
ABSTRACT
This study aimed to investigate inter- and intra-athlete technique variability in pre-elite and elite Australian fast bowlers delivering new ball conventional swing bowling. Ball grip angle and pelvis, torso, shoulder, elbow, wrist, upper arm, forearm, and hand kinematics were investigated at the point of ball release for inswing and outswing deliveries. Descriptive evaluations of group and individual data and k-means cluster analyses were used to assess inter- and intra-bowler technique variability. Inter-athlete technique and ball grip variability were identified, demonstrating that skilled bowlers use individualised strategies to generate swing. Functional movement variability was demonstrated by intra-athlete variability in successful swing bowling trials. Bowlers demonstrated stable technique parameters in large proximal body segments of the pelvis and torso, providing a level of repeatability to their bowling action. Greater variation was observed in bowling arm kinematics, allowing athletes to manipulate the finger and ball position to achieve the desired seam orientation at the point of ball release. This study demonstrates that skilled bowlers use individualised techniques and grips to generate swing and employ technique variations in successive deliveries. Coaches should employ individualised training strategies and use constraints-led approaches in training environments to encourage bowlers to seek adaptive movement solutions to generate swing.
Introduction
In cricket, bowlers are categorised as either spin or fast bowlers based on the spin and speed they impart on the ball. Spin bowlers are sub-classified as either finger spin or wrist spin bowlers based on the direction of spin they impart on the ball, and often use slow release speeds (Beach et al., Citation2016). Fast bowlers aim to release the ball at higher speeds and are sub-classified as either medium (release speed of 27–36 m/s) or fast paced (release speed greater than 36 m/s) (Abernethy, Citation1981). Fast bowlers project the ball towards batters at high speeds to limit their reaction time to restrict run scoring and claim wickets. A tactic they can employ to promote this is swing bowling, particularly when the ball is new. Swing bowling is the horizontal deviation of the ball as it travels towards the batter. To create swing, bowlers use techniques that impact the speed, seam orientation and spin imparted on the ball (Lindsay & Spratford, Citation2020; Lindsay et al., Citation2022; Mehta, Citation2005). Bowlers can create outswing, where the ball deviates away from the batter’s body, and inswing, where the ball swings towards the batter’s body. Swing bowling has reduced the performance of batters in controlled testing environments (Sarpeshkar et al., Citation2017) and reduced scoring and increased dismissal rates in matches (Mehta et al., Citation2022). If bowlers can create swing, it is a tactic that can create a competitive advantage.
Recent research has begun to understand the tactics and techniques highly skilled bowlers and coaches employ for swing bowling (Lindsay et al., Citation2024). Through interviews with elite Australian bowlers and coaches, researchers revealed that bowlers aim to release the ball with an upright and angled seam to generate swing. However, when participants described their strategies to achieve this, there were differences in the arm, hand and finger positions between bowlers (inter-individual variability). Additionally, some bowlers reported using technique variations (intra-individual variability) to create the same and opposite direction of swing which may also highlight their intention to create uncertainty for the batter. This study identified potential inter- and intra-athlete technique variability, however, the authors suggested that biomechanical investigations were required to link participant perceptions with their actions.
Investigations of movement variability in fast bowling are limited. Salter et al., (Citation2007) investigated ball release height, centre of mass velocity, front knee angle, hip-shoulder alignment angle, stride length, arm angular velocity and ball release speed in a group of fast bowlers and a single fast bowler. When comparing the standard deviations for between-bowlers and within-bowler conditions, greater variability was reported in group results. Perrett et al., (Citation2020) examined spinal lateral flexion and axial rotation velocity, and reported inter-athlete movement variability between front foot contact and ball release, along with lower levels of intra-athlete variability. Based on these studies (Perrett et al., Citation2020; Salter et al., Citation2007), low intra-athlete variation and greater inter-athlete variation demonstrate individualised techniques.
Dynamical systems theory explains that individual movement patterns are established by the process of self-organisation (Kelso, Citation1995). The complex nature of fast bowling means that many different degrees of freedom can be used to deliver the ball. Inter-athlete movement variability is linked to intrinsic dynamics and affected by factors such as anthropometry, joint mobility, muscular strength and previous experiences, leading to the development of individualised coordination patterns (Glazier & Mehdizadeh, Citation2019; Salter et al., Citation2007). Athletes also adapt their movements to the constraints of dynamic performance environments (Andrews et al., Citation2024; Crowther et al., Citation2023; Phillips et al., Citation2014; Seifert et al., Citation2013). Ecological dynamics considers the interaction between individual, task, and environment constraints in sporting tasks (Crowther et al., Citation2023; Newell & Corcos, Citation1993). The complexity involved with repeatedly finding a solution to satisfy the demands of a sporting task where the constraints change from one trial to the next leads to functional movement variability, demonstrating the motor system’s flexibility and adaptability (Crowther et al., Citation2023). Athletes continuously adjust their technique to achieve consistent performance outcomes (Robins et al., Citation2006). When considering inter-athlete technique variations and functional movement variability (intra-athlete variability), no one solution exists to complete the task of bowling. The infinite number of possible movement solutions has potential to cause confusion among athletes and coaches about how to best encourage swing bowling for any one individual.
Athletes use technique parameters with different levels of variation to consistently produce desired performance outcomes. Invariant movement components, with relatively low variability, increase the consistency of repeated movements. These invariant characteristics are often captured in traditional coaching manuals (Woolmer et al., Citation2008), however, they do not capture the subtle adjustments being made as the individual interacts with the task, environment or individual constraints. Tennis researchers identified that athletes have low variance in trunk kinematics during serves, suggesting this provides stability to “posture” during the movement (Whiteside et al., Citation2015). Conversely, greater variation was observed in arm kinematics as athletes manoeuvred the racquet to strike the ball in varying locations due to inconsistencies in the ball toss. These findings are similar to the perceptions of elite bowlers and coaches, provided in published research, who discussed the techniques used by fast bowlers (Lindsay et al., Citation2024; Phillips et al., Citation2014). It was suggested that elite fast bowlers use invariant technique aspects in the larger proximal body segments (such as the torso) and variable technique aspects in the smaller distal body segments (such as the bowling arm). Variant components provide athletes with opportunities to adjust their movement and adapt to constraints during a performance, whether that be in competition or not. Fast bowlers may use a combination of invariant and variant technique parameters to consistently generate swing.
This study aimed to investigate inter- and intra-athlete technique variability of pelvis and upper-body kinematics in fast bowlers delivering swing with new cricket balls. We hypothesised that 1) athletes would use individualised techniques, 2) participants would demonstrate intra-athlete variability with invariant and variant technique components, and 3) relatively less variance would be observed in the pelvis and torso due to their large size and proximal location, with greater variation found in the bowling arm. Investigating inter- and intra-athlete technique variations will provide insights into individualised bowling actions and direct coaches to stable and variant technique aspects that allow athletes to consistently generate swing.
Methods
Participants
Twelve Australian male elite (international u19, n = 4 and senior state, n = 6) and pre-elite (state u19, n = 2) medium and fast bowlers (age 19.9 ± 2.1 years, mass 85.1 ± 9.7 kg, height 192.5 ± 4.9 cm, maximum release speed 34.5 ± 1.3 m/s) participated in this study. Convenience sampling was used to recruit participants who were attending a post-season training camp at the time of testing. Pre-elite and elite fast bowlers were chosen to participate as the research team believed they would have the skillset required to consistently generate swing. For the purpose of this study, medium and fast pace participants are collectively termed “fast bowlers” (Abernethy, Citation1981). Individual participants are referred to using alphanumeric combinations. For example, participant one and participant twelve are referred to as P1 and P12, respectively. All participants provided informed consent. The University of Canberra Human Research Ethics Committee approved this study (approval #4645).
Data collection
Data collection was conducted at an indoor training facility with artificial wickets and space for full-length run-ups. Retro-reflective markers were affixed to athletes in accordance with the University of Western Australia (UWA) full-body marker set (Besier et al., Citation2003; Campbell, Alderson, et al., Citation2009; Campbell, Lloyd, et al., Citation2009; Chin et al., Citation2010; Lloyd et al., Citation2000; Wells et al., Citation2018). Three (diameter = 13 mm, thickness = 0.1 mm, mass = 0.02 grams) retro-reflective tape patches (3M Scotchlite High Gain Reflective Sheeting 7610) were affixed to new four-piece red Kookaburra Turf cricket balls (mass 0.156 kg) in locations used in previous research (Sakurai et al., Citation2013; Spratford et al., Citation2018; Whiteside et al., Citation2013). This tape was chosen to minimise the aerodynamic influence on the ball during flight. After marker placement, participants completed static and dynamic trials to determine joint centres and functional axes of rotations. They then completed a self-selected warm-up and were familiarised with testing procedures.
Each participant was right-handed and bowled three overs (18 deliveries/trials) consisting of nine outswing and nine inswing deliveries as though bowling to a right-handed batter. The delivery order was block randomised and the swing direction was counterbalanced to ensure there was no order effect. Participants either delivered six outswing, six inswing, three outswing and three inswing deliveries or the same delivery structure with the opposing swing direction. This allowed participants to make technical adjustments based on the outcome of previous trials and replicated a match situation (Connor et al., Citation2020; Lindsay & Spratford, Citation2020).
Each participant used one new cricket ball. Participants gripped the ball so the side with two markers was facing the intended direction of swing to minimise aerodynamic interference. Before each delivery, the ball grip angle was measured using a protractor. This measured the angle created by the primary seam relative to the index and middle fingers. Zero degrees represents a seam parallel to the fingers and positive and negative angles represent a seam angled towards the right and left, respectively, in relation to the bowler’s direction of travel. Participants were instructed to replicate match conditions as closely as possible by bowling over the wicket as though bowling to a right-handed batter, generating swing in the desired direction and pitching the ball on a good-to-full length approximately three-to-seven metres from the batter’s stumps (Justham et al., Citation2010; Lindsay et al., Citation2024) at match intensity. After each delivery, the ball was assessed for scuffs or damaged markers. If required, the ball was polished, or markers were replaced to maintain a smooth surface.
Body and ball marker trajectories were captured by a 40-camera (MX T40) Vicon motion capture system (Oxford Metrics Ltd., Oxford, UK) sampling at 250 Hz. The system captured the pre-delivery stride, delivery stride and the entirety of the ball flight when pitched at least two metres from the stumps at the batter’s end. The global origin was positioned at the intersection of the bowling and return creases on the left-hand side as viewed from the bowler’s end of the wicket. The x-axis pointed down the wicket, the y-axis was orthogonal to the wicket and the z-axis upwards. A video camera (Vicon Bonita) capturing 125 frames per second positioned behind the stumps at the bowler’s end was used to identify wide deliveries for exclusion from the dataset.
Data processing
Three-dimensional body and ball marker trajectories were reconstructed and labelled using Vicon Nexus software v2.12 (Oxford Metrics Ltd., Oxford, UK). Body marker trajectories were filtered using a fourth-order zero-lag Butterworth filter with a cut-off frequency of 14 Hz, following a residual analysis and visual inspection of the data (Winter, Citation2009). Data were modelled using the valid and reliable UWA upper and lower-body models (Besier et al., Citation2003; Campbell, Alderson, et al., Citation2009; Campbell, Lloyd, et al., Citation2009; Chin et al., Citation2010; Lloyd et al., Citation2000; Wells et al., Citation2018) to calculate kinematics. Shoulder joint centres were calculated with a regression equation using the location of markers and participant height and mass (Campbell, Alderson, et al., Citation2009; Campbell, Lloyd, et al., Citation2009). Elbow joint centres were defined as the mid-point point between medial and lateral humeral epicondyle markers (Chin et al., Citation2010; Wells et al., Citation2018). Wrist joint centres were defined as the mid-point between medial and lateral styloid process markers (Chin et al., Citation2010; Lloyd et al., Citation2000). Hip joint centres were calculated using optimal common shape technique (OCST) and symmetrical centre of rotation estimation (SCoRE) Vicon Nexus processes during a hip circumduction task (Ehrig et al., Citation2006). Knee joint centres were calculated using OSCT and symmetrical axis of rotation estimation (SARA) Vicon Nexus processes during a squatting task (Ehrig et al., Citation2007). Ankle joint centres were defined as the mid-point between medial and lateral malleolus markers. Kinematics were normalised to 100% (101 points) of the delivery stride between back foot contact and ball release using a custom cubic spline Python script. Back foot contact was defined as the frame when the foot marker trajectories were observed to change due to foot contact with the ground (Felton et al., Citation2019; Worthington et al., Citation2013). Ball release was defined as the frame when the distance between a hand and ball marker increased by greater than 20 mm relative to the previous frame (Felton et al., Citation2019; Lindsay & Spratford, Citation2020; Worthington et al., Citation2013).
Pelvis, torso and bowling arm kinematics were investigated. Global z orientation measured segment rotation in the global z-axis. Zero degrees was orientated down the wicket towards the batter’s end, increasing positively in a clockwise direction and negatively in an anti-clockwise direction as viewed superiorly. Global y orientation measured segment lateral rotation in the global y-axis. Zero degrees was orientated perpendicular to the ground, increasing positively in a clockwise direction and negatively in an anti-clockwise direction, as viewed from the bowler’s end of the wicket. Joint angles are reported using the International Society of Biomechanics recommendations (Wu et al., Citation2002, Citation2005). Positive angles represent thorax flexion, right-side thorax lateral flexion, shoulder adduction, elbow flexion, elbow pronation, wrist ulnar deviation and wrist flexion. Negative angles represent thorax extension, left-side thorax lateral flexion, shoulder abduction, elbow extension, elbow supination, wrist radial deviation, and wrist extension. Pelvis, trunk, and thorax kinematics provided insight into body position and potential invariant technique components (Lindsay et al., Citation2024; Phillips et al., Citation2014; Whiteside et al., Citation2015). Bowling arm and hand kinematics were included as they can manipulate the ball orientation at release (Lindsay et al., Citation2022, Citation2024). All body kinematics are reported at ball release. Delivery swing was calculated using methods outlined in previous research (Lindsay & Spratford, Citation2020). Inswing deliveries swung to the right and outswing deliveries swung to the left relative to the original direction of travel as viewed from the bowler’s end of the wicket, with respect to a right-handed batter.
To investigate functional movement variability, only successful trials were analysed. Bowlers must have achieved the desired outcome (pitched in the designated area, correct swing direction, and travelled between the wide lines for a right-handed batter) and created at least 0.2° of swing. When pitched on a good length, approximately six metres from the batter’s stumps (Justham et al., Citation2010), this amount of swing equates to a lateral deviation of approximately three centimetres to the left or right of the initial trajectory when the ball bounces (Lindsay & Spratford, Citation2020). Researchers have reported reduced ball speed and deviation from the intended target line for bat-and-ball impacts greater than three centimetres from the midline of cricket bats (Bower, Citation2012; Peploe et al., Citation2018). Therefore, in match situations, this amount of swing could provide a competitive advantage to bowlers by reducing scoring rates and increasing the opportunities to claim wickets. Across both swing directions, unsuccessful trials were excluded for pitching outside the designated area (n = 20), swinging in the wrong direction (n = 31), swinging less than 0.2° (n = 55) and being a wide delivery (n = 20). In total, 50 trials across 11 participants were included for outswing and 40 trials across eight participants were included for inswing.
Data analysis
The following process outlines the exploratory data analysis used to understand inter- and intra-athlete variability for inswing and outswing deliveries. This series of steps were completed independently for each swing direction. All analyses were conducted in RStudio v2022.07.2 (RStudio Team, Boston, MA, USA). Descriptive statistics were calculated for each variable on a group (mean, standard deviation, and minimum and maximum values) and individual (minimum and maximum values due to varying amounts of successful trials) level. This allowed descriptive evaluations within variable scales and variance structures before further exploration.
The following steps were undertaken relative to the implementation of a clustering model involving body kinematic variables. Variables were normalised relative to a standard normal distribution (μ = 1, σ = 0) before entry into two separate multivariate k-means clustering models to account for differences in within-variable variances and unit scaling. Each k-means model was created for inswing and outswing trials. The gap statistic determined the number of clusters to be retained within an iterative clustering algorithm (Tibshirani et al., Citation2001). The gap statistic was run using the R package Cluster v2.1.2, with the maximum number of clusters pre-set to the number of participants included in each swing direction. The number of clusters (k) was then selected, such that Gap(k) ≥ Gap(k +1) – sk +1, which conceptually identified the point at which the rate of increase of the gap statistic decreased. K-means clustering was conducted using the R package Cluster v2.1.2 (Hartigan & Wong, Citation1979). The number of clusters (k) was then selected according to the gap statistic and the outcomes of clustering were descriptively analysed by 1) the creation of participant x cluster matrices demonstrating counts of the number of trials in each cluster for each participant, and 2) visualising the results of cluster assignment at the participant and trial level descriptively using scatterplots. The variables included in the scatterplots were chosen to display a range of kinematics and to explore the hypothesis of stable and variable body kinematics.
Results
presents group means, standard deviations and ranges for outswing and inswing. Large variances were found in ball grip angle, global body segment orientations, wrist radial/ulnar deviation angular velocity, elbow measures and shoulder abduction. On average, participants generated 0.8° and 0.5° of swing for outswing and inswing deliveries, respectively. Individual participant ranges are presented in Appendix .
Table 1. Group means, standard deviations and ranges for ball grip angle, body kinematics at ball release and delivery swing angles.
The gap statistic determined the optimal number of clusters was ten and nine for outswing and inswing, respectively. shows the participant x cluster matrix for outswing deliveries. P1, P2, P5, P6, P7, P9, and P10 had most trials assigned to one cluster while P4, P8, P11 and P12 showed a greater spread across multiple clusters. P3 did not bowl any successful outswing deliveries.
Table 2. Matrix demonstrating the number of trials for each participant included in each cluster for outswing deliveries.
shows the participant x cluster matrix for inswing deliveries. P1, P4, P7, P8, and P10 had most trials assigned to one cluster while P3, P11 and P12 showed a greater spread across multiple clusters. P2, P5, P6 and P9 did not bowl any successful inswing deliveries.
Table 3. Matrix demonstrating the number of trials for each participant included in each cluster for inswing deliveries.
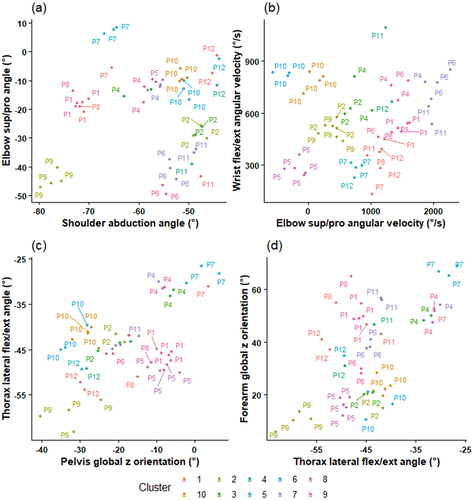
contains four scatterplots of variables included in the outswing cluster analysis. shows greater variance in elbow supination-pronation than shoulder abduction for most participants and clusters. demonstrates close groupings of most individual participants trials and clusters. shows low variance in pelvis orientation and lateral thorax flexion in most participants. demonstrates greater stability in thorax lateral flexion than forearm global z orientation in most participants and clusters.
Figure 1. Outswing scatterplots of kinematic variables and clusters. In the labelling on the scatterplots, “P” pertains to “participant” and the subsequent number is the ID of each participant.
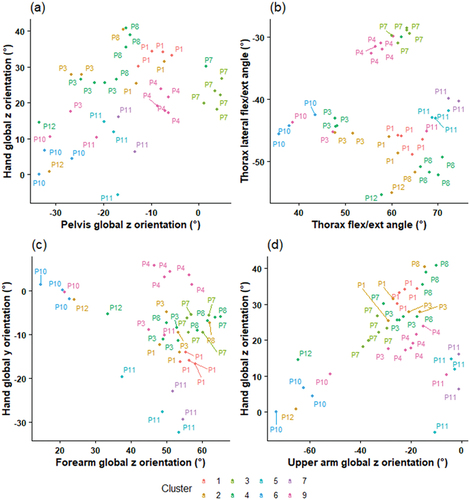
contains four scatterplots of variables included in the inswing cluster analysis. demonstrates greater variance in hand global z orientation than pelvis orientation. shows a close grouping of individual participant trials for thorax forward and lateral flexion, demonstrating low intra-athlete variation. demonstrates high variation of forearm and hand global orientations in most participants. shows variation in upper arm and hand global z orientations.
Figure 2. Inswing scatterplots of kinematic variables and clusters. In the labelling on the scatterplots, “P” pertains to “participant” and the subsequent number is the ID of each participant.
Discussion
Descriptive evaluations and k-means cluster analyses were used to investigate inter- and intra-athlete variability of conventional new ball swing bowling techniques in elite and pre-elite Australian fast bowlers. The hypotheses were confirmed with athletes demonstrating inter- and intra-individual variation, including invariant and variant parameters. Individualised coaching strategies should be employed, and introducing varying constraints in training environments can encourage athletes to explore movement patterns to generate swing.
Group variability and lower intra-athlete variation confirmed individualised techniques were employed. In the group results, elbow supination-pronation angle, hand global z orientation, thorax flexion-extension angle, and upper arm global z orientation displayed ranges greater than 30° for both swing directions (). Wrist ulnar-radial deviation angle had the lowest variation (16.9° for outswing, 12.2° for inswing). When comparing this variable in P1 and P2 outswing trials (), low intra-individual but large inter-individual variation was found (11.9–13.9° for P1 and 27.9–30.6° for P2). Ball grip angle showed a large range in the group results (outswing: −36.0–0.0°, inswing: 0.0–21.3°, ), demonstrating straight and angled seam grips. Based on the perceptions of elite bowlers and coaches in published research (Lindsay et al., Citation2024) and a Cricket Australia coaching manual (Pyke, Citation2010), individualised grips allow athletes to create the desired seam orientation based on their own movement solutions. Combining individualised grips and bowling actions also helps bowlers to maintain their finger position behind the seam of the ball at release to impart backspin and reduce unwanted seam movement (scrambled or wobbling seam) (Lindsay & Spratford, Citation2020; Lindsay et al., Citation2022, Citation2024). Bowlers combined individualised grips and techniques to create the desired seam orientation at release.
Most participants trials were predominantly assigned to one cluster (). This demonstrates differences between bowlers and is exemplified in , with most individual participants’ trials being closely grouped. The matrices also show most participants had trials assigned to multiple clusters, demonstrating intra-bowler variation. This study supports earlier research reporting both inter- and intra-athlete variability in spinal kinematics of fast bowlers (Perrett et al., Citation2020). The authors reported greater inter- than intra-participant variation, supporting the results of the current study. In addition to individualised techniques, the participants of this study employed variation in repeated deliveries.
The individualised techniques can be explained using an ecological dynamics approach where emphasis is placed on the interaction between individuals and their environment (Araújo et al., Citation2006). These complex interactions lead to the development of individualised motor solutions and movement patterns (Salter et al., Citation2007; Vantorre et al., Citation2014). As such, fast bowlers develop individualised techniques that allow them to generate swing. A fast bowler’s technique can be viewed as a dynamical system whereby movement is influenced by factors such as fatigue, ball condition, opponents, or the outcome of previous deliveries (Crowther et al., Citation2023; Phillips et al., Citation2014; Seifert et al., Citation2013). This is supported by research reporting that fast bowlers use variable movement phase timings while maintaining ball release speed (Andrews et al., Citation2024). The interactions between performers and task and environment constraints result in intra-athlete technique variability which also increases inter-athlete variation due to the continuous development and refinement of athlete-specific motor solutions and movement patterns (Vantorre et al., Citation2014). From an ecological dynamics perspective, no one “optimal” technique exists and instead, there are an infinite number of possible movement solutions that could be used by fast bowlers.
This is the first study to quantitatively investigate intra-athlete variability of upper-body kinematics in fast bowlers. Participants demonstrated differing variability in both swing directions, as shown by participant data ranges (), the proportion of individual participants’ trials assigned to multiple clusters () and variable plotting (). For example, when delivering outswing, P4 displayed lower ranges for most body kinematics and had closer trial grouping compared to P10 ( and ), indicating lower variability. Differences in playing level, with P4 being elite (senior state) and P10 being pre-elite (state u19) may have influenced this disparity. However, some elite bowlers also showed large variation, suggesting that intra-athlete variability may be individualised and not influenced by experience. In a similar overhead movement, skilled baseball pitchers have demonstrated less variation than those competing at lower levels (Fleisig et al., Citation2009). However, higher-skilled golfers have demonstrated both greater and lower levels of variability (Bradshaw et al., Citation2009; Sayers et al., Citation2017). This study appears to support the contradictions in previous literature, and research is needed to investigate movement variability in relation to swing bowling performance in athletes of different levels to provide cricket-specific evidence. Nonetheless, athletes with varying experience possess differences in skill proficiency (Phillips et al., Citation2010) and should be considered by coaches.
The bowlers in this study employed differing levels of intra-athlete variability across body segments and joints. Baseball researchers have reported stability in hand motion of successful fastball pitches despite variability in other body segments (Matsuo et al., Citation2020). Similarly, investigators have reported proximal segment variability creates end effector (distal body segment providing last point of contact with the ball) stability for consistent performance outcomes (Côté et al., Citation2002). In this study, bowlers used similar variation at the elbow, forearm, wrist and hand when examining individual athlete techniques (). Some participants used consistent global hand orientations and others employed inconsistent positions. In fastball pitches, the hand motion influences ball trajectory by controlling the projection angle and spin axis imparted on the ball (Matsuo et al., Citation2020; Whiteside et al., Citation2016). In swing bowling, bowlers aim to maintain a finger position behind the seam at release to create an angled and upright seam (Lindsay et al., Citation2024). While finger kinematics were not investigated in this study, we hypothesise that bowlers may employ low variability in the position of the index and middle fingers relative to the seam. This strategy may assist bowlers to consistently release the ball with an upright seam position.
Within proximal body segments of the pelvis, torso and thorax, lower variation was observed in most participants in comparison to bowling arm kinematics. This is consistent with researchers suggesting that certain technique parameters require greater stability than others (Gray, Citation2021; Lindsay et al., Citation2024; Whiteside et al., Citation2015). Research of tennis serving mechanics identified trunk position to be the most repeatable technique parameter with greater variation observed in arm kinematics (Whiteside et al., Citation2015). The researchers suggested this provides athletes with a consistent “posture” in repeated serves and allows the arm to manoeuvre the racquet and strike the ball. Based on the perceptions of elite cricket bowlers and coaches (Lindsay et al., Citation2024), it is possible that bowlers in this study employed similar strategies to manoeuvre their forearm and hand orientation to position their fingers on the posterior seam surface at release. We hypothesise that fast bowlers employ reduced variability in proximal body segments for consistency in their deliveries and utilise greater variation in distal body segments to manipulate the finger and ball position to achieve the desired seam orientation at release.
The movement variability observed in the current study may once have been interpreted as “noise” and “system error” (Bartlett et al., Citation2007; Hamill et al., Citation1999), however, it demonstrates the flexibility of the motor system. The traditional presentation of study results as the mean of multiple trials may prevent the identification of variability. Using individual trials and participant ranges in this study demonstrated functional movement variability that allowed bowlers to consistently generate controlled swing at match intensity. Adapting to constraints such as fatigue and the outcome of previous deliveries likely contributed to the refinement of motor solutions that influenced how movement occurred in subsequent trials (Phillips et al., Citation2012; Seifert et al., Citation2013). We hypothesise that greater intra-athlete variability would be observed in open environments, such as matches, due to the complex interaction of task, environment, and performer constraints (Crowther et al., Citation2023; Renshaw et al., Citation2022). Bowlers likely have stable and variant technique parameters that allow them to produce consistent outcomes while simultaneously adapting to constraints of performance environments (Gray, Citation2021; Lindsay et al., Citation2024; Whiteside et al., Citation2015). From a swing bowling perspective, excessive variation in large proximal body segments, such as the torso, may create variability that is dysfunctional, negatively impacting distal segments that influence the ball trajectory, seam orientation and spin axis (Lindsay & Spratford, Citation2020; Lindsay et al., Citation2022; Whiteside et al., Citation2015). Coaches and athletes should avoid striving for techniques that minimise variation (Glazier & Wheat, Citation2014) and view movement variability as functional and an essential component of adaptability (Seifert et al., Citation2013).
Some limitations exist for this study. Indoor facilities were used without a battery present, limiting task and environmental constraints, and reducing the replication of their complex interactions under match conditions. Examining technique variability in a match environment may yield different results to those presented in this study due to a greater number of interacting constraints (Crowther et al., Citation2023; Renshaw et al., Citation2022). This study captured deliveries with varying trajectories (pitching within three to seven metres from the stumps at the batter’s end and travelling within the wide lines), introducing variability into the analyses. Baseball researchers have reported that greater consistency in arm kinematics is observed in consistent pitch locations within the strike zone (Glanzer et al., Citation2021). Therefore, analysing deliveries with similar release points and trajectories may yield less variability than reported in this study. This study used tape patches attached to the ball to track ball flight and calculate swing. While steps were taken to minimise aerodynamic interference such as ball orientation and thin tape material used, this may have influenced the air surrounding the ball and subsequent flight path. Future research should explore ball-tracking technology that does not require tracking material being attached to balls. This study investigated pelvis and upper-body kinematics at the discrete point of ball release. Researchers have identified lower-body movement variability in fast bowlers (Salter et al., Citation2007) and components of the delivery stride that influence ball velocity and trajectory (Portus et al., Citation2000). Further research is required to investigate whole-body movement patterns during the entirety of the bowling action to provide a deeper understanding of the techniques employed by bowlers. Finally, only successful trials were analysed, and the differences in athlete techniques in comparison to unsuccessful deliveries is unknown. It is likely that dysfunctional movement variability was present in these trials and resulted in sub-optimal performance levels (Carson et al., Citation2014). Research is needed to compare technique variability in successful and unsuccessful swing bowling deliveries.
This study provides practical applications for coaches, athletes and researchers. Fast bowlers use individualised techniques and generalising group results to individuals should be avoided (Fisher et al., Citation2018; Phillips et al., Citation2014). When implementing strategies to improve performance, coaches should employ individualised approaches based on their observations of each technique. Additionally, constraints-led approaches to training can be used to modify the individual, task and environment constraints (Renshaw et al., Citation2022). This would challenge bowlers to seek adaptive movement solutions and explore different ways to generate swing (Phillips et al., Citation2012). From a research perspective, future investigations of fast bowling should include individual analyses to best inform practice.
Conclusion
The results of this study support the hypotheses with bowlers demonstrating inter- and intra-individual variability in bowling action kinematics. Researchers and coaches should consider how data are presented and avoid generalising group results to individuals. Individual analyses and coaching strategies should be implemented to understand and improve athletic performance. The identification of intra-individual movement variability demonstrates that coaches and athletes must understand movement variation as an essential component of adapting to individual, task and environmental constraints that allow athletes to produce consistent performance outcomes. Bowlers likely limit variability in the proximal body segments of the pelvis and torso and use greater variability in distal bowling arm segments to consistently generate swing. Stable technique parameters provide a level of repeatability to the bowling action, and variant aspects are used to manipulate the finger and ball orientation to consistently produce the desired release position. These findings could assist coaches to improve swing bowling performance in athletes of all levels.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Abernethy, B. (1981). Mechanisms of skill in cricket batting. Australian Journal of Sports Medicine, 13(1), 3–10.
- Andrews, M. H., Gorman, A. D., & Crowther, R. H. (2024). Functional movement variability to maintain delivery speed in cricket fast bowling. European Journal of Sport Science, 24(4), 415–421. https://doi.org/10.1002/ejsc.12045
- Araújo, D., Davids, K., & Hristovski, R. (2006). The ecological dynamics of decision making in sport. Psychology of Sport and Exercise, 7(6), 653–676. https://doi.org/10.1016/j.psychsport.2006.07.002
- Bartlett, R., Wheat, J., & Robins, M. (2007). Is movement variability important for sports biomechanists? Sports Biomechanics, 6(2), 224–243. https://doi.org/10.1080/14763140701322994
- Beach, A. J., Ferdinands, R. E., & Sinclair, P. J. (2016). The kinematic differences between off-spin and leg-spin bowling in cricket. Sports Biomechanics, 15(3), 295–313. https://doi.org/10.1080/14763141.2016.1161819
- Besier, T. F., Sturnieks, D. L., Alderson, J. A., & Lloyd, D. G. (2003). Repeatability of gait data using a functional hip joint centre and a mean helical knee axis. Journal of Biomechanics, 36(8), 1159–1168. https://doi.org/10.1016/S0021-9290(03)00087-3
- Bower, R. (2012). The sweet spot of a cricket bat for low speed impacts. Sports Engineering, 15(2), 53–60. https://doi.org/10.1007/s12283-012-0087-6
- Bradshaw, E. J., Keogh, J. W., Hume, P. A., Maulder, P. S., Nortje, J., & Marnewick, M. (2009). The effect of biological movement variability on the performance of the golf swing in high-and low-handicapped players. Research Quarterly for Exercise and Sport, 80(2), 185–196. https://doi.org/10.1080/02701367.2009.10599552
- Campbell, A. C., Alderson, J. A., Lloyd, D. G., & Elliott, B. C. (2009). Effects of different technical coordinate system definitions on the three dimensional representation of the glenohumeral joint centre. Medical & Biological Engineering & Computing, 47(5), 543–550. https://doi.org/10.1007/s11517-009-0467-7
- Campbell, A. C., Lloyd, D. G., Alderson, J., & Elliott, B. (2009). MRI development and validation of two new predictive methods of glenohumeral joint centre location identification and comparison with established techniques. Journal of Biomechanics, 42(10), 1527–1532. https://doi.org/10.1016/j.jbiomech.2009.03.039
- Carson, H. J., Collins, D., & Richards, J. (2014). Intra-individual movement variability during skill transitions: A useful marker? European Journal of Sport Science, 14(4), 327–336. https://doi.org/10.1080/17461391.2013.814714
- Chin, A., Lloyd, D., Alderson, J., Elliott, B., & Mills, P. (2010). A marker-based mean finite helical axis model to determine elbow rotation axes and kinematics in vivo. Journal of Applied Biomechanics, 26(3), 305–315. https://doi.org/10.1123/jab.26.3.305
- Connor, J. D., Renshaw, I., Farrow, D., & Wood, G. (2020). Defining cricket batting expertise from the perspective of elite coaches. PLOS, 15(6), e0234802. https://doi.org/10.1371/journal.pone.0234802
- Côté, J. N., Mathieu, P. A., Levin, M. F., & Feldman, A. G. (2002). Movement reorganization to compensate for fatigue during sawing. Experimental Brain Research, 146(3), 394–398. https://doi.org/10.1007/s00221-002-1186-6
- Crowther, R. H., Renshaw, I., Spratford, W. A., Sayers, M. G., & Gorman, A. D. (2023). Previous experiences can influence individual cricket spin bowlers’ perceptions of affordances in the performance landscape. Psychological Research, 87(6), 1–4. https://doi.org/10.1007/s00426-022-01769-6
- Ehrig, R. M., Taylor, W. R., Duda, G. N., & Heller, M. O. (2006). A survey of formal methods for determining the centre of rotation of ball joints. Journal of Biomechanics, 39(15), 2798–2809. https://doi.org/10.1016/j.jbiomech.2005.10.002
- Ehrig, R. M., Taylor, W. R., Duda, G. N., & Heller, M. O. (2007). A survey of formal methods for determining functional joint axes. Journal of Biomechanics, 40(10), 2150–2157. https://doi.org/10.1016/j.jbiomech.2006.10.026
- Felton, P., Lister, S., Worthington, P. J., & King, M. (2019). Comparison of biomechanical characteristics between male and female elite fast bowlers. Journal of Sports Sciences, 37(6), 665–670. https://doi.org/10.1080/02640414.2018.1522700
- Fisher, A. J., Medaglia, J. D., Jeronimus, B. F. (2018). Lack of group-to-individual generalizability is a threat to human subjects research. Proceedings of the National Academy of Sciences, 115(27), E6106–E15. https://doi.org/10.1073/pnas.1711978115
- Fleisig, G., Chu, Y., Weber, A., & Andrews, J. (2009). Variability in baseball pitching biomechanics among various levels of competition. Sports Biomechanics, 8(1), 10–21. https://doi.org/10.1080/14763140802629958
- Glanzer, J. A., Diffendaffer, A. Z., Slowik, J. S., Drogosz, M., Lo, N. J., & Fleisig, G. S. (2021). The relationship between variability in baseball pitching kinematics and consistency in pitch location. Sports Biomechanics, 20(7), 879–886. https://doi.org/10.1080/14763141.2019.1642378
- Glazier, P. S., & Mehdizadeh, S. (2019). Challenging conventional paradigms in applied sports biomechanics research. Sports Medicine, 49(2), 171–176. https://doi.org/10.1007/s40279-018-1030-1
- Glazier, P. S., & Wheat, J. S. (2014). An integrated approach to the biomechanics and motor control of cricket fast bowling techniques. Sports Medicine, 44(1), 25–36. https://doi.org/10.1007/s40279-013-0098-x
- Gray, R. (2021). How we learn to move: A revolution in the way we coach & practice sports skills.
- Hamill, J., van Emmerik RE, Heiderscheit, B. C., & Li, L. (1999). A dynamical systems approach to lower extremity running injuries. Clinical Biomechanics, 14(5), 297–308. https://doi.org/10.1016/S0268-0033(98)90092-4
- Hartigan, J. A., Wong, M. A. (1979). Algorithm as 136: A k-means clustering algorithm. Journal of the Royal Statistical Society: Series C (Applied Statistics), 28(1), 100–108. https://doi.org/10.2307/2346830
- Justham, L., Cork, A., West, A. (2010). Comparative study of the performances during match play of an elite-level spin bowler and an elite-level pace bowler in cricket. Proceedings of the Institution of Mechanical Engineers, Part P: Journal of Sports Engineering & Technology, 224(4), 237–247. https://doi.org/10.1243/17543371jset77
- Kelso, J. S. (1995). Dynamic patterns: The self-organization of brain and behavior. MIT press.
- Lindsay, C., Clark, B., Middleton, K., Crowther, R., & Spratford, W. (2022). How do athletes cause ball flight path deviation in high-performance interceptive ball sports? A systematic review. International Journal of Sports Science & Coaching, 17(3), 683–698. https://doi.org/10.1177/17479541211047360
- Lindsay, C., Crowther, R., Clark, B., Middleton, K., Keegan, R., & Spratford, W. (2024). Bowler and coach experiential knowledge of new ball swing bowling in elite cricket. Journal of sports sciences. In Press. https://doi.org/10.1080/02640414.2024.2323319
- Lindsay, C., & Spratford, W. (2020). Bowling action and ball flight kinematics of conventional swing bowling in pathway and high-performance bowlers. Journal of Sports Sciences, 38(14), 1650–1659. https://doi.org/10.1080/02640414.2020.1754717
- Lloyd, D. G., Alderson, J., & Elliott, B. (2000). An upper limb kinematic model for the examination of cricket bowling: A case study of Mutiah Muralitharan. Journal of Sports Sciences, 18(12), 975–982. https://doi.org/10.1080/026404100446775
- Matsuo, T., Jinji, T., Hirayama, D., Nasu, D., Katsumata, Y., & Morishita, Y. (2020). Consistent hand dynamics are achieved by controlling variabilities among joint movements during fastball pitching. Frontiers in Sports and Active Living, 2, 579377. https://doi.org/10.3389/fspor.2020.579377
- Mehta, R. D. (2005). An overview of cricket ball swing. Sports Engineering, 8(4), 181–192. https://doi.org/10.1007/bf02844161
- Mehta, S., Phatak, A., Memmert, D., Kerruish, S., & Jamil, M. (2022). Seam or swing? Identifying the most effective type of bowling variation for fast bowlers in men’s international 50-over cricket. Journal of Sports Sciences, 40(14), 1587–1591. https://doi.org/10.1080/02640414.2022.2094140
- Newell, K. M., & Corcos, D. M. (1993). Variability and motor control: Human kinetics publishers.
- Peploe, C., Sa, M.-N., Harland, A., & King, M. (2018). The relationships between impact location and post-impact ball speed, bat torsion, and ball direction in cricket batting. Journal of Sports Sciences, 36(12), 1407–1414. https://doi.org/10.1080/02640414.2017.1389484
- Perrett, C., Bussey, M. D., & Lamb, P. F. (2020). Movement variability in the spinal kinematics of fast bowlers. ISBS Proceedings Archive, 38(1), 284.
- Phillips, E., Davids, K., Renshaw, I., & Portus, M. (2010). Expert performance in sport and the dynamics of talent development. Sports Medicine, 40(4), 271–283. https://doi.org/10.2165/11319430-000000000-00000
- Phillips, E., Davids, K., Renshaw, I., & Portus, M. (2014). Acquisition of expertise in cricket fast bowling: Perceptions of expert players and coaches. Journal of Science and Medicine in Sport, 17(1), 85–90. https://doi.org/10.1016/j.jsams.2013.03.005
- Phillips, E., Portus, M., Davids, K., & Renshaw, I. (2012). Performance accuracy and functional variability in elite and developing fast bowlers. Journal of Science and Medicine in Sport, 15(2), 182–188. https://doi.org/10.1016/j.jsams.2011.07.006
- Portus, M., Sinclair, P. J., Burke, S. T., Moore, D. J., & Farhart, P. J. (2000). Cricket fast bowling performance and technique and the influence of selected physical factors during an 8-over spell. Journal of Sports Sciences, 18(12), 999–1011. https://doi.org/10.1080/026404100446801
- Pyke, F. (2010). Cutting edge cricket: Human kinetics.
- Renshaw, I., Davids, K., O’Sullivan, M., Maloney, M. A., Crowther, R., & McCosker, C. (2022). An ecological dynamics approach to motor learning in practice: Reframing the learning and performing relationship in high performance sport. Asian Journal of Sport and Exercise Psychology, 2(1), 18–26. https://doi.org/10.1016/j.ajsep.2022.04.003
- Robins, R., Wheat, J., Irwin, G., & Bartlett, R. (2006). The effect of shooting distance on movement variability in basketball. Journal of Human Movement Studies, 50(4), 217–238.
- Sakurai, S., Reid, M., & Elliott, B. (2013). Ball spin in the tennis serve: Spin rate and axis of rotation. Sports Biomechanics, 12(1), 23–29. https://doi.org/10.1080/14763141.2012.671355
- Salter, C. W., Sinclair, P. J., & Portus, M. R. (2007). The associations between fast bowling technique and ball release speed: A pilot study of the within-bowler and between-bowler approaches. Journal of Sports Sciences, 25(11), 1279–1285. https://doi.org/10.1080/02640410601096822
- Sarpeshkar, V., Mann, D. L., Spratford, W., & Abernethy, B. (2017). The influence of ball-swing on the timing and coordination of a natural interceptive task. Human Movement Science, 54, 82–100. https://doi.org/10.1016/j.humov.2017.04.003
- Sayers, M. G., Stuelcken, M. C., & Gorman, A. D. (2017). Functional variability in golf putting differentiates between highly skilled players. International Journal of Sport Psychology, 48(1), 70–81. https://doi.org/10.5555/20173134205
- Seifert, L., Button, C., & Davids, K. (2013). Key properties of expert movement systems in sport: An ecological dynamics perspective. Sports Medicine, 43(3), 167–178. https://doi.org/10.1007/s40279-012-0011-z
- Spratford, W., Whiteside, D., Elliott, B., Portus, M., Brown, N., & Alderson, J. (2018). Does performance level affect initial ball flight kinematics in finger and wrist-spin cricket bowlers? Journal of Sports Sciences, 36(6), 651–659. https://doi.org/10.1080/02640414.2017.1329547
- Tibshirani, R., Walther, G., & Hastie, T. (2001). Estimating the number of clusters in a data set via the gap statistic. Journal of the Royal Statistical Society: Series B (Statistical Methodology), 63(2), 411–423. https://doi.org/10.1111/1467-9868.00293
- Vantorre, J., Chollet, D., & Seifert, L. (2014). Biomechanical analysis of the swim-start: A review. Journal of Sports Science & Medicine, 13(2), 223.
- Wells, D. J., Donnelly, C. J., Elliott, B. C., Middleton, K. J., & Alderson, J. A. (2018). The inter-tester repeatability of a model for analysing elbow flexion-extension during overhead sporting movements. Medical & Biological Engineering & Computing, 56(10), 1853–1860. https://doi.org/10.1007/s11517-018-1820-5
- Whiteside, D., Chin, A., Middleton, K. (2013). The validation of a three-dimensional ball rotation model. Proceedings of the Institution of Mechanical Engineers, Part P: Journal of Sports Engineering & Technology, 227(1), 49–56. https://doi.org/10.1177/1754337112436913
- Whiteside, D., Elliott, B. C., Lay, B., & Reid, M. (2015). Coordination and variability in the elite female tennis serve. Journal of Sports Sciences, 33(7), 675–686. https://doi.org/10.1080/02640414.2014.962569
- Whiteside, D., McGinnis, R., Deneweth, J., Zernicke, R., & Goulet, G. (2016). Ball flight kinematics, release variability and in‐season performance in elite baseball pitching. Scandinavian Journal of Medicine & Science in Sports, 26(3), 256–265. https://doi.org/10.1111/sms.12443
- Winter, D. A. (2009). Biomechanics and motor control of human movement. John Wiley & Sons.
- Woolmer, B., Noakes, T., & Moffett, H. (2008). Bob Woolmer’s art and science of cricket. Struik Pub.
- Worthington, P. J., King, M. A., & Ranson, C. A. (2013). Relationships between fast bowling technique and ball release speed in cricket. Journal of Applied Biomechanics, 29(1), 78–84. https://doi.org/10.1123/jab.29.1.78
- Wu, G., Siegler, S., Allard, P., Kirtley, C., Leardini, A., Rosenbaum, D., Whittle, M., D’Lima, D. D., Cristofolini, L., Witte, H., Schmid, O., & Stokes, I. (2002). ISB recommendation on definitions of joint coordinate system of various joints for the reporting of human joint motion—part I: Ankle, hip, and spine. Journal of Biomechanics, 35(4), 543–548. https://doi.org/10.1016/S0021-9290(01)00222-6
- Wu, G., Van der Helm, F. C., Veeger, H. D., Makhsous, M., Van Roy, P., Anglin, C., Nagels, J., Karduna, A. R., McQuade, K., Wang, X., Werner, F. W., & Buchholz, B. (2005). ISB recommendation on definitions of joint coordinate systems of various joints for the reporting of human joint motion—Part II: Shoulder, elbow, wrist and hand. Journal of Biomechanics, 38(5), 981–992. https://doi.org/10.1016/j.jbiomech.2004.05.042
Appendix
Table A1. Individual participant ranges for ball grip angle, body kinematics and delivery swing angles for outswing trials.
Table A2. Individual participant ranges for ball grip angle, body kinematics and delivery swing angles for inswing trials.