spine. Journal of Biomechanics, 35, 543–548.
- Yeadon, M. R. (1990). The simulation of aerial movement – III. The determination of the angular momentum of human body. Journal of Biomechanics, 23(1), 75–83.
- Yu, B., Gabriel, D., Noble, L., & An, K. (1999). Estimate of the optimum cutoff frequency for the butterworth low-pass digital filter. Journal of Applied Biomechanics, 15, 318–329.