
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
This study examined the processes of perceptual-motor calibration/recalibration of defensive football players in a 1-vs-1 scenario. Ankle weights were used to reduce the acceleration capabilities of players performing an anticipation test, with the aim being to examine the player’s response to the disturbance in terms of when movement was initiated and the impact on the mechanisms that underpinned anticipation, namely gaze behaviour. The ankle weights disturbed the perceptual-motor system and players initiated movement significantly earlier in the 1-vs-1 anticipation test. Analyses of perceptual-motor calibration/recalibration revealed that players acted closer to their maximal action capabilities prior to the addition of ankle weights, which negatively influenced the scaling of action capabilities. Moreover, players were unable to recalibrate whilst wearing ankle weights. However, following the withdrawal of the ankle weights, players were able to recalibrate within 11–15 trials. Players did not adapt gaze behaviour as a result of the disturbance being placed on the perceptual-motor system, but task familiarization resulted in more efficient eye movements. The results of this study show the importance of providing players the opportunity to “scale” action to perceptual information.
1. Introduction
The affordance-based control (Fajen et al., Citation2009) account of anticipation in sport proposes that the perception of opportunities for action (affordances) is scaled to an individual’s action capabilities (van der Kamp et al., Citation2018). Specifically, in team sports such as football, if calibrated, a player’s capabilities will guide what opportunities for action are available relative to the immediate environmental situation. The strong dependence that athlete–environment interactions have on the capabilities of sportspeople to act has been supported empirically by studying skills in boxing (Hristovski et al., Citation2006), football (Wilson et al., Citation2019), tennis (Mecheri et al., Citation2019) and parkour (Croft & Bertram, Citation2017). While action capabilities may shape the field of affordances available to a sportsperson (Rietveld & Kiverstein, Citation2014), these affordances still need to be picked up by the athlete. For this, attunement (i.e., exploiting the information specifying an affordance) and calibration (i.e., finding the appropriate scaling between information and action) are crucial. That is, performance differences between skilled and less-skilled sportspeople can, in some part, be attributed to variations in the informational variables that are exploited in the control of actions (Araujo et al., Citation2006; Passos et al., Citation2008). Following an ecological approach, movement provides learners with the opportunity to both detect and discover information (Gibson, Citation1979). A player may be perceptually attuned and have the capability to act, but successful performance requires players to then “scale” the action to perceptual information and vice-versa (Withagen & Michaels, Citation2004). In affordance-based control theory, this “scaling” is termed perceptual-motor calibration (Fajen et al., Citation2009).
Calibration can be considered “structural” and describes the scaling of actions to the environment (Fajen et al., Citation2009). For example, years of practice will enable football players to become sensitive to their own action capabilities; experience offers a knowing of what they “can” and “cannot” do (Fajen et al., Citation2009). However, the calibration can also be more functional and task specific (Brand & de Oliveira, Citation2017). Whilst it has been suggested that long-term experience will generally calibrate a sportsperson to their performance environment (Fajen et al., Citation2009), they will still be more or less calibrated in certain specific situations (Brand & de Oliveira, Citation2017). On the other hand, recalibration happens after an acute disturbance in either perception or action, rendering the current perception–action relationship inaccurate (Brand & de Oliveira, Citation2017). For example, in football, when a player’s acceleration capabilities are compromised by fatigue, recalibration might enable a player to cope, relatively quickly, with the acute change (Brand & de Oliveira, Citation2017). Using the example of defensive 1-vs-1 situations in football, Wilson et al. (Citation2021) found that recalibration following a significant reduction in acceleration speed resulted in defenders moving earlier to allow more time to intercept an attacker’s run. It has been reported that recalibration occurs rapidly when there is a good match between the action that requires recalibration and the movements that players make during exploration (van Andel et al., Citation2017). In sum, calibration requires experience and practice to scale the perception–action relationship to general or more specific changes to the environment (Seifert et al., Citation2021), whereas recalibration is a process that enables performers to deal with more acute disturbances to the perceptual-motor system (e.g., fatigue).
In the sport domain, the concepts of calibration and recalibration have exclusively been explored by way of changes in sporting equipment. For example, Scott and Gray (Citation2010) explored perceptual-motor control adjustments in response to changes in baseball bat weights when facing simulated pitches. Batters switched from a standard bat to lighter or heavier bats. The switch to either the lighter or the heavier bats caused significant increases in swing timing errors; however, batters recalibrated quickly within 5–10 trials. Nakamoto et al. (Citation2012) investigated the aftereffects of the use of a heavy-weighted bat in a batting warm-up. Eight college baseball players performed three warm-ups before a simulated batting task: a normal warm-up with a standard bat (0.85 kg); a recalibrated warm-up with a weighted bat (1.2 kg) followed by swings with a standard bat (0.85 kg); and a weighted warm-up with a 1.2 kg bat. When the batters needed to correct their swings to changes in ball velocity, larger timing errors were produced in the weighted than the normal warm-up condition. Thus, results indicate that in the context of baseball batting, which has severe spatiotemporal constraints, warm-ups with a weighted bat create adverse effects for perceptual-motor calibration. Whilst the research on perceptual-motor calibration and recalibration from a sport equipment perspective is compelling, limited research has aimed to identify and assess the mechanisms that underpin the recalibration process.
Wilson et al. (Citation2021) reported that fatigue induced through small-sided football games resulted in a reduction in action capabilities and defending players tended to move earlier when they were prospectively anticipating the change of direction of an attacking player. This supported previous studies that found action capabilities directly influence anticipation (Brault et al., Citation2012; Dicks et al., Citation2010a; Mecheri et al., Citation2019; Wilson et al., Citation2019). Despite the importance placed on action capabilities, players can only safely and accurately perform near the limits of their action capabilities if they are attuned and calibrated (Fajen et al., Citation2009). Fajen et al. (Citation2009) proposed that the perception of affordances is influenced by perceptual attunement, which refers to an ability to adapt and attend to different information. A defender in a 1-vs-1 situation in football will use vision (Alder et al., Citation2016) and action (van der Kamp et al., Citation2008) to exploit kinematic informational variables of their opponent. However, it has been proposed that perceptual attunement alone may not be sufficient for accurate performance because calibration – the scaling of action to perceptual information – is also necessary (Withagen & Michaels, Citation2004). For example, from the defensive standpoint of the 1-vs-1 situation in football, defenders will need to attune to certain informational variables and subsequently, scale perception-action to these informational variables.
Gaze patterns offer a means to make inferences as to “what” information is exploited (Alder et al., Citation2014; Roca et al., Citation2013). It has been reported that during the penalty kick in football, gaze behaviours – such as the location and timing of fixations – vary markedly between goalkeepers early in the run-up, whereas gaze variation decreases at around 250 ms before ball contact (Navia et al., Citation2017; Ramsey et al., Citation2020). One might expect that with practice, goalkeepers would be more likely to converge towards more informative areas (e.g., about the lower leg), as opposed to information (e.g., torso movement) that occurs earlier in the kinematic chain. However, the information sources that players exploit during a 1-vs-1 defensive situation are not known. Furthermore, little is known about the process of perceptual-motor recalibration when changes in action capabilities render the perception–action relationship inaccurate. A novel means to address this current shortcoming in understanding is through the examination of in-situ gaze behaviour using mobile eye-tracking systems. When players perceive their direct opponents’ upcoming actions, an analysis of eye movements can identify the gaze patterns that players use, and inferences can be made about attunement to information (Abernethy et al., Citation2012). Many gaze behaviour studies have focussed analysis on fixations (Alder et al., Citation2014; Williams & Davids, Citation1998), which are measures of an absence of eye movement; that is, where players are orienting their gaze (de Oliveira et al., Citation2009). Crucially, for the purposes of this study, the analysis of gaze behaviour offers a means to deepen the current understanding of the processes underpinning perceptual-motor calibration and recalibration.
The purpose of the current study was to examine the processes of perceptual-motor calibration and recalibration of defensive football players in a 1-vs-1 anticipation test. First, an extended period of task familiarization trials was offered to players to allow them to scale action capabilities to the perception-action task (i.e., calibrate). Second, following evidence (e.g., Wilson et al., Citation2021) that acceleration capabilities influence the anticipatory actions of athletes in 1-vs-1 anticipation situations, ankle weights were used as an acute and targeted manipulation of acceleration capabilities (see Ramenzoni et al., Citation2008) to measure recalibration. To examine the processes of perceptual-motor calibration/recalibration in the context of changes to action capabilities, the current study implemented the framework proposed by Brand and de Oliveira (Citation2017), which is to measure performance at 1) Baseline – action without constraining the motor system; 2) Disturbance – action with the motor system constrained; and 3) Removal – action without constraining the motor system.
It was hypothesized that during the baseline phase, calibration would be an iterative process, and players would be better calibrated to the task with exploration (i.e., number of trials prior to disturbance). Secondly, a reduction in 5-m acceleration capabilities would disturb the perceptual-motor system, and players would move earlier in the 1-vs-1 anticipation test as a result of recalibration (Wilson et al., Citation2021). In addition to the study of movement times as a feature of (re)calibration, we included the novel measurement of eye movements to further understand the calibration process. Given the lack of existing research examining eye movements in the context of calibration, we conducted exploratory analysis of changes in gaze patterns, informed by extant measures in the anticipation literature (e.g., location and duration of final fixation: Alder et al., Citation2014). Based on these measures, our exploratory hypotheses were that players would contend with the disturbance placed on the perceptual-motor system by an adaptation in gaze patterns. Specifically, it was expected that when the ankle weights were applied (disturbance), players would move earlier and, therefore, fixate on the earlier unfolding kinematic information of the tester (e.g., head, torso and hips), resulting in an earlier onset of final fixation. Finally, it was hypothesized that perceptual-motor recalibration would be required when the disturbance (i.e., ankle weights) was applied and withdrawn, and in line with the baseline phase, this would be an iterative process, with players being better recalibrated to the task with exploration (i.e., number of trials in each respective phase) (Brand & de Oliveira, Citation2017).
2. Methods
2.1. Participants
Male (n = 17) college and university outfield football players (mean ± SD: age: 19.1 ± 1.2 years, mass: 72.3 ± 5.8 kg, height: 1.77 ± 0.08 m) took part as participants in the study. One male college outfield player (age: 19 years, mass 75.2 kg, height: 1.82 m) was recruited as the sole tester in the 1-vs-1 anticipation test. All participants, including the tester, averaged over 11 years of playing experience (mean ± SD: 11.7 ± 2.8 years). The local university ethics board granted ethical approval (LB62935).
2.2. 1-vs-1 anticipation test
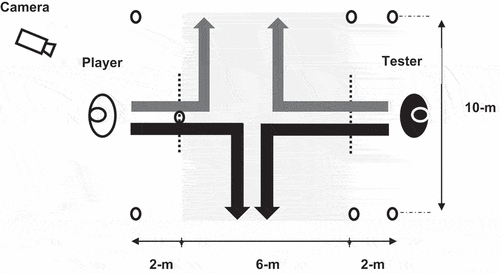
Anticipation was tested via a 1-vs-1 anticipation test, which was adapted from a basketball anticipation test, which has acceptable test–retest reliability (ICC = 0.89–0.99, Scanlan et al., Citation2014). The test mimicked the typical actions required by defenders in football during corners, throw-ins and free-kick situations. Players responded to the movement initiation and directional changes of a tester. Players were required to move towards the tester and then match the direction taken by the tester (attacker without ball). Once the direction change had been initiated, both the tester and the player were asked to race to the end, which was 5-m in either direction, see . The tester was instructed by the lead investigator as to the required direction change prior to each trial and was asked to initiate the change in direction within a 6-m zone. The tester was not permitted to purposely deceive the player and was only permitted to initiate a change of direction by planting their outside foot. That is, if the tester were changing direction to their right, they could only initiate a change of direction by planting their left foot (Scanlan et al., Citation2014). All trials were recorded on a high-speed camera (Fastec TS3, California, USA) at a frame rate of 240 Hz.
Figure 1. 1-vs-1 anticipation test.
2.3. Pre-planned acceleration test
An acute manipulation of acceleration capabilities was required at the disturbance phase, with the aim being to significantly reduce the players acceleration capabilities but not in a way that completely diminished their chances of being successful in the 1-vs-1 anticipation test. Therefore, players (n = 16) took part in a pre-planned acceleration test. The test required the players to start 1.5-m in front of timing gates (Brower System, USA) and then make a 90°Change of Direction (COD) to the test end point, 5-m away. The players completed six trials (COD to the left for three trials; COD to the right for three trials) in unweighted and weighted conditions. T-tests confirmed that players were significantly slower in the COD test when wearing 450 g ankle weights (unweighted = 1110 ± .100 ms, weighted = 1170 ± .70 ms, t (31) = −4.43, p < .02, d = 0.66). Overall, an average 6% performance reduction was observed, and it was determined that 450 g ankle weights were an appropriate reduction of acceleration capabilities.
2.4. Procedure
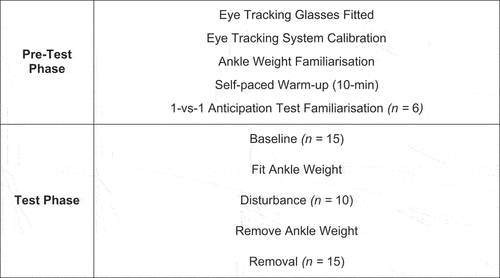
This study employed a within-subject test design. Testing procedures for each player were carried out in one data collection visit and testing took place on grass and 3 G football pitches. An overview of the experimental procedure is presented in . Prior to participation, players were explained the experimental process in detail and completed the participant consent form. The eye-tracking glasses were then fitted to the player’s head (Tobii Pro 2, Stockholm, Sweden), and a practice calibration of the eye tracker was carried out. Following this, a test of the fitting and unfitting of the 450 g ankle weights was conducted, which were attached by two Velcro straps just above the ankle and fitted so there was space between the ankle strap and the player’s football boots. The players then carried out a self-paced warm-up of approximately 10 minutes and then underwent a process of 1-vs-1 anticipation test familiarization, which consisted of six randomized left or right trials. As required by the manufacturer (Tobii Pro 2, Stockholm, Sweden), eye tracking calibration consisted of players fixating on a pre-determined target location point that was 1.5 m from their standing position, and calibration was repeated prior to each trial. Furthermore, the players were exposed to this process during 1-vs-1 anticipation test familiarization. All players then completed forty 1-vs-1 anticipation test trials, which were broken down into three distinct phases (Brand & de Oliveira, Citation2017) that were always completed in the same order: 1) baseline; 2) disturbance; and 3) removal. The baseline phase required players to complete fifteen 1-vs-1 anticipation test trials. Prior to the disturbance phase, a 450 g ankle weight was attached to each of the player’s legs; players then completed ten 1-vs-1 anticipation test trials. The ankle weights were removed before players completed the final fifteen 1-vs-1 anticipation test trials in the removal phase, see Scott and Gray (Citation2010), and more recently, Brand and de Oliveira (Citation2023), for trial number rationale.
Figure 2. Timeline of experimental protocol.
2.5. 1-vs-1 anticipation test dependent variables
Movement Time: Movement time was determined as the time interval from the first identifiable outside foot contact initiating directional change of the tester to the first identifiable foot contact initiating the response of the player (Gabbett et al., Citation2008). That is, if the tester was turning left, it was initiated by a cutting movement of the right foot. In this case, the timing and placement of the right foot was used to determine movement time and vice versa for turning right. A positive value was recorded if movement time occurred after the first identifiable foot contact of the tester, and negative value was recorded if movement time occurred before the first identifiable foot contact of the tester.
Tester Trial Time and Player Trial Time: Tester trial time was determined as the time interval from the first identifiable foot contact initiating directional change of the tester to the first frame confirming that a part of the tester’s body had broken the line of the endpoint marker. Player trial time was determined as the time interval from the first identifiable foot contact initiating the response of the player to the first frame confirming that a part of the player’s body had broken the line of the endpoint marker.
Action-Scaled Time: Based on the assumption that athletes can be sensitive to their own action capabilities (see van der Kamp et al., Citation2018), beating the tester to the endpoint in the 1-vs-1 anticipation test requires that movement time is scaled according to a player’s perception of how fast they, and the tester, can travel to the 5-m endpoint. As such, action-scaled time was equal to (Player trial time + Movement time)/Tester trial time, with a value equal to or less than 1 implying the player was successful (i.e., acted within their maximal action capabilities). For example, if the tester requires 1 s to travel from the change of direction point to the endpoint (tester trial time) and the player (player trial time) requires 1.2 s to travel the same distance, then the player will arrive at the endpoint 0.2 s after the tester if the player initiates a movement response at the same point as the tester plants his outside foot to change direction. In this example, the player has timed their action outside of their maximal action capabilities. For the player to scale within their maximal action capabilities, the player must initiate a response 0.2 s earlier than the tester to arrive at the endpoint at the same time, and more than 0.2 s earlier to arrive at the endpoint before the attacker (see Zheng et al., Citation2022).
2.6. Gaze behaviour
A Tobii Pro 2 mobile eye-tracking system (Stockholm, Sweden) was used to record gaze behaviours. The mobile system is a head-mounted monocular eye-tracking system that computes point of gaze within a scene through the calculation of the vector between the player’s pupil and cornea (Alder et al., Citation2014). The calibration consisted of players fixating on a pre-determined target location point that was approximately 1.5 m from their standing position. Gaze behaviour data were recorded at 25 Hz, and the video footage from the Tobii Pro 2 system was subjected to frame-by-frame analysis using Adobe Premiere Pro CC 2020 (San Jose, USA).
2.7. Gaze behaviour dependent variables
In this study, five gaze behaviours were considered: i) number of fixations; ii) fixation duration; iii) scan ratio; iv) duration of final fixation; and v) location of final fixation (Alder et al., Citation2014). Fixations were defined as gaze remaining at a location for a minimum duration of 120 ms (Alder et al., Citation2014). Fixations, i.e., the absence of eye movement allow for visual perception to take place (Holmqvist et al., Citation2011) and inferences can be made about the player’s attention to the tester’s postural information (Abernethy et al., Citation2012). The number of fixations per trial was counted, and the mean duration per fixation calculated. As a composite measure, the scan ratio was calculated as the sum of fixations per trial divided by the duration of all fixations in that trial. These measures were chosen because they highlight proactive gaze behaviours and would indicate that players sought alternative perceptual information at the disturbance and removal phases, respectively. The final fixation was defined as the fixation that occurred during the time interval from the frame where the tester lifted their outside foot to initiate directional change to the frame where the foot contacted the ground, i.e., if the tester were changing direction to their right, the first frame that the tester planted their left foot (Scanlan et al., Citation2014). Duration of final fixation and location of final fixation are variables of interest because previous findings have signified that goalkeepers converge to similar locations later in a penalty taker’s run-up (Navia et al., Citation2017). Therefore, if the final fixation location and timing of fixation are critical to performance, this might reflect better scaling of actions in the 1-vs-1 anticipation test. Following initial inspection of the gaze data, the final fixation location was categorized as legs, change of direction area (COD-A) and other. The rationale being that “other” comprised of body segments that are proximal to the tester, i.e., head, torso and hips, and the final key postural information likely emanated within the COD-A, which is where the tester makes the first identifiable outside foot contact initiating a directional change. The definition of these locations is provided in and augmented by a visual representation.
Figure 3. Gaze behaviour locations of interest.

2.8. Statistical analysis
Sample Size: Data collection for this study was severely impacted by COVID-19 and the national lockdown imposed by the UK government. A sensitivity test using G*Power software (v.3.0.10) (Faul et al., Citation2007) indicated that the sample size of this study (n = 17) shows a detectable effect size of 0.36, with an alpha of a = 0.05 and a power of β = 0.80.
Tester Trial Time and Player Trial Time Manipulation Check: Shapiro–Wilk tests of normality confirmed that the tester trial time and the player trial time data both met assumptions of parametric tests. Therefore, as a check of the ankle weight manipulation, tester and player differences between baseline, disturbance and removal phases were tested by two separate repeated measures of univariate analysis of variance and follow-up t-tests.
Movement Time: To explore the effect of the disturbance, differences in movement time between phases (i.e., baseline, disturbance, and removal) and between critical transitions were analysed. Critical transitions were defined as the final baseline trial (trial 15) and the first disturbance trial (trial 16) and the final disturbance trial (trial 25) and the first removal trial (trial 26). Shapiro–Wilk tests of normality confirmed that all dependent variables met assumptions of parametric tests. Differences in movement time between phases were tested by a repeated measures univariate analysis of variance and follow-up t-tests. To examine the distinct effect of the disturbance on movement time, paired-samples t-tests were conducted. To test the relationship between player trial time and movement time, the change scores for each critical transition were calculated ( movement time & player trial time for trials 15–16 & for trials 25–26) and subjected to a Pearson’s correlation analysis.
Action-Scaled Time: To analyse the process of perceptual-motor calibration (i.e., in the baseline phase) and recalibration (i.e., impact of the ankle weights at the disturbance and removal phases), action-scaled time was the dependent variable of interest. To examine the process of perceptual-motor calibration, a linear regression analysis was conducted at the baseline, disturbance, and removal phases, whereas for perceptual-motor recalibration, quadratic regression analyses were conducted between the two critical transitions – baseline/disturbance and disturbance/removal. Regression analyses assess the relationship between the dependent variable (i.e., action-scaled time) and the independent variable (i.e., trial number). Following the inspection of action-scaled time scatterplots and using a paired-samples t-test, a general assessment of recalibration was conducted between the last five baseline trials and the last five removal trials. All data are presented as mean ± SD and significance was accepted at p ≤ .05 for all statistical tests. Effect sizes are reported as Cohen’s d, with 95% confidence intervals (CI) of the observed effect calculated. Effect sizes are interpreted according to Cohen’s recommendations; small (0.2), moderate (0.5) and large (0.8).
2.9. Gaze behaviour statistical analysis
Initial inspection of the gaze behaviour data identified that 440/680 trials (65%) were successful in locating point of gaze. The eye tracking system had issues locating the point of gaze in direct sunlight, which resulted in an 11-player sample that consisted of 165 baseline, 110 disturbance and 165 removal trials. Shapiro–Wilk tests of normality confirmed that assumptions for parametric testing were not met for the number of fixations, duration of fixations, scan ratio and duration of final fixation. Therefore, differences in these gaze variables were tested by non-parametric difference tests.
Between Testing Phases: To explore the effect of the disturbance on gaze behaviour, differences between phases (i.e., baseline, disturbance & removal) were tested by using Friedman Test’s. Post-hoc Wilcoxon-Signed Rank tests were conducted for all significant observations. Final fixation location differences between phases were tested by using a chi-square test.
Successful Versus Unsuccessful Trials: To investigate if any gaze behaviour variables could discriminate between successful and unsuccessful trials, trials from across the three testing phases were identified by applying the action-scaled time threshold of ≤1.00 s. In total, there were 90 trials (baseline = 27 trials; disturbance = 19 trials; removal = 44 trials). For comparison, 90 unsuccessful trials (action-scaled time ≥ 1.10) were randomly selected from across the three testing phases. This resulted in an equal number of successful and unsuccessful trials at each testing phase. Wilcoxon-Signed Rank tests were used to check for significant differences in the number of fixations, duration of fixations, scan ratio and duration of final fixation between successful and unsuccessful trials. Final fixation location differences between successful and unsuccessful trials were examined by using a chi-square test.
Changes in Gaze Behaviour Across Testing Phases: To further investigate perceptual-motor calibration, linear regression analyses were conducted at the baseline phase for number of fixations, fixation duration, scan ratio and duration of final fixation, whereas for perceptual-motor recalibration, quadratic regression analyses were conducted between the two critical transitions – baseline/disturbance and disturbance/removal – to assess the predictivity of trial number for each of the gaze behaviour outcome variables (i.e., number of fixations, fixation duration, scan ratio, duration of final fixation). Ordinal linear and quadratic regression analyses were conducted for the final fixation location. All visual search data are presented as median (Mdn) and significance was accepted at p ≤ .05 for all statistical tests. Effect sizes are reported as Cohen’s d, which were converted from Pearson’s correlation coefficient (r), and 95% confidence intervals (CI) of the observed effect were calculated. Effect sizes are interpreted according to Cohen’s recommendations; small (0.2), moderate (0.5) and large (0.8).
3. Results
3.1. 1-vs-1 anticipation test
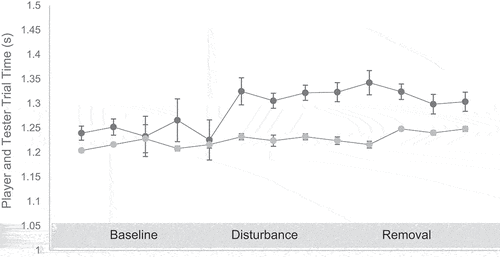
Player Trial Time and Tester Trial Time Manipulation Check: A one-way ANOVA indicated that player trial time was significantly different between baseline, disturbance and removal testing phases (F (2,660) = 17.65 p ˂ .01). Follow-up t-tests revealed that players were significantly slower in the disturbance phase compared to the baseline phase (t(163) = −2.45, p = .001, d = 0.52 95% CI [.25, .28]), and players were significantly slower at the removal phase compared to the baseline phase (t(241) = −5.37, p = .001, d = 0.46, 95% CI [.43, .46]), see . There was no significant difference in player trial time between the disturbance phase and the removal phase (t(158) = 0.89, p = .12, d = 0.07, 95% CI [−.01, .01]), respectively. Furthermore, tester trial time was significantly different between baseline, disturbance and removal testing phases (F (2,660) = 3.33 p = .03). However, follow-up t-tests revealed that the tester was only significantly slower at removal phase compared to the baseline phase (t(241) = −2.24, p = .02, d = 0.23, 95% CI [.22, .24]) and there were no significant differences in tester trial time between baseline and disturbance (t(164) = −0.84, p = .40, d = 0.10, 95% CI [−.12, .14]), and disturbance and removal (t(159) = 0.53, p = .12, d = 0.06, 95% CI [.10, .14]), respectively (see ).
Figure 4. Player trial time and tester trial time across the three testing phases. Player (black) and tester (Grey).
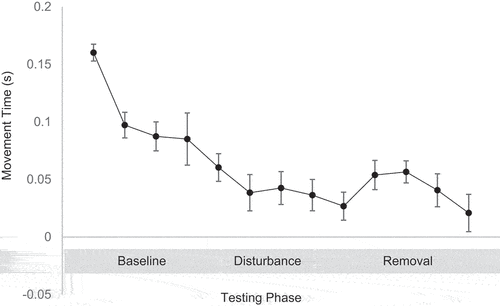
Movement Time: A one-way ANOVA indicated that the movement time was significantly different between baseline, disturbance and removal testing phases (F (2,660) = 19.01, p = .001). Follow-up t-tests revealed that players initiated a significantly earlier movement at the disturbance phase compared to the baseline phase (t(163) = 3.17, p = .002, d = 0.35, 95% CI [.13, .56]) and players initiated a significantly earlier movement at the removal phase compared to the baseline phase (t(241) = 6.13, p = .001, d = 0.55, 95% CI [.37, .74]). There was no significant difference in movement time between the disturbance phase and the removal phase (t(158) = 1.28, p = .20, d = 0.14, 95% CI [−.07, .39]) (see ). The tests of critical transitions between phases found that players initiated movement significantly later (t(16) = −2.35, p = .03, d = 0.80, 95% CI [.06, 1.52]) in the last trial of the baseline phase (trial 15) compared to the first weighted trial in the disturbance phase (trial 16). Furthermore, there was a significant relationship observed between delta change movement time and delta change player trial time between trial 15 and trial 16 (r = 0.65, p = .004). There were no significant differences in movement time (t(16) = 0.20, p = .20, d = 0.07 95% CI (−.60, .74)) and player trial time (t(16) = 1.661, p = .11, d = 0.56 95% CI (−.13, 1.26)) between the last weighted trial (trial 25) and the first trial where the weight was withdrawn (trial 26), neither was there a relationship observed between delta change movement time and delta change player trial time (r = 0.10, p = .15).
Figure 5. Movement time across the three testing phases.
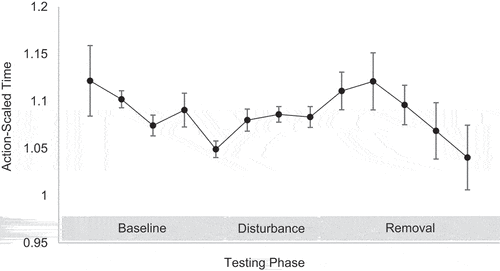
Action-Scaled Time: The baseline linear regression showed that trial number significantly predicted action-scaled time explaining 27% of the variance (F (1,14) = 6.37, p = .02, R2 = 0.27). Specifically, players were closer to their action boundary at the end of baseline compared to the start of baseline (). The disturbance and the removal linear regression both found that trial number did not significantly predict action-scaled time (F (1,9) = 0.56, p = .46, R2 = 0.06 & F (1,14) = 1.16, p = .30, R2 = 0.08, respectively). However, the baseline/disturbance quadratic regression showed that trial number significantly predicted action-scaled time explaining 23% of the variance (F (2,22) = 3.29, p = .05, R2 = 0.23). The same analysis for disturbance/removal was not significant (F (2,22) = 1.53, p = .23, R2 = 0.12). T-tests revealed that there were no significant difference in action-scaled time between the last five baseline trials (M = 1.08 ± .01) and the last five removal trials (M = 1.06 ± .02; t(81) = 0.91, p = .46, d = 0.08 95% CI (−.16, .44)), which suggests that players action-scaled time returned to those produced pre-disturbance.
Figure 6. Action-scaled time across the three testing phases. Action-scaled time = player trial time + movement time/tester trial time.
3.2. Gaze behaviour
Between Testing Phases: The number of fixations was significantly different between the three testing phases (i.e., baseline, disturbance and removal) (X2 (2) = 11.83, p = .03). Post-hoc analysis revealed that players made significantly less fixations in removal compared to baseline (z = 0.52, p = .01, r = 0.27, 95% CI [.19, .55]). Furthermore, there were no significant differences between baseline and disturbance (z = 0.19, p = .27, r = 0.12, 95% CI [.03, .32]) and disturbance and removal (z = 0.33, p = .06, r = 0.20, 95% CI [.10, .44]), respectively. There were no statistically significant observations in the remaining gaze variables (see ). Furthermore, there were no significant difference (p = .54) in final fixation location between baseline (50% legs; 17% COD-A; 33% other), disturbance (53% legs; 15% COD-A; 32% other), and removal (44% legs; 19% COD-A; 37% other).
Table 1. Between testing phases gaze behaviour with Friedman test statistics.
Successful Versus Unsuccessful Trials: Despite small effects being observed, there were no significant differences in the duration of fixations between successful and unsuccessful trials (z = −1.52, p = .12, r = 0.28, 95% CI [−.01, .58]). Likewise, albeit non-significant, the players tended to fixate longer in the final fixation during successful trials compared to unsuccessful trials (z = −1.45, p = .14, r = 0.25, 95% CI [−.04, .54]). Furthermore, there were no significant differences between successful and unsuccessful trials for any other gaze behaviour measure (see ). Furthermore, there was no significant difference (p = .96) in final fixation location between successful (57% legs; 10% COD-A; 33% other) and unsuccessful trials (55% legs; 12% COD-A; 33% other).
Table 2. Wilcoxon-Signed Rank tests of gaze behaviour variables. Successful compared to unsuccessful trials.
Changes in Gaze Behaviour Across Testing Phases: The baseline linear regression showed that trial number did not significantly predict number of fixations (F (1,14) = 1.25, p = .28, R2 = 0.08), duration of fixations (F (1,14) = 0.05, p = .81, R2 = 0.01), scan ratio (F (1,14) = 0.21, p = .64, R2 = 0.02) or duration of final fixation (F (1,14) = 0.75, p = .40, R2 = 0.06). Furthermore, in the baseline phase, there were no significant changes in final fixation location as a function of trial number: legs (X2 (1,14) = 7.83, p = .09, R2 = 0.10), COD-A (X2 (1,14) = 4.17, p = .12, R2 = 0.08) and other (X2 (1,14) = 1.01, p = .79, R2 = 0.01). The baseline/disturbance quadratic regression analysis showed that trial number predicted number of fixations and accounted for 35% of the variance (see ). The disturbance/removal regression did not significantly predict number of fixations per trial despite accounting for 19% of the variance (see ). However, the linear regression of all available gaze data trials showed that trial number significantly predicted number of fixations and accounted for 49% of the variance (F (1, 39) = 37.331, p = .001, R2 = .49). That is, players utilized fewer fixations as trial number increased. Analysis of the three other gaze behaviour variables showed that trial number did not significantly explain a significant portion of the observed variance (see ).
Table 3. Quadratic regression analysis of gaze behaviour between two testing transitions.
4. Discussion
The current study implemented the framework of Brand and de Oliveira (Citation2017) to examine the processes of perceptual-motor calibration/recalibration of defensive football players in a 1-vs-1 anticipation test. Ankle weights were used to reduce the 5-m acceleration capabilities of football players, to examine how the players adapted to the disturbance in terms of when movement was initiated and how gaze behaviour changed. It was hypothesized that a reduction in 5-m acceleration capabilities would disturb the perceptual-motor system. As a result, if players recalibrated, this would be reflected in consistent earlier movement times in the 1-vs-1 anticipation test and if players adapted the information that they exploited, this would be exemplified by changes in gaze behaviours. Furthermore, it was expected that a recalibration process would be required for players to rescale the link between perception-action when the ankle weights were applied and withdrawn (Brand & de Oliveira, Citation2017; Scott & Gray, Citation2010).
Firstly, the findings of the current study highlight the importance of providing players the opportunity to “scale” action to perceptual information (Withagen & Michaels, Citation2004). Players new to the 1-vs-1 anticipation test became better calibrated to the test with exploration (i.e., trials), with trial number explaining a significant proportion of the variance in the measure of action-scaled time prior to the disturbance. As designed, the implementation of ankle weights resulted in a perceptual-motor disturbance (Nakamoto et al., Citation2012; Scott & Gray, Citation2010), as participants were significantly slower in the 1-vs-1 anticipation test whilst wearing the ankle weights. Moreover, to contend with the perceptual-motor disturbance, players initiated movement significantly earlier in the 1-vs-1 anticipation test. More specifically, at the critical transition from unweighted (final baseline trial) to weighted (first disturbance trial) there was a significant relationship between the change in player trial time (i.e., action capabilities) and the change in when movement was initiated. Specifically, greater changes in players movement times were associated with relatively earlier initiation of the players’ response to their opponent (i.e., the tester). The disturbing effect of the ankle weights on the perceptual-motor system was further highlighted by the quadratic regression analysis that showed that the transition between baseline and disturbance phase explained a significant portion of the variance in action-scaled time. Specifically, players acted closer to the limits of their action capabilities immediately prior to the implementation of the ankle weights and action-scaled time was negatively impacted by the ankle weights. Moreover, players appeared unable to contend with the disturbance within the 10 trials of the disturbance phase. The initial withdrawal of ankle weights at the removal phase seemed to act as a further disturbance; however, within 11–15 trials, players were once again operating closer to the limits of their action capabilities. This finding therefore appears to differ from the results of Scott and Gray (Citation2010) who found that, following a change from heavier to lighter and lighter to heavier baseball bats, batters were able to recalibrate within 5–10 trials after the bat change.
This discrepancy in findings between the current study and the work of Scott and Gray (Citation2010) supports the view of Brand and de Oliveira (Citation2017) that perceptual-motor recalibration is likely task specific. There is some evidence from daily perception-action tasks, such as walking and stepping over objects (van Hedel & Dietz, Citation2004) that larger disturbances result in longer recalibration periods (van Andel et al., Citation2017). In other words, when a disturbance causes a greater error, the recalibration period is longer. Therefore, the discrepancy in the time taken to recalibrate between the 1-vs-1 anticipation test and simulated baseball pitches (Scott & Gray, Citation2010) may be explained by the magnitude of the disturbance placed on the perceptual-motor system in the 1-vs-1 anticipation test. Returning to the results of the present study specifically, the results suggest that each trial gave participants the opportunity to move closer to their maximal action capabilities.
Action-scaled time (AST) is a relational measure of both a player and opponent’s action capabilities, which in the context of the 1-vs-1 anticipation test is time taken to travel 5-m. Based on the assumption that athletes can be sensitive to their own action capabilities (van der Kamp et al., Citation2018), beating the tester to the endpoint in the 1-vs-1 anticipation test requires that the initiation of movement is scaled according to a player’s own action capabilities relative to the opponent. The results of the present study show that affordance-based control (Fajen, Citation2005) may offer a suitable approach for understanding how defensive players contend with situations that are characterized by spatiotemporal constraints (see Zheng et al., Citation2021). Accordingly, affordance-based control holds that in successful performance, players act in a way that action requirements do not exceed their maximum capabilities (Fajen et al., Citation2009). In the 1-vs-1 defensive anticipation task examined in the current study, evidence for affordance-based control only became apparent when a measure of the defender’s action capabilities and their initiation of movement was considered in conjunction with the attacker’s action capabilities. Therefore, future work in this area will need to consider a measure of both the defender’s and their direct opponent’s maximum action capabilities and an indication of the defender’s scaling of their action capabilities in comparison to that of their opponent.
The current study is the first examination of gaze behaviours when changes in the perceptual-motor system (i.e., application and withdrawal of ankle weights) disturb perception-action during a sport anticipation situation. Specifically, it was expected that when the ankle weights were applied, players would move earlier and, therefore, fixate on the earlier unfolding kinematic information of the tester (e.g., head, torso and hips), resulting in an earlier onset of final fixation. However, there were no changes in the gaze behaviour dependent variables used in this study as a result of ankle weights being applied and withdrawn. There were less fixations at the disturbance phase compared to the baseline phase and removal phase compared to disturbance phase, respectively. That is, players utilized fewer fixations as trial number increased and not as a result of the application and withdrawal of the ankle weights. As expected, players converged towards the later kinematic locations provided by the legs compared to the head, torso, hips and change of direction point combined, but there was no difference in final fixation location between successful and unsuccessful trials. That is, players were just as likely to fixate on the legs when they were unsuccessful (action-scaled time = ≥1.00), as when they were successful (action-scaled time = ˂ 1.00) at beating the attacker to the 5-m endpoint. In the context of the 1-vs-1 anticipation test, the legs may be viewed as “honest” kinematic information (Brault et al., Citation2012), so performance differences in the 1-vs-1 anticipation test seem not to be explained by the locations of exploited kinematic information. However, when comparisons were made globally (i.e., across testing phases) between successful and unsuccessful trials, players tended to fixate longer in successful trials (small effect) and the duration of final fixation tended to be longer in successful trials compared to unsuccessful trials (small effect). Therefore, despite there being no difference in final fixation location between successful and unsuccessful trials, it is likely that a longer, earlier final fixation of the legs may have had positive implications on 1-vs-1 anticipation test performance. Taken together, these gaze behaviour findings show in the context of a 1-vs-1 anticipation task examined in the current study, adaptation of affordance-based control appeared to be relative to movement time as opposed to changes in the gaze behaviours measured.
Alternatively, the failure of the disturbance to have a noticeable effect on visual search behaviour may be a result of the relatively low complexity of the 1-vs-1 anticipation test. The test required players to anticipate the timing of an attacker’s 90° direction change and the attacker was requested to change direction by planting his left foot to change direction right or by planting his right foot to change direction left. It is plausible that the kinematic information that unfolded in the attacker’s actions prior to this point specified their directional change, and participants may have attuned to this information, irrespective of the changes to their own action capabilities. Increasing the complexity of the task, for example, the inclusion of a ball with alternative passing options or trials containing purposefully deceptive actions may have resulted in greater changes in perceptual attunement when the perceptual-motor system had been disturbed, which may have manifested in changes in gaze behaviour. In addition, conventional gaze behaviour measures were used in the current study that have commonly been adopted in anticipation studies. It is likely that these variables may not adequately capture the changes in gaze patterns that reflect calibration. Therefore, future work would benefit from the introduction of new gaze measures, such as entropy to reflect exploration (Hacques et al., Citation2022).
In the current study, ankle weights were imposed on players to compromise their capability to accelerate. Players' immediate adaptation to the disturbance was to move earlier in response to their opponent’s change in direction. This supports the assumption that athletes are sensitive to their own action capabilities (see van der Kamp et al., Citation2018). From an applied point of view, practitioners should therefore consider the impact of reduced action capabilities (e.g., fatigue) not only on physical performance outcomes but also on the anticipatory (perceptual-motor) skill of players. Time is needed to rescale the link between perception and action, and therefore, players should be given sufficient opportunity to train under reduced action capabilities (Alder et al., Citation2019). This would provide players with the opportunity to re-/calibrate and learn to better contend with disturbances of the perceptual-motor system.
Fatigue can be considered as a fast-changing constraint because it can change over the course of a game (Balagué et al., Citation2019). Other important constraints change more slowly. For example, action capabilities will change as a result of natural occurring geometric factors, such as the rapid increase in height, weight, and limb length during the adolescent growth spurt. Future research is needed to examine the processes and mechanisms of calibration associated with changes to constraints, which may operate over different timescales. From an applied perspective, this is important work because perceptual-motor performance discriminates between players who participate in high-level football talent development programmes and those who do not (Deprez et al., Citation2015). There is evidence that the rapid gains in limb length during the period of time in which a child will experience their fastest growth in stature (Peak height velocity, Lloyd & Oliver, Citation2012) can lead to decrements in motor control performance (commonly referred to as “adolescent awkwardness”). The communication of improved understanding of the concepts of calibration/recalibration may encourage coaches to be more patient with talented individuals whose performance is below expectations.
The current study was the first to examine the processes of perceptual-motor calibration/recalibration of defensive players in a 1-vs-1 anticipation setting. However, it is crucial to acknowledge that previous research that provided the rationale for this work was arguably better able to isolate the concepts of calibration/recalibration (e.g., Scott & Gray, Citation2010). When adopting a representative design as was the case in the current study, there is a possibility that variables extraneous to the study of calibration/recalibration may have impacted upon athlete performance. For example, it is plausible that the kinematic information that unfolded in the tester’s actions specified their directional change, so players may have attuned to this information, irrespective of the changes to their own action capabilities (see also, Dicks et al., Citation2010b). However, it should be acknowledged that movement time and action-scaled timing error may be due to the pick-up of information, specifically, the player may have failed to pick-up the information that specified the direction change and the timing of direction change of the tester. Nevertheless, the 1-vs-1, attacker–defender interaction is a feature of invasion sport (e.g., Football, Rugby & Basketball) and if such settings are to be studied in order to enhance understanding of performance and injury prevention (Kadlec et al., Citation2023), it is important that researchers strive for research protocols that are reflective of the tasks that characterize sport (Dicks et al., Citation2009).
In conclusion, the manipulation of action capabilities was found to alter the movement response of players but not their gaze behaviour. Therefore, it appears that adaptation of affordance-based control (Fajen, Citation2005) in a 1-vs-1 anticipation task is likely relative to movement time (scaling) as opposed to changes in gaze behaviour (attunement). A reduction in 5-m acceleration capabilities disturbed the perceptual-motor system and players initiated movement significantly earlier in the 1-vs-1 anticipation test, which supports the notion that athletes are sensitive to their own action capabilities (see Dicks et al., Citation2010a; van der Kamp et al., Citation2018). Analysis of perceptual-motor calibration/recalibration via the measure action-scaled time revealed that players were best calibrated at the end of the baseline phase and the application of ankle weights negatively impacted action-scaled time. That is, players moved earlier but not early enough to contend with their reduced speed. Moreover, the players did not appear to have sufficient time in this study to recalibrate in the disturbance phase. However, following the withdrawal of the ankle weights, the players were able to recalibrate within 11–15 trials. Despite the results showing players not adapting gaze behaviour as a result of the disturbance being placed on the perceptual-motor system, task familiarization resulted in changes in gaze patterns (i.e., less fixations). The findings of the current study highlight the importance of providing players the opportunity to “scale” action to perceptual information and support the view that calibration/recalibration is an iterative process, whereby each time a perception-action coupling is used, it updates the informational link between perception and action. In sum, whether it is a negative (e.g., fatigue) or a positive (e.g., faster) change in action capabilities, sportspeople require practice time to “rescale” the link between the perceptual information of the performance environment and their action.
Acknowledgments
Mr Archie Wilson and Mr George Wilson for their assistance in the data collection and data analysis process. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Abernethy, B., Farrow, D., Gorman, A., & Mann, D. L. (2012). Anticipatory behaviour and expert performance. In N. Hodges, & A. M. Williams (Eds.), Skill acquisition in sport: Research, theory and practice (2nd ed., pp. 287–305). Routledge .
- Alder, D., Ford, P. R., Causer, J., & Williams, A. M. (2014). The coupling between gaze behavior and opponent kinematics during anticipation of badminton shots. Human Movement Science, 37, 167–179. https://doi.org/10.1016/j.humov.2014.07.002
- Alder, D., Ford, P. R., Causer, J., & Williams, A. M. (2016). The effects of high-and low-anxiety training on the anticipation judgments of elite performers. Journal of Sport & Exercise Psychology, 38(1), 93–104 https://doi.org/10.1123/jsep.2015-0145
- Araujo, D., Davids, K., & Hristovski, R. (2006). The ecological dynamics of decision making in sport. Psychology of Sport and Exercise, 7(6), 653–676. https://doi.org/10.1016/j.psychsport.2006.07.002
- Balagué, N., Pol, R., Torrents, C., Ric, A., & Hristovski, R. (2019). On the relatedness and nestedness of constraints. Sports Medicine-Open, 5(1), 1–10. https://doi.org/10.1186/s40798-019-0178-z
- Brand, M. T., & de Oliveira, R. F. (2017). Recalibration in functional perceptual-motor tasks: A systematic review. Human Movement Science, 56(Pt B), 54–70. https://doi.org/10.1016/j.humov.2017.10.020
- Brand, M. T., & de Oliveira, R. F. (2023). Perceptual-motor recalibration is intact in older adults. Human Movement Science, 87, 103047. https://doi.org/10.1016/j.humov.2022.103047
- Brault, S., Bideau, B., Kulpa, R., Craig, C. M., & Gribble, P. L. (2012). Detecting deception in movement: The case of the side-step in rugby. PLoS One, 7(6), e37494. https://doi.org/10.1371/journal.pone.0037494
- Croft, J. L., & Bertram, J. E. (2017). Affordance boundaries are defined by dynamic capabilities of parkour athletes in dropping from various heights. Frontiers in Psychology, 8, 1571. https://doi.org/10.3389/fpsyg.2017.01571
- de Oliveira, R. F., Oudejans, R. R., & Beek, P. J. (2009). Experts appear to use angle of elevation information in basketball shooting. Journal of Experimental Psychology: Human Perception and Performance, 35(3), 750. https://doi.org/10.1037/a0013709
- Deprez, D. N., Fransen, J., Lenoir, M., Philippaerts, R. M., & Vaeyens, R. (2015). A retrospective study on anthropometrical, physical fitness, and motor coordination characteristics that influence dropout, contract status, and first-team playing time in high-level soccer players aged eight to eighteen years. The Journal of Strength & Conditioning Research, 29(6), 1692–1704. https://doi.org/10.1519/JSC.0000000000000806
- Dicks, M., Button, C., & Davids, K. (2010b). Examination of gaze behaviors under in situ and video simulation task constraints reveals differences in information pickup for perception and action. Attention, Perception and Psychophysics, 72(3), 706–720. https://doi.org/10.3758/APP.72.3.706
- Dicks, M., Davids, K., & Button, C. (2009). Representative task design for the study of perception and action in sport. International Journal of Sport Psychology. https://psycnet.apa.org/record/2010-05712-005
- Dicks, M., Davids, K., & Button, C. (2010a). Individual differences in the visual control of intercepting a penalty kick in association football. Human Movement Science, 29(3), 401–411. https://doi.org/10.1016/j.humov.2010.02.008
- Fajen, B. R. (2005). Perceiving possibilities for action: On the necessity of calibration and perceptual learning for the visual guidance of action. Perception, 34(6), 717–740. https://doi.org/10.1068/p5405
- Fajen, B. R., Riley, M. A., & Turvey, M. T. (2009). Information, affordances, and the control of action in sport. International Journal of Sport Psychology, 40(1), 79–107.
- Faul, F., Erdfelder, E., Lang, A. G., & Buchner, A. (2007). G*power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–191. https://doi.org/10.3758/BF03193146
- Gabbett, T. J., Kelly, J. N., & Sheppard, J. M. (2008). Speed, change of direction speed, and reactive agility of rugby league players. Journal of Strength and Conditioning Research, 22(1), 174–181. https://doi.org/10.1519/JSC.0b013e31815ef700
- Gibson, J. J. (1979). The ecological approach to visual perception. Houghton Mifflin.
- Hacques, G., Dicks, M., Komar, J., & Seifert, L. (2022). Visual control during climbing: Variability in practice fosters a proactive gaze pattern. PLoS One, 17(6), e0269794. https://doi.org/10.1371/journal.pone.0269794
- Holmqvist, K., Nyström, M., Andersson, R., Dewhurst, R., Jarodzka, H., & Van de Weijer, J. (2011). Eye tracking: A comprehensive guide to methods and measures. OUP Oxford.
- Hristovski, R., Davids, K., Araujo, D., & Button, C. (2006). How boxers decide to punch a target: Emergent behaviour in nonlinear dynamical movement systems. Journal of Sports Science and Medicine, 5(CSSI), 60–73. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3863932/
- Kadlec, D., Miller-Dicks, M., & Nimphius, S. (2023). Training for “worst-case” scenarios in sidestepping: Unifying Strength and Conditioning and perception–action approaches. Sports Medicine-Open, 9(1), 22. https://doi.org/10.1186/s40798-023-00566-8
- Lloyd, R. S., & Oliver, J. L. (2012). The youth physical development model: A new approach to long-term athletic development. Strength & Conditioning Journal, 34(3), 61–72. https://doi.org/10.1519/SSC.0b013e31825760ea
- Mecheri, S., Laffaye, G., Triolet, C., Leroy, D., Dicks, M., Choukou, M. A., & Benguigui, N. (2019). Relationship between split-step timing and leg stiffness in world-class tennis players when returning fast serves. Journal of Sports Sciences, 37(17), 1962–1971. https://doi.org/10.1080/02640414.2019.1609392
- Nakamoto, H., Ishii, Y., Ikudome, S., & Ohta, Y. (2012). Kinesthetic aftereffects induced by a weighted tool on movement correction in baseball batting. Human Movement Science, 31(6), 1529–1540. https://doi.org/10.1016/j.humov.2012.04.005
- Navia, J. A., Dicks, M., van der Kamp, J., & Ruiz, L. M. (2017). Gaze control during interceptive actions with different spatiotemporal demands. Journal of Experimental Psychology: Human Perception and Performance, 43(4), 783. https://doi.org/10.1037/xhp0000347
- Passos, P., Araujo, D., Davids, K., Gouveia, L., Milho, J., & Serpa, S. (2008). Information-governing dynamics of attacker-defender interactions in youth rugby union. Journal of Sports Sciences, 26(13), 1421–1429. https://doi.org/10.1080/02640410802208986
- Ramenzoni, V. C., Riley, M. A., Shockley, K., & Davis, T. (2008). Short article: Carrying the height of the world on your ankles: Encumbering observers reduces estimates of how high an actor can jump. Quarterly Journal of Experimental Psychology, 61(10), 1487–1495. https://doi.org/10.1080/17470210802100073
- Ramsey, H., Button, C., Davids, K., Hacques, G., Seifert, L., & Dicks, M. (2020). Considerations for the study of individual differences in gaze control during expert visual anticipation: An exploratory study. Movement & Sport Sciences - Science & Motricité, 4(110), 39–47. https://doi.org/10.1051/sm/2020006
- Rietveld, E., & Kiverstein, J. (2014). A rich landscape of affordances. Ecological Psychology, 26(4), 325–352. https://doi.org/10.1080/10407413.2014.958035
- Roca, A., Ford, P. R., McRobert, A. P., & Williams, A. M. (2013). Perceptual-cognitive skills and their interaction as a function of task constraints in soccer. Journal of Sport & Exercise Psychology, 35(2), 144–155. https://doi.org/10.1123/jsep.35.2.144
- Scanlan, A., Humphries, B., Tucker, P. S., & Dalbo, V. (2014). The influence of physical and cognitive factors on reactive agility performance in men basketball players. Journal of Sports Sciences, 32(4), 367–374. https://doi.org/10.1080/02640414.2013.825730
- Scott, S., & Gray, R. (2010). Switching tools: Perceptual-motor recalibration to weight changes. Experimental Brain Research, 201(2), 177–189. https://doi.org/10.1007/s00221-009-2022-z
- Seifert, L., Dicks, M., Wittmann, F., & Wolf, P. (2021). The perception of nested affordances: An examination of expert climbers. Psychology of Sport and Exercise, 52, 101843. https://doi.org/10.1016/j.psychsport.2020.101843
- van Andel, S., Cole, M. H., & Pepping, G. J. (2017). A systematic review on perceptual-motor calibration to changes in action capabilities. Human Movement Science, 51, 59–71. https://doi.org/10.1016/j.humov.2016.11.004
- van der Kamp, J., Dicks, M., Navia, J. A., & Noël, B. (2018). Goalkeeping in the soccer penalty kick. German Journal of Exercise and Sport Research, 48(2), 169–175. https://doi.org/10.1007/s12662-018-0506-3
- van der Kamp, J., Rivas, F., Van Doorn, H., & Savelsbergh, G. (2008). Ventral and dorsal system contributions to visual anticipation in fast ball sports. International Journal of Sport Psychology, 39(2), 100–130.
- van Hedel, H. J., & Dietz, V. (2004). Obstacle avoidance during human walking: Effects of biomechanical constraints on performance. Archives of Physical Medicine and Rehabilitation, 85(6), 972–979. https://doi.org/10.1016/j.apmr.2003.07.006
- Williams, A. M., & Davids, K. (1998). Visual search strategy, selective attention, and expertise in soccer. Research Quarterly for Exercise and Sport, 69(2), 111–128. https://doi.org/10.1080/02701367.1998.10607677
- Wilson, R., Alder, D., Dicks, M., & Poolton, J. (2021). Examining the effect of reduced action capabilities on defensive anticipation in a 1-vs-1 task. Science and Medicine in Football, 5(3), 234–241. https://doi.org/10.1080/24733938.2020.1865561
- Wilson, R., Dicks, M., Milligan, G., Poolton, J., Alder, D., Dicks, M., O’Halloran, J., & Navarro, M. (2019). An examination of action capabilities and movement time during a soccer anticipation task. Movement & Sport Sciences - Science & Motricité, 102(102), 61–70. https://doi.org/10.1051/sm/2019001
- Withagen, R., & Michaels, C. F. (2004). Transfer of calibration in length perception by dynamic touch. Perception and Psychophysics, 66(8), 1282–1292. https://doi.org/10.3758/bf03194998
- Zheng, R., de Reus, C., & van der Kamp, J. (2021). Goalkeeping in the soccer penalty kick: The dive is coordinated to the kicker’s non-kicking leg placement, irrespective of time constraints. Human Movement Science, 76, 102763. https://doi.org/10.1016/j.humov.2021.102763
- Zheng, R., van der Kamp, J., Song, X., & Savelsbergh, G. (2022). Affordance-based control in deceptive and non-deceptive penalties in soccer goalkeeping: Gender matters! Psychology of Sport and Exercise, 58, 102096. https://doi.org/10.1016/j.psychsport.2021.102096